Monge M, Volet DP Semir J
Texto
Imagem


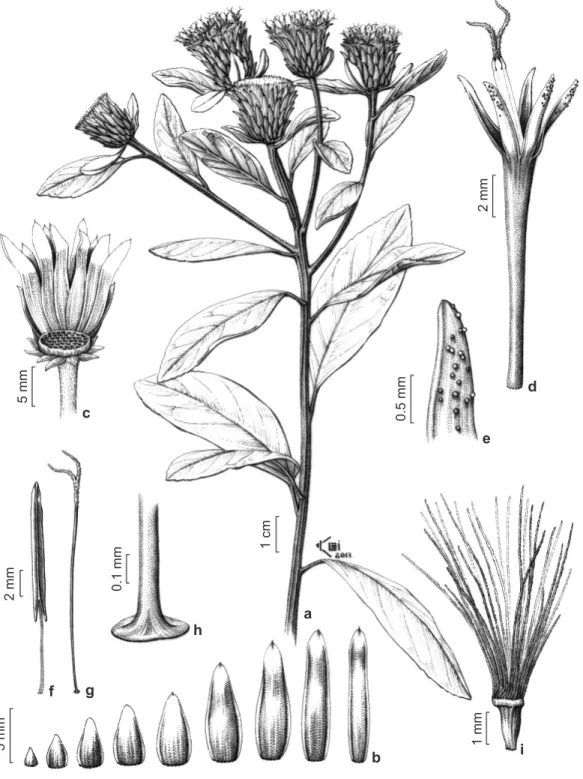
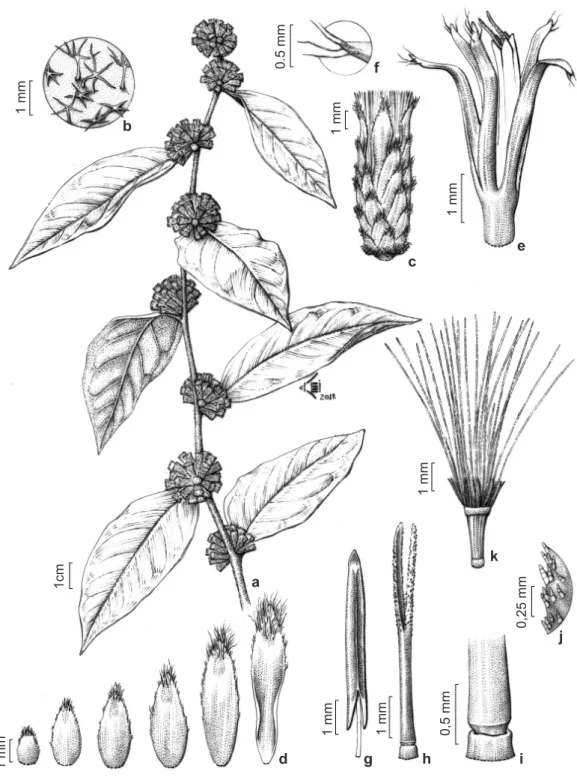

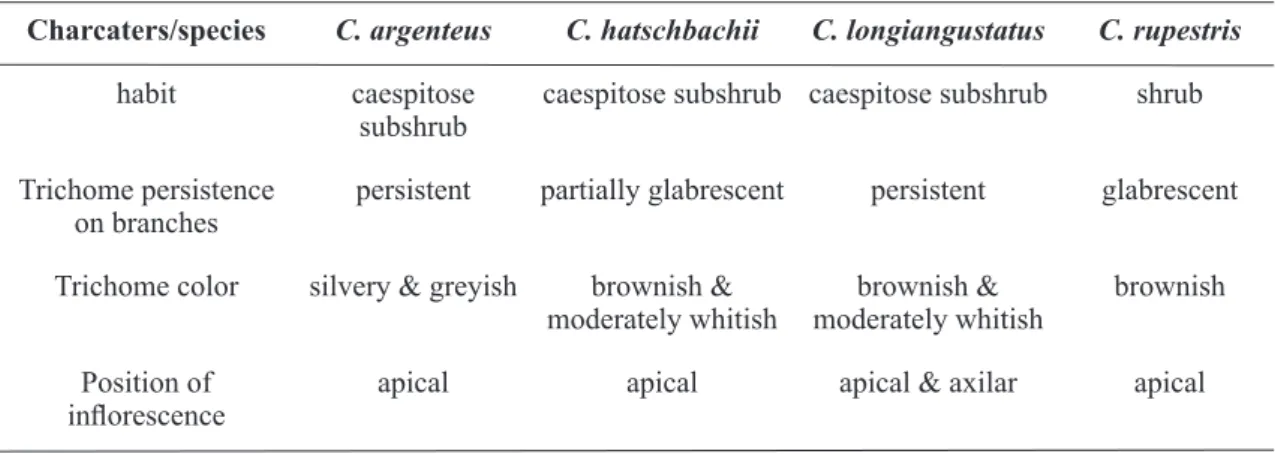
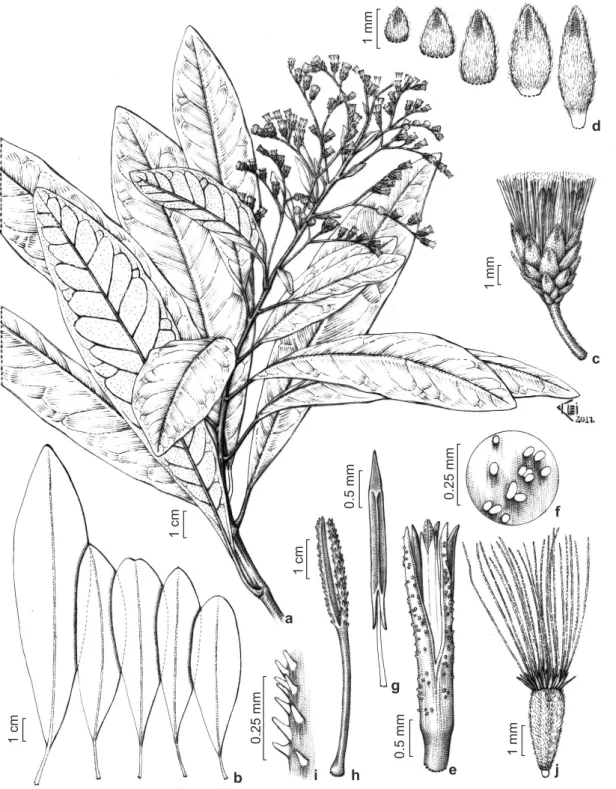
Documentos relacionados
Os resultados indicam: (a) baixa utilização de referências que abordem as teorias da contabilidade gerencial, baixa incidência de referências a artigos publicados em
O potencial de economias de energia proveniente das medidas de eficiência energética estabelecidas nos acordos de racionalização dos consumos de energia dos PREn foi de 295 ktep,
Fundamentado nos preceitos da teoria da visão baseada em recursos, o estudo investiga a cultura organizacional como recurso estratégico sustentável para o desempenho das
A infecção leva a formas sintomáticas em até 70% dos infectados, sendo suas formas clínicas, nos pacientes sintomáticos, divididas em três fases: aguda ou febril, com duração de
Entre os assuntos que podiam provocar avaliações consideradas “erradas” pelos integralistas estavam: a relação entre o integralismo e o fascismo; a questão do
Se a readmissão ocorreu no segundo ano após o desligamento, destaca- ram-se os jovens com menos de 17 anos, com a 8ª série concluída ou ainda em andamento, remuneração média de um
As identidades são compreendidas, em geral, como características específicas de um grupo que os diferenciam de outros. Em relação aos espaços de confluência de
No setor elétrico, destaca-se também no modelo baseado na taxa de chefia dos domicílios, empregado pela ANEEL para projetar o número de domicílios ocupados, urbanos e rurais, para
