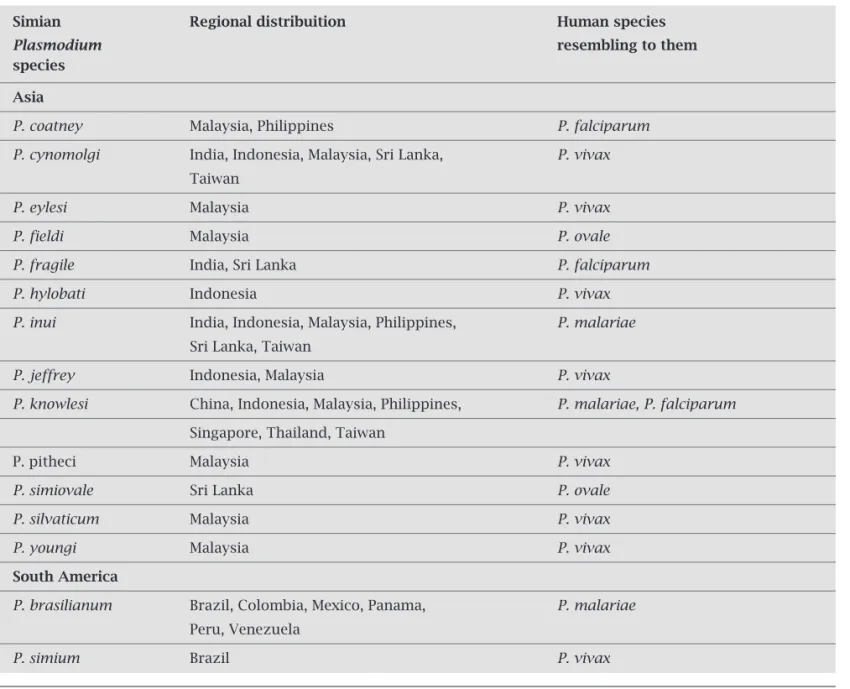
The emerging of the fifth malaria parasite
Texto
Imagem



Documentos relacionados
The specifi c interaction of the AE with ATP-dependent K + channels suggests a possible pre- synaptic modulation of the extract, since it was recently demonstrated that this
Similarly, the Australian and New Zealand College of Anaesthetists also produced a statement on fatigue where specifi c principles and responsibilities are defi ned for
The main interest of our study is that both patients were subjected to general anesthesia, with the use of a specifi c reversal agent for non-depolarizing neuromuscular block,
*** Overall and sex-specifi c MR for Brazil and regions, as well as annual Guarani mortality rates, specifi c for ICD-10 chapters, were standardized by age and sex by the
Comparison between smokers captured to a specifi c intervention for smoke cessation and the general population may help to understand the characteristics of smokers seeking
The similarities among conceptions by youth from the three countries were: the risk factors (unprotected sexual practice and contact with specifi c population groups),
This article contains a brief summary of health care models, emphasizing the role of the population survey as a management tool and analyzes the specifi c case of the
With regards to the specifi c assessment by type of cancer and race, increasing trends of mortality were acknowledged for blacks in the categories of oropha- rynx, tongue and
