Lílian Figueiredo Moreira
Tropismo tecidual dos genótipos principais de
Trypanosoma cruzi
em
camundongos BALB/c com infecções mistas, não tratados e tratados na
fase aguda da infecção, avaliados pela técnica de LSSP-PCR.
UNIVERSIDADE FEDERAL DE OURO PRETO
NÚCLEO DE PESQUISAS EM CIÊNCIAS BIOLÓGICAS – NUPEB PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
Tropismo tecidual dos genótipos principais de
Trypanosoma cruzi
em
camundongos BALB/c com infecções mistas, não tratados e tratados na
fase aguda da infecção, avaliados pela técnica de LSSP-PCR.
Autora: Lílian Figueiredo Moreira Orientadora: Profa. Dra. Marta de Lana
Co-Orientadora: Profa Dra. Helen Rodrigues Martins
Dissertação apresentada ao Programa
de Pós-Graduação em Ciências
Biológicas do Núcleo de Pesquisas em Ciências Biológicas da Universidade Federal de Ouro Preto, como requisito parcial para a obtenção do título de
Mestre; área de concentração:
Imunobiologia de Protozoários.
M838t Moreira, Lilian Figueiredo
Tropismo tecidual dos genótipos principais de Trypanosoma cruzi em camundongos BALB/c com infecções mistas, não tratados e tratados na fase aguda da infecção, avaliados pela técnica de LSSP-PCR [manuscrito] / Lilian Figueiredo Moreira - 2009.
150 f.: grafs.; tabs.
Orientadora: Profª. Drª. Marta de Lana.
Co-orientadora: Profª. Drª. Helen Rodrigues Martins
Dissertação (Mestrado) - Universidade Federal de Ouro Preto. Instituto de Ciências Exatas e Biológicas. Núcleo de Pesquisas em Ciências Biológicas. Programa de Pós-graduação em Ciências Biológicas.
Área de concentração: Imunobiologia de Protozoários.
1. Trypanosoma cruzi - Teses. 2. Genótipos - Teses. 3. Infecção - Teses. 4. Benzonidazol - Teses. 5. Tecidos (Anatomia e fisiologia) - Cultura e meios de cultura - Teses. I. Universidade Federal de Ouro Preto. II. Título.
CDU: 616.937:663.1
iii
"Aquele que não tem um objetivo, raramente sente prazer
em qualquer empreendimento."
Figueiredo, L.M. Dedicatória
iv
Dedicatória
Aos meus pais Célia e Antônio, à minha irmã Sônia, à minha avó Nhanhá e tias Verinha, Ritinha e Nenem por toda dedicação, amor, confiança e por sempre apoiarem as minhas decisões...
Figueiredo, L.M. Agradecimentos
v
Agradecimentos
Àqueles que diretamente contribuíram para a realização desse trabalho...
A Deus, por sempre fazer o melhor em minha vida....por todas as minhas conquistas e por colocar no meu caminho pessoas tão especiais...
À professora Marta de Lana, por me acolher tão bem no laboratório de doença de Chagas e por ter acreditado em mim para a realização desse trabalho. Agradeço pelo seu otimismo em sua orientação e pelos ensinamentos transmitidos. Só tenho a agradecer as verdadeiras demonstrações de carinho...
À Helen Rodrigues Martins, agradeço pela grande co-orientação desse trabalho. Devo a você grande parte do meu aprendizado sobre a doença de Chagas. Você me ensinou a dar os primeiros passos e hoje posso compartilhar aqui muito daquilo que aprendi a seu lado....Muito obrigada!
Aos professores do Laboratório de doença de Chagas Maria Terezinha Bahia, André Talvani e Evandro Marques de Menezes Machado pela amizade e por ter esclarecido tantas dúvidas...
À Cláudia Martins Carneiro, pelo auxílio na realização das necropsias, por toda atenção dada sempre que precisei e por ver as coisas sempre de uma maneira tão simples...
À Andréa Mara Macedo, por ter aberto as portas do seu laboratório para que eu pudesse aprender as metodologias necessárias na execução desse trabalho.
Ao Claudiney, por ter me ensinado, com toda a paciência, a técnica de LSSP-PCR utilizada nesse estudo. Obrigada pela atenção!
Figueiredo, L.M. Agradecimentos
vi
equipamentos e reagentes tão necessários na execução desse projeto e pela disponibilidade em me atender.
À professora Simone Resende por sua gentileza em fornecer alguns animais utilizados nesse projeto.
À Vanja, pela amizade e presteza durante todos esses anos. Os conhecimentos adquiridos com você foram primordiais no meu crescimento e execução desse trabalho. Você é uma pessoa maravilhosa!
À Jack, uma irmã que Deus me deu para caminhar junto a mim nessa etapa tão importante de nossas vidas...te agradeço por toda ajuda cedida todas as vezes que eu precisei... com você aprendi e dividi tantos momentos... foram tantas risadas, choros, apertos e cansaço....agradeço pela sincera amizade compartilhada durante todos esses anos de convivência...você é uma pessoa maravilhosa e fez toda a diferença....obrigada por tudo!
Ao Maykon, pela grande amizade e por toda ajuda cedida na realização desse trabalho. Sem você a concretização desse sonho seria bem mais difícil...obrigada por tornar o ambiente de trabalho mais agradável....meus sinceros agradecimentos....
À Míriam, pessoa maravilhosa e sempre prestativa. Obrigada por ter me esclarecido tantas dúvidas durante o mestrado. Agradeço também pela verdadeira amizade oferecida e por todos os conselhos dados quando eu mais precisei...
Aos amigos do Laboratório de doença de Chagas Maykon, Lívia, Auffy, Dani, Vitor, Lucas, Renata Branquinho, Gláucia, Alexandre Rotondo, Lílian Melo, Guilherme, André Gravel, Régia, Rafael, Tassiane, Ivo, Vivian, Fabiane, Maíra, Magna e Isabel pela maravilhosa convivência, amizade e ajuda durante todos esses anos...
Figueiredo, L.M. Agradecimentos
vii
Stêfany e em especial ao Ricardo, Lorena e Fábio que me ensinaram muito durante a minha iniciação científica.
Aos amigos do LIP, Míriam, Rodrigo, Eduardo, Tiago, Roberta, Pedro, Amanda, Pauline, Leandro e Marco pelos momentos de descontração e disponibilidade em ajudar...muito obrigada!
Aos amigos do Laboratório de Bioquímica e Patologia pelo material cedido e ajuda sempre que precisei.
Aos meus amigos da turma de mestrado: Jack, Lílian, Larissa, Melina, Lorena, Tiago, João, Emanuel e Míriam pela agradável convivência durante as disciplinas....
À Ana Salomé, pessoa maravilhosa e sempre de bom humor, agradeço pelo cuidado com o material utilizado na realização desse trabalho.
Aos bioteristas, em especial à Cristina pelo cuidado com os animais.
À Érika pela agradável convivência e pela atenção dada em todos os momentos.
À Cida pela disponibilidade em resolver todos os problemas....
Ao biotério da UFMG por ter cedido os animais utilizados nesse trabalho.
À UFOP, pelo ensino de qualidade e possibilidade em realizar esse trabalho.
À CAPES, pela concessão da bolsa de mestrado.
Figueiredo, L.M. Agradecimentos
viii
Agradeço também à algumas pessoas que indiretamente contribuíram para a realização desse sonho...
Às minhas companheiras de república: Jack, Marisa, Mari, Débis e Taís por todos esses anos de convivência, companheirismo, cumplicidade e amizade sincera... vocês foram pessoas fundamentais durante todos esses anos, tornando os meus dias sempre mais agradáveis e me fazendo muito mais feliz! Meus sinceros e eternos agradecimentos...
Ao Sr. Deusdedit e família e à Lúcia por todo carinho, atenção e ajuda prestada durante todos esses anos de moradia em Ouro Preto. Muito obrigada!
Aos meus adoráveis e eternos amigos Geo, Arnaldo, Maykon, Marcela, Claudinha, Lívia, Rúbia, Carol e Douglas. A amizade de vocês foi fundamental para vencer mais essa etapa....foram tantos os momentos vividos juntos....quantas risadas e choros compartilhados....
À minha querida família, em especial aos meus pais, avó, irmã e às minhas tias Verinha, Ritinha e Nenem por todo carinho e amor e por acreditarem em mim e nunca pouparem esforços em tornar mais esse sonho realidade... amo vocês!
Ao Djalma, meu amor, por sempre apoiar as minhas decisões e por me estimular a fazer sempre o melhor... você mais do que ninguém sabe o significado desta conquista, pois esteve ao meu lado de forma incondicional. Só tenho a agradecer tanto amor e carinho....
Figueiredo, L.M. Resumo
ix
A correlação entre a genética do T. cruzi e as formas clínicas da doença de Chagas não está bem estabelecida. Apesar da relação parasito x hospedeiro ser complexa e multifatorial, uma das razões para essa falta de correlação pode ser a freqüente ocorrência de infecções mistas na natureza. Nesse sentido, o principal objetivo desse estudo foi avaliar, por meio da técnica de LSSP-PCR, o impacto de infecções mistas no tropismo tecidual de clones de T. cruzi pertencentes aos genótipos principais 19, 20, 39 e 32 na fase crônica da infecção. Para tal, foram avaliados seis tecidos (coração, músculo esquelético, cólon, baço, bexiga e cérebro) de quatro camundongos BALB/c fêmeas com infecção mista por clones de T. cruzi de cada genótipo principal. Os animais foram inoculados, via IP, com 5.000 tripomastigotas sanguíneos de cada clone, combinados aos pares, totalizando 24 misturas distintas. Esses camundongos foram divididos em dois grupos: não tratados (INT) e tratados (IT) com benzonidazol (BZ) e avaliados comparativamente às respectivas infecções monoclonais (10.000 tripomastigotas sanguíneos). A técnica de PCR detectou o kDNA dos oito clones de T. cruzi em todos os seis tecidos avaliados de animais INT. Após o tratamento houve redução na positividade da PCR especialmente nos animais infectados com clones dos genótipos 32 e 39 em comparação com aqueles infectados com os genótipos 19 e 20. Não foi observada uma associação entre a distância filogenética dos clones e o tropismo tecidual do kDNA do T. cruzi. Entretanto, clones dos genótipos 19 e 20 foram os únicos que apresentaram, durante a fase crônica, positividade na PCR no cérebro e maior positividade na bexiga. A comparação entre a distribuição tecidual do kDNA de clones do T. cruzi em camundongos com infecção mista e monoclonal revelou que na maioria das misturas estudadas, exceto Cuica cl1 + SO3 cl5 (20+39) houve alteração do tropismo tecidual do kDNA nos animais INT e IT. A maioria dos tecidos de animais com infecção mista (INT e IT) apresentou PCR negativa, ao contrário do observado nos animais com infecção monoclonal. Entretanto, a PCR foi positiva em animais infectados com 18/24 misturas. A identificação dos clones presentes nesses tecidos por LSSP-PCR revelou resultados esperados, considerando as infecções monoclonais, na maioria das misturas. Curiosamente, o perfil genético dos clones remanescentes nos tecidos de animais infectados com as misturas OPS21 cl11 + P209 cl1 e OPS21 cl11 + Bug2148 cl1 e tratados demonstrou a presença do clone OPS21 cl1, 100% susceptível ao BZ. Outro resultado interessante foi obtido na caracterização dos clones remanescentes nos tecidos de animais infectados com a mistura SO3 cl5 + IVV cl4 (39+32) tratados com BZ, que revelou um perfil indeterminado não compatível com o esperado para os clones originais utilizados na inoculação. A caracterização por LSSP-PCR e microssatélites dos clones presentes nos tecidos de animais infectados com as misturas que apresentaram alteração no perfil de susceptibilidade ao BZ revelou resultados semelhantes e esperados, considerando as infecções monoclonais, na maioria das misturas. Além disso, a técnica de LSSP-PCR revelou alteração no tropismo tecidual do kDNA dos clones nesses mesmos animais, sugestiva de interação entre os parasitos componentes da mistura. A técnica de LSSP-PCR apresentou maior sensibilidade quando comparada à de microssatélites, sendo capaz de detectar uma maior porcentagem de ambos os clones nos tecidos de animais INT. A identificação por LSSP-PCR dos clones remanescentes nos tecidos de animais considerados curados (TC) e que apresentaram PCR em tecido positiva demonstrou apenas a presença de um clone e os resultados foram semelhantes àqueles encontrados nos animais tratados não curados (TNC), levantando um importante questionamento a respeito do critério de cura na doença de Chagas. Somente clones dos genótipos 19 ou 20 estavam presentes nos tecidos de animais TC e infectados com as combinações genotípicas 19+32, 19+39 e 20+39, exceto na mistura Cuica cl1 + Bug2148 cl1 (20+39), que demonstrou a presença do clone pertencente ao genótipo 39. Os resultados obtidos nesse trabalho revelaram alteração no tropismo tecidual do kDNA do T. cruzi em infecções mistas sugestiva de interações entre os clones da mistura. Os dados em conjunto sugerem que a correlação entre a genética do parasito e as diferentes formas clínicas da doença é ainda mais difícil de ser estabelecida em infecções mistas em relação às infecções monoclonais.
Figueiredo, L.M. Abstract
x
The correlation between the T. cruzi genetics and the clinical forms of Chagas’ disease is
not well established. Despite the host x parasite relationship be complex and multifactorial one reason for this may be the frequent occurrence in nature of mixed infections. The aim of this study was to evaluate by LSSP-PCR technique, the impact of mixed infections in tissue tropism of T. cruzi clonal stocks from major genotypes 19, 20, 39 and 32 in the chronic
phase of infection. For this, were evaluated six tissues (heart, skeletal muscle, colon, spleen, bladder and brain) of four female BALB/c mice with mixed infection by IP route, with
T. cruzi clonal stocks from each genotype (5.000 blood trypomastigotes of each) combined
in pairs, in a total of 24 different mixtures. Mice were divided in two groups: not treated (INT) and treated (IT) with benznidazole (BZ) and compared to the respective monoclonal infection (10.000 blood trypomastigotes of each clone). The PCR technique detected kDNA
of eight clones of T. cruzi in all six tissues of INT animals. After treatment it was observed
reduction of the PCR positivity, particularly in animals infected with clones of genotypes 32 and 39 when compared with those infected with genotypes 19 and 20. No association was observed between the parasites phylogenetic distance and the tissue tropism of T. cruzi
kDNA. However, clones of genotypes 19 and 20 were the only who showed during the chronic phase, PCR positivity in the brain and higher positivity in bladder. The comparison between the tissue distribution of T. cruzi kDNA of each clone in mice with monoclonal
and mixed infection revealed that most of the mixtures studied, except Cuica cl1 + SO3 cl5 (20 +39) changed the kDNA tissue tropism, both in IT and INT animals. Most tissues of animals with mixed infection (INT and IT) presented negative PCR, in contrast to that verified in animals with monoclonal infection. However, the PCR was positive in animals infected with 18/24 mixtures. The identification of each clone present in these tissues by LSSP-PCR revealed the expected results considering the monoclonal infections in the majority of the mixtures. Interestingly, the genetic profile of the remaining clonal stock in tissues of animals infected with treated mixtures OPS21 cl11 + P209 cl1 and OPS21 cl11 + Bug2148 cl1 demonstrated the presence of the clone OPS21 cl1, 100% susceptible to BZ. Another interesting result was obtained in the characterization of the remaining clones in tissues of animals infected with the mixture SO3cl5 + IVV cl4 (39 +32) treated with BZ, which showed an indeterminate profile not compatible with the expected for the original clones used for inoculation. The characterization by LSSP-PCR and microsatellite of the clones present in tissues of animals infected with mixtures that showed change in the BZ susceptibility profile showed expected and similar results considering the monoclonal infections, in most of the mixtures. Moreover, the technique of LSSP-PCR revealed changes in tissue tropism of kDNA of each clone in these animals, suggestive of interaction phenomena between the parasites present in the mixture. The technique of LSSP-PCR showed higher sensitivity when compared to microsatellites being able to detect a higher percentage of both clones in tissues of INT animals. The LSSP-PCR for identification of clones remaining in tissues from animals considered cured (TC), which showed positive tissue-PCR revealed only the presence of one clone and the results were similar to those obtained in treated not cured (TNC) animals, raising an important question regarding the cure criteria in Chagas’ disease. Only clones of genotypes 19 and 20 were present in tissues of TC animals infected with and the genotype combinations 19+32, 19+39 and 20+39, except the mixture Cuica cl1 + Bug2148 cl1 (20 +39), which presented the presence of the clone belonging to genotype 39. The results obtained in this study showed changes in tissue
tropism of T. cruzi kDNA in mixed infections, suggestive of interactions between the clones
of the mixture. Data together suggest that the genetic correlation between the parasite genetic and the different clinical forms of the disease is much more difficult to be established in mixed infections compared to monoclonal infections.
Figueiredo, L.M. Lista de Tabelas
xi
Lista de tabelas
Tabela I: Hospedeiros, origem geográfica, virulência e susceptibilidade ao benzonidazol dos oito clones de Trypanosoma cruzi utilizados nesse estudo. ... 28
Tabela II: Misturas de clones do Trypanosoma cruzi pertencentes aos genótipos
principais utilizados na infecção de camundongos BALB/c combinados segundo o padrão de virulência e susceptibilidade ao benzonidazol... 30
Tabela III: Misturas teóricas de clones de Trypanosoma cruzi pertencentes às
diferentes misturas utilizadas na avaliação da sensibilidade da técnica de LSSP-PCR. ... 35
Tabela IV: Misturas de clones do Trypanosoma cruzi pertencentes aos genótipos
principais utilizados na infecção de camundongos BALB/c combinados segundo o padrão de virulência e susceptibilidade ao benzonidazol não tratados e tratados com este fármaco... 36
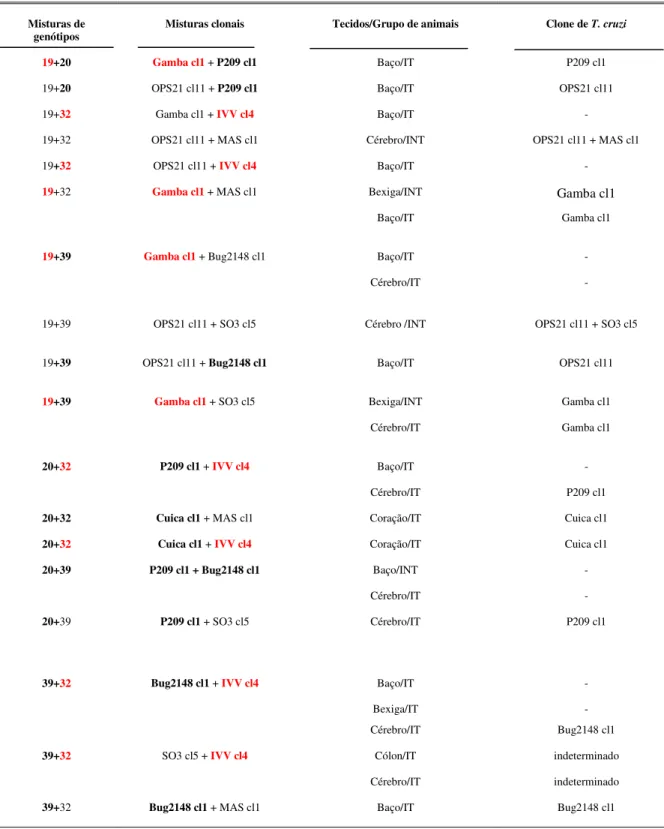
Tabela V: Alteração na distribuição tecidual de clones do Trypanosoma cruzi em
camundongos BALB/c com infecções mistas pertencentes aos genótipos principais avaliados pela técnica de PCR. ... 47
Tabela VI: Identificação de clones de Trypanosoma cruzi por LSSP-PCR provenientes
de tecidos de camundongos BALB/c infectados com misturas de clones e que apresentaram positividade na PCR quando comparado com as suas respectivas infecções monoclonais. ... 51
Tabela VII: Identificação de isolados de Trypanosoma cruzi provenientes de
camundongos BALB/c infectados com as nove misturas que apresentaram alteração no perfil de susceptibilidade ao benzonidazol e nas três misturas caracterizadas pela técnica de microssatélites em que houve predominância do clone mais susceptível ao benzonidazol*. ... 71
Tabela VIII: Distribuição tecidual do kDNA de clones de Trypanosoma cruzi
Figueiredo, L.M. Lista de Tabelas
xii
Tabela IX: Identificação de clones do Trypanosoma cruzi em tecidos por LSSP-PCR e
no sangue e tecidos por isoenzimas* e microssatélites* de camundongos BALB/c infectados com diferentes misturas pertencentes aos genótipos principais que apresentaram alteração no perfil de susceptibilidade ao benzonidazol e nas três misturas em que houve predominância do clone mais susceptível ao benzonidazol**... 81
Tabela X: Identificação de clones de Trypanosoma cruzi provenientes de camundongos
BALB/c com infecções mistas tratados com benzonidazol e considerados curados (TC) pelo critério de cura e que apresentaram PCR em tecido positiva. ... 83
Tabela X (continuação): Identificação de isolados de Trypanosoma cruzi provenientes
Figueiredo, L.M. Lista de Figuras
xiii
Lista de figuras
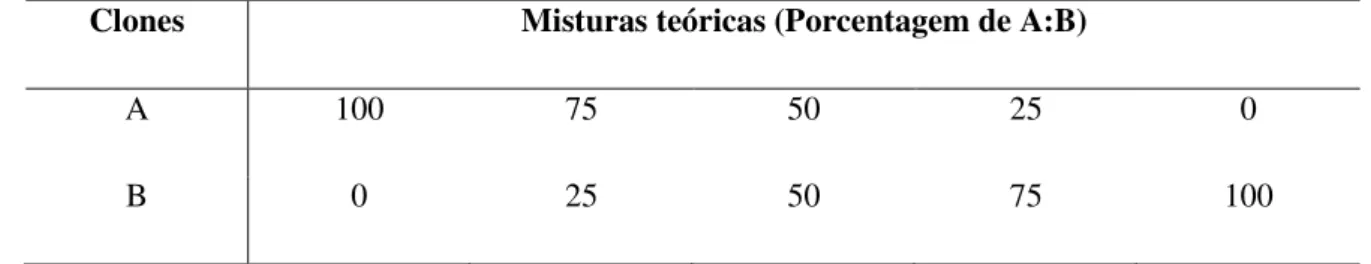
Figura 1: Positividade global da PCR para o kDNA do Trypanosoma cruzi em
diferentes tecidos de animais na fase crônica da infecção não tratados (IT, n= 16) e tratados com benzonidazol (INT, n= 16). ... 40
Figura 2: Positividade global da PCR para o kDNA do Trypanosoma cruzi em
diferentes tecidos de animais com infecções monoclonais pelos genótipos principais desse parasito não tratados (INT, n= 16) e tratados com benzonidazol (IT, n=16) na fase crônica da infecção. ... 41
Figura 3: Positividade da PCR para o kDNA do Trypanosoma cruzi em diferentes
tecidos na fase crônica da infecção de animais com infecção monoclonal por clones pertencentes aos genótipos principais, não tratados (INT, n=16) e tratados (IT, n=16) com benzonidazol. ... 42
Figura 4: Perfis de LSSP-PCR representativos dos oito clones de Trypanosoma cruzi
pertencentes aos genótipos principais (19, 20, 39 e 32) revelados em gel de poliacrilamida 6% e corados pela prata. PM: Padrão de peso molecular 100pb; clones P209 cl1 e Cuica cl1 (genótipo 20, T. cruzi I), clones Gamba cl1 e OPS21
cl11 (genótipo 19, T. cruzi I), clones Bug2148 cl1 e SO3 cl5 (genótipo 39, T. cruzi)
e clones IVV cl4 e MAS cl1 (genótipo 32, T. cruzi II). ... 45
Figura 5: Perfis de LSSP-PCR obtidos a partir de misturas teóricas de DNA do
Trypanosoma cruzi (A+B), revelados em gel de poliacrilamida 6% e corados pela
prata. Canaleta 1: PM: padrão de peso molecular 100pb; 2: DNA de referência do clone Gamba cl1 (100%); 3: mistura dos clones Gamba cl1 + IVV cl4 (75% + 25%); 4: mistura dos clones Gamba cl1 + IVV cl4 (50% + 50%); 5: mistura dos clones IVV cl4 + Gamba cl1 (75% + 25%); 6: DNA de referência do clone IVV cl4 (100%). ... 45
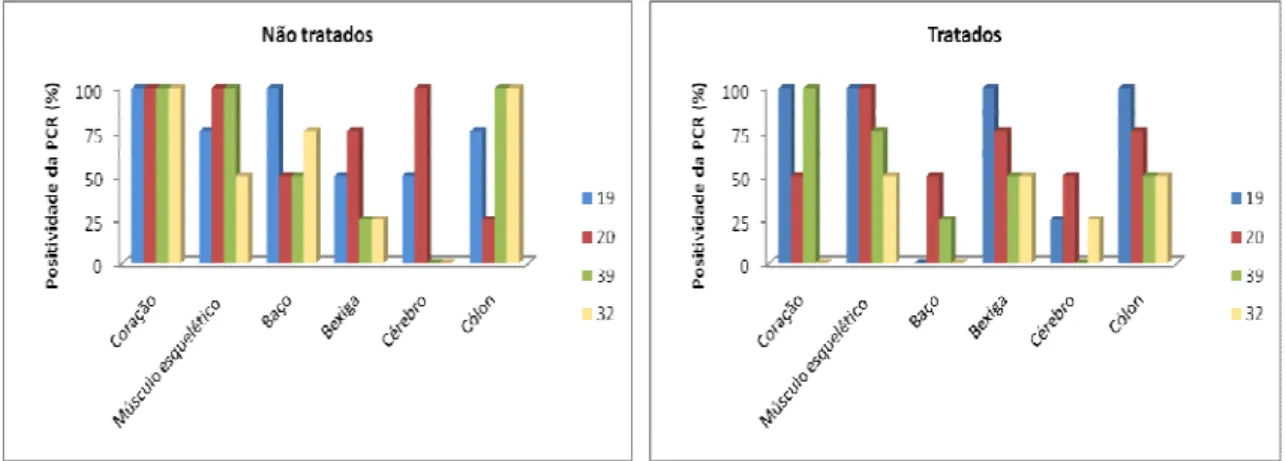
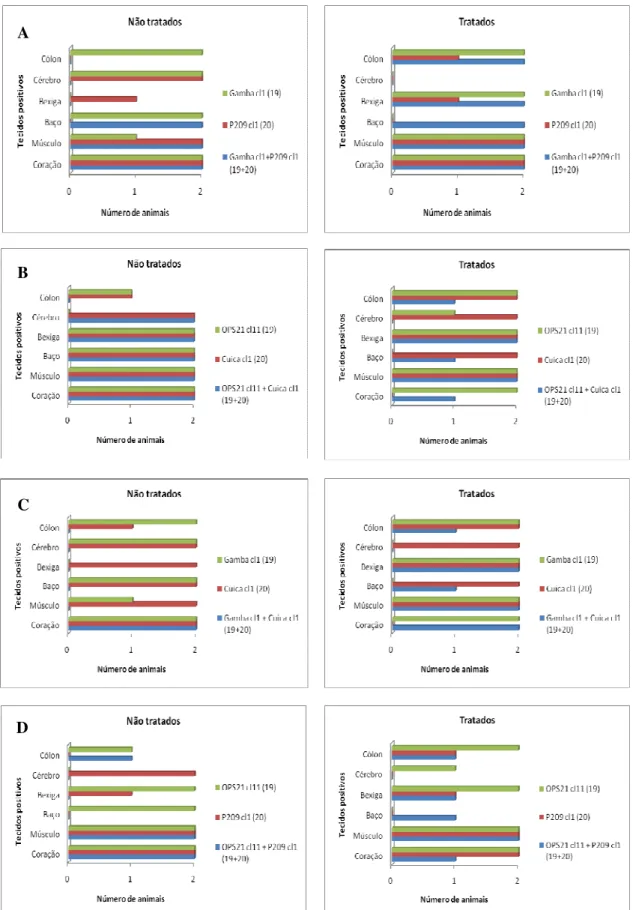
Figura 6: Comparação do tropismo tecidual do kDNA de clones de Trypanosoma cruzi
Figueiredo, L.M. Lista de Figuras
xiv
Figura 7: Perfil de LSSP-PCR representativo dos resultados obtidos na identificação de clones remanescentes nos tecidos de animais tratados e infectados com as misturas Gamba cl1 + P209 cl1: IT1 e IT2 e OPS21 cl11 + P209 cl1: IT3-IT5. IT: clones remanescentes em tecidos de animais tratados com benzonidazol. ... 52
Figura 8: Comparação do tropismo tecidual do kDNA de clones de Trypanosoma cruzi
em camundongos com infecções mistas pelos genótipos principais 19+32 não tratados (INT) e tratados (IT) com benzonidazol com suas respectivas infecções monoclonais na fase crônica da infecção. ... 53
Figura 9: Perfil de LSSP-PCR representativo dos resultados obtidos da identificação de clones presentes nos tecidos de animais infectados com a mistura OPS21 cl11 + MAS cl1 não tratados com benzonidazol. INT: clones presentes em tecidos de animais não tratados com benzonidazol. ... 54
Figura 10: Perfil de LSSP-PCR representativo dos resultados obtidos da identificação de clones presentes nos tecidos de animais infectados com a mistura Gamba cl1 + MAS cl1 não tratados e tratados com benzonidazol. INT: clones presentes em tecidos de animais não tratados com benzonidazol. IT: clones remanescentes em tecidos de animais tratados com benzonidazol. ... 55
Figura 11: Comparação do tropismo tecidual do kDNA de clones de Trypanosoma
cruzi em camundongos com infecções mistas pelos genótipos principais 19+39 não
tratados (INT) e tratados (IT) com benzonidazol com suas respectivas infecções monoclonais na fase crônica da infecção. ... 57
Figura 12: Comparação do tropismo tecidual do kDNA de clones de Trypanosoma
cruzi em camundongos com infecções mistas pelos genótipos principais 20+32 não
tratados (INT) e tratados (IT) com benzonidazol com suas respectivas infecções monoclonais na fase crônica da infecção. ... 60
Figueiredo, L.M. Lista de Figuras
xv
Figura 14: Comparação do tropismo tecidual do kDNA de clones de Trypanosoma
cruzi em camundongos com infecções mistas pelos genótipos principais 20+39 não
tratados (INT) e tratados (IT) com benzonidazol com suas respectivas infecções monoclonais na fase crônica da infecção. ... 63
Figura 15: Perfil de LSSP-PCR representativo dos resultados obtidos da identificação de clones presentes nos tecidos de animais infectados com a mistura P209 cl1 + SO3 cl5 tratados com benzonidazol. IT: clones remanescentes em tecidos de animais tratados com benzonidazol... 65
Figura 16: Comparação do tropismo tecidual do kDNA de clones de Trypanosoma
cruzi em camundongos com infecções mistas pelos genótipos principais 39+32 não
tratados (INT) e tratados (IT) com benzonidazol com suas respectivas infecções monoclonais na fase crônica da infecção. ... 66
Figura 17: Perfil de LSSP-PCR representativo dos resultados obtidos da identificação de clones remanesentes nos tecidos de animais infectados com a mistura SO3 cl5 + IVV cl4 tratados com benzonidazol. IT: clones remanescentes em tecidos de animais tratados com benzonidazol... 68
Figura 18: Perfil de LSSP-PCR representativo dos resultados obtidos de isolados de animais infectados com diferentes combinações genotípicas do Trypanosoma cruzi
e que apresentaram alteração no perfil de susceptibilidade ao benzonidazol. INT:
isolados de animais não tratados com benzonidazol. TNC: isolados de animais tratados com benzonidazol e não curados. ... 74
Figura 19: Perfil de LSSP-PCR representativo dos resultados obtidos de isolados de animais infectados com diferentes combinações genotípicas do Trypanosoma cruzi
Figueiredo, L.M. Lista de Abreviaturas
xvi
Lista de abreviaturas
AATV Pesquisa de anticorpos antitripomastigotas vivos pela citometria de fluxo
BZ Benzonidazol
CEBIO Biotério do Instituto de Ciências Biológicas
d.a.i. Dias após a infecção
dATP 2’-desoxinucleotídeo de Adenosina-5’-trifosfato
dCTP 2’-desoxinucleotídeo de Citocina-5’-trifosfato
dGTP 2’-desoxinucleotídeo de Guanina-5’-trifosfato
dTTP 2’-desoxinucleotídeo de Timina-5’-trifosfato
dNTP’s 2’-desoxinucleotídeos-5’-trifosfato
DTU Discrete typing unity
ELISA Enzyme Linked-Immuno-Sorbent Assay
ESF Exame de sangue a fresco
FA Fase aguda
FC Fase crônica
Hc Hemocultura
INT Infectado não tratado
IP Intraperitoneal
IT Infectado tratado
kDNA DNA do cinetoplasto
LSSP-PCR Low-stringency Single Primer PCR
TC Tratado curado
Figueiredo, L.M. Sumário
xvii
Sumário
1.0) Introdução ... 20
1.1) A doença de Chagas e o Trypanosoma cruzi ... 2
1.2) Variabilidade genética do T. cruzi ... 3
1.3) Correlação entre a diversidade genética do T. cruzi e suas propriedades biológicas ... 11
1.4) Genética do parasito e distribuição tecidual do T. cruzi ... 14
1.5) Genética do hospedeiro e distribuição tecidual do T. cruzi ... 15
1.6) Infecções mistas ou policlonais e suas conseqüências para o hospedeiro ... 17
2.0) Justificativa ... 21
3.0) Objetivos ... 23
3.1) Objetivo Geral ... 24
3.2) Objetivos específicos ... 24
4.0) Delineamento experimental ... 25
5.0) Animais, material e métodos ... 27
5.1) Clones do Trypanosoma cruzi ... 28
5.2) Obtenção dos inóculos ... 29
5.3) Infecções experimentais avaliadas ... 29
5.3.1) Confirmação da infecção ... 30
5.4) Tratamento dos animais ... 31
5.4.1) Fármaco ... 31
5.4.2) Esquema de tratamento ... 31
Figueiredo, L.M. Sumário
xviii
5.6) Comparação do tropismo tecidual do kDNA do T. cruzi nas infecções monoclonais e mistas tratadas e não tratadas pela técnica de PCR ... 32
5.6.1) PCR em tecido ... 32
5.7) Determinação do perfil de LSSP-PCR dos oito clones de T. cruzi pertencentes aos genótipos principais ... 33
5.7.1) Caracterização genética por LSSP-PCR ... 34
5.8) Avaliação da sensibilidade da técnica de LSSP-PCR para detecção dos clones de T. cruzi nos tecidos de animais com infecções mistas ... 35
5.9) Identificação das populações do T. cruzi presentes nos tecidos de camundongos com infecções mistas não tratados e tratados com benzonidazol pela técnica de LSSP-PCR ... 35
6.0) Resultados ... 38
6.1) Positividade da PCR em tecidos de animais com infecção monoclonal por clones de T.
cruzi não tratados (INT) e tratados (IT) com benzonidazol (BZ) agrupados de acordo com os genótipos principais ... 40
6.2) Determinação do perfil de LSSP-PCR dos oito clones de T. cruzi pertencentes aos genótipos principais ... 44
6.2.1) Avaliação da sensibilidade da técnica de LSSP-PCR para detecção dos clones de T. cruzi em infecções mistas ... 45 6.3) Comparação pela técnica de PCR do tropismo tecidual de clones de T. cruzi em camundongos com infecções monoclonais e mistas submetidos ou não ao tratamento com BZ e identificação dos clones presentes nos tecidos por LSSP-PCR ... 46
6.3.1) Avaliação do tropismo tecidual do kDNA do T. cruzi e identificação dos clones presentes em tecidos de animais infectados com a combinação genotípica 19+20... 48
6.3.2) Avaliação do tropismo tecidual do kDNA de T. cruzi e identificação dos clones presentes em tecidos de animais infectados com a combinação genotípica 19+32... 52
Figueiredo, L.M. Sumário
xix
6.3.4) Avaliação do tropismo tecidual do kDNA do T. cruzi e identificação dos clones presentes em tecidos de animais infectados com a combinação genotípica 20+32... 59
6.3.5) Avaliação do tropismo tecidual do kDNA de T. cruzi e identificação dos clones presentes em tecidos de animais infectados com a combinação genotípica 20+39... 62
6.3.6) Avaliação do tropismo tecidual do kDNA de T. cruzi e identificação dos clones presentes em tecidos de animais infectados com a combinação genotípica 39+32... 65
6.4) Identificação de clones de T. cruzi remanescentes nos tecidos de camundongos infectados com misturas pertencentes aos genótipos principais que apresentaram alteração no perfil de susceptibilidade ao benzonidazol ... 69
6.5) Identificação de clones de T. cruzi remanescentes nos tecidos de camundongos infectados com misturas dos genótipos principais e tratados com BZ considerados curados (TC) pelo critério de cura utilizado e que apresentaram PCR em tecido positiva ... 82
7.0) Discussão ... 89
7.1) Avaliação da positividade da PCR em camundongos com infecção monoclonal pelos genótipos principais de T. cruzi não tratados e tratados com o benzonidazol ... 90 7.2) Tropismo tecidual de clones de T. cruzi pertencentes aos genótipos principais em camundongos com infecção mista e monoclonal não tratados e tratados com benzonidazol e identificação nos tecidos que apresentaram alteração na positividade da PCR ... 94
7.3) Identificação de clones de T. cruzi remanescentes nos tecidos de camundongos infectados com misturas que apresentaram alteração no perfil de susceptibilidade ao benzonidazol ... 99
7.4) Identificação de clones remanescentes nos tecidos de animais tratados curados (TC) que apresentaram positividade na PCR em tecido ... 102
8.0) Conclusões ... 106
Figueiredo, L.M. Introdução
2 1.1) A doença de Chagas e o Trypanosoma cruzi
O Trypanosoma (Schizotrypanum) cruzi (Chagas, 1909), protozoário
pertencente à ordem kinetoplastida, é o agente etiológico da doença de Chagas (Tripanossomíase Americana), uma das mais importantes doenças parasitárias da América Latina (Schmuñis, 2000).
Nas últimas décadas, grandes avanços têm sido obtidos no controle das transmissões vetorial e transfusional da doença, principalmente através da Iniciativa do Cone Sul, o qual reduziu em aproximadamente 60% o índice da infecção humana (Dias
et al, 2002b; Schofield et al., 2006; Dias 2007). Embora esses programas de controle
tenham alcançado resultados satisfatórios, a transmissão ainda é ativa em outras partes do continente. Segundo dados da Organização Mundial de Saúde, aproximadamente 15 milhões de indivíduos estão infectados pelo T. cruzi na América Latina. A incidência
anual é de 41.200 novos casos e 12.500 mortes anuais ocorrem como conseqüência dos danos irreversíveis ao coração e ao trato digestivo (WHO, 2007). Dessa forma, a doença de Chagas é a maior causa de morbidade e mortalidade nos países onde é endêmica.
A infecção pelo T. cruzi determina no homem quadros clínicos com
características muito variadas (Dias 1992; Prata 1999; Macedo et al., 2004; Coura 2007;
WHO 2007). Após a infecção e um subseqüente período de incubação, tem-se início a fase aguda da doença de Chagas. Na ausência de tratamento específico, os sintomas podem persistir por até dois meses, com taxa de mortalidade variando de 2-8%, principalmente em crianças. Com a instalação da resposta imune do hospedeiro, as manifestações da fase aguda regridem e instala-se gradativamente a fase crônica, na qual a parasitemia e o parasitismo podem permanecer escassos por toda a vida do indivíduo (Dias, 1992). A forma crônica indeterminada é a mais freqüente, entretanto 30 a 40% dos pacientes chagásicos crônicos apresentam graus variáveis de miocardiopatia e 8 a 10% apresentam a forma digestiva, variando as formas clínicas e seu percentual entre países e, no interior de cada país, entre distintas áreas endêmicas (Dias, 1992).
Figueiredo, L.M. Introdução
3
de T. cruzi, não podendo descartar, contudo, um possível papel dos aspectos ambientais,
nutricionais genéticos e imunológicos do hospedeiro (Macedo & Pena, 1998; Devera et al., 2003; Macedo et al., 2004; Tibayrenc, 2007).
1.2) Variabilidade genética do T. cruzi
O T. cruzi é uma espécie heterogênea constituída por várias sub-populações do
parasito que circulam nos ambientes domésticos e silvestres, entre os seres humanos, animais reservatórios e vetores (Morel et al., 1986, Zingales et al., 1998). Variações
intra-específicas nesse parasito têm sido observadas com base no comportamento biológico das cepas em animais de laboratório (Silva & Nussenzweig, 1953; Andrade & Andrade, 1966; Andrade, 1985), vetores (Garcia & Dvorak, 1982; Lana et al., 1998) e
meios de cultivo celular e acelular (Dvorak et al., 1980; Sanchez et al., 1990; Laurent et al., 1997; Revollo et al., 1998) e essas diferenças estão associadas às características
bioquímicas e moleculares dos isolados (Bice & Zeledón 1970, Petana & Coura 1974, Miles et al., 1977, Melo & Brener 1978, Morel et al., 1980, Andrade 1985, Araújo &
Chiari 1988, Carneiro et al., 1991, Macedo & Pena 1998).
A variabilidade genética entre as populações de T. cruzi tem sido demonstrada
tanto em nível de proteína quanto de DNA. Em nível protéico, o primeiro estudo que demonstrou essa ampla diversidade genética foram os estudos iniciais de isoenzimas citoplasmáticas. Diferenças isoenzimáticas entre amostras do T. cruzi foram observadas
pela primeira vez por Toyé, (1974) utilizando as enzimas ASAT (aspartato aminotransferase) e ALAT (alanina aminotransferase). Miles et al., (1977, 1978, 1981)
estudando o perfil de isoenzimas em amostras de T. cruzi coletadas na Bahia e na região
Amazônica, encontraram três grupos isoenzimáticos distintos que foram denominados zimodemas Z1, Z2 e Z3. Estudos epidemiológicos demonstraram que Z1 e Z3 estavam associados ao ciclo silvestre e Z2 ao ciclo doméstico de transmissão da doença. Em seu trabalho posterior, Miles et al., (1981) demonstraram uma predominância do Z1 na
Figueiredo, L.M. Introdução
4
Romanha, (1982) e Carneiro et al., (1990) identificaram quatro perfis
eletroforéticos de isoenzimas analisados de amostras do T. cruzi isoladas de áreas
endêmicas das regiões sul e sudeste do Brasil que foram denominados de zimodemas A, B, C e D. A comparação desses zimodemas com os descritos por Miles et al., (1977,
1978, 1980) mostrou uma forte correlação entre Z2 e ZA.
Estudos subseqüentes realizados por Tibayrenc e Ayala, (1986) avaliando 15 locus enzimáticos em 121 amostras de T. cruzi de diferentes regiões geográficas, pertencentes
tanto ao ciclo silvestre quanto ao ciclo doméstico, identificaram 43 grupos genéticos distintos, os quais foram denominados clones ou genótipos naturais. Posteriormente, Tibayrenc e Brenière, (1988) verificaram que quatro desses genótipos ou “clonets” (19, 20, 39 e 32) se apresentavam em maior distribuição entre os hospedeiros e localidades geográficas distintas, constituindo 53,7% dos isolados e, por isso, representariam os genótipos principais do parasito. Esses autores sugeriram que esses estoques clonais ubíquos teriam grande importância epidemiológica para a doença de Chagas, merecendo estudos preferenciais, em diferentes aspectos. De fato, diversos trabalhos demonstraram a presença desses genótipos em diferentes áreas da América Latina: genótipos 19 e 39 na Bolívia, inclusive em infecções mistas em vetores e em humanos (Noireau et al.,
1995; Brenière et al., 1995; Brenière et al., 2002; Bastrenta et al., 2003); genótipos 19,
39 e 32 no Chile (Solari et al., 2001; Wallace et al., 2001; Coronado et al., 2006); e
genótipo 39 na Argentina (Solari et al., 1992). Entretanto, a prevalência desses
genótipos em outros países, inclusive no Brasil, ainda não foi determinada.
Tibayrenc & Ayala, (1988) propuseram que o T. cruzi possui estrutura e evolução
predominantemente clonal, observação que foi corroborada por vários autores empregando diferentes marcadores moleculares (Oliveira et al., 1998; Barnabé et al.,
2000; Brisse et al., 2000; Westenberger et al., 2005; Subileau et al., 2009). Tal modelo
Figueiredo, L.M. Introdução
5
Os polimorfismos de fragmentos de restrição do kDNA ou RFLPs (Restriction
Fragment Lenght Polymorphisms) foram os primeiros estudos de polimorfismo de DNA
descritos em T. cruzi (Morel et al., 1980). Essa metodologia foi proposta a partir da
análise do polimorfismo de sítios de restrição dos minicírculos por serem estes numerosos nesta espécie, conferindo elevada sensibilidade à metodologia e permitindo evidenciar a variabilidade intra-específica do kDNA exibidos por diferentes populações do parasito (Morel et al., 1980). No entanto, a classificação das amostras baseadas na
análise do perfil de restrição do kDNA (esquizodemas) necessita de amplificação prévia do isolado por cultura, podendo levar à seleção de sub-populações do parasito, a partir da população inicial presente no hospedeiro (Deane et al., 1984b).
Sturm et al., (1989) otimizaram essa metodologia pela realização da digestão
com enzimas de restrição diretamente do fragmento de 330pb, amplificado pela reação em cadeia da polimerase (PCR) e que continha as regiões variáveis do minicírculo. Essa estratégia aumentou a sensibilidade da técnica, possibilitando que o estudo de esquizodemas fosse realizado nas amostras biológicas, sem prévia expansão e manuseio em cultura.
Com a crescente evolução das técnicas de biologia molecular, diferentes metodologias têm sido também aplicadas para a genotipagem de cepas de T. cruzi em
nível de DNA genômico ou nuclear. Dentre elas podemos destacar a análise do polimorfismo de DNA amplificado aleatoriamente (RAPD-Random amplified polymorfic DNA) e a impressão digital do DNA (DNA fingerpriting). Steindel et al.,
(1993) empregaram essa metodologia para analisar diversas cepas isoladas no estado de Santa Catarina e demonstraram uma clara similaridade genética, detectada pelo RAPD, entre cepas de um mesmo zimodema. Similarmente, Tibayrenc et al., (1993) avaliaram
24 estoques clonais de T. cruzi previamente caracterizados por isoenzimas e observaram
paridade entre as duas técnicas além de demonstrar a convergência de isolados de T. cruzi para duas grandes subdivisões.
Já a técnica do DNA fingerpriting baseia-se na análise do DNA nuclear pela
utilização de sondas de mini-satélites humanos (DNA fingerprinting) e permitiu
Figueiredo, L.M. Introdução
6
A grande diversidade observada com os métodos descritos anteriormente não foi observada por Souto & Zingales, (1993) quando estudaram o gene que codifica o rRNA 24Sα em diversos isolados de T. cruzi. Utilizando a técnica de PCR, os autores
clonaram e seqüenciaram dois produtos distintos de diferentes isolados, com um fragmento de 125pb e com fragmento de 110pb, o que permitiu a classificação em dois grupos genéticos principais dentro da espécie T. cruzi: o grupo 1 e o grupo 2,
respectivamente. O aumento do número de amostras permitiu demonstrar a existência de um terceiro grupo de cepas que apresentavam os dois fragmentos (110 e 125 pb), correspondentes à perfis híbridos, sendo então denominados grupo 1/2 (Souto et al.,
1996). Posteriormente, Mendonça et al., (2002) e Pimenta (2002) ao analisar isolados
da Amazônia pertencentes ao Z3, através das seqüências do gene rDNA 24Sα
detectaram um fragmento de tamanho intermediário (117pb), correspondente a um novo produto de amplificação do rRNA.
Análises da seqüência do gene mini-exon também determinaram a divisão de cepas e isolados do T. cruzi em dois grupos: as cepas que apresentaram um produto de
300pb foram designadas como pertencentes ao grupo 1 (ou linhagem 1) e aquelas que apresentaram um produto de 350pb foram designadas como pertencentes ao grupo 2 (ou linhagem 2) – Souto et al., (1996).
Similarmente, Fernandes et al., (1998) estudaram as seqüências de genes de
mini-exons de isolados de T. cruzi obtidos de diferentes reservatórios, e verificaram a
existência de dois grupos genéticos distintos, linhagem 1 (associada principalmente com isolados humanos) e linhagem 2 (associada com o ciclo silvestre do parasito). Entretanto, algumas cepas não apresentavam produto de amplificação para o gene de mini-exon, apesar de apresentaram grande homologia com a seqüência do gene de cepas do grupo 2. Desse modo, essas cepas foram classificadas como Grupo 2’ e apresentaram correlação com o Z3 de Miles.
Ao analisar os dados obtidos pelos resultados das seqüências codificadoras para o rRNA e para o gene do miniexon na avaliação das relações filogenéticas entre os isolados do T. cruzi, Souto et al., (1996) propuseram a classificação do T. cruzi em três
Figueiredo, L.M. Introdução
7
o mini-exon e grupo 1/2 que apresenta ambos os produtos de amplificação do rRNA e do produto do grupo I do mini-exon.
O estudo do cariótipo do T. cruzi, demonstrou que a organização cromossomal
entre diferentes cepas é bem conservada, entretanto, o número e o tamanho das bandas cromossômicas separadas por PFGE (Pulse Field Gel Electrophoresis) apresenta grande
variação entre as cepas e clones podendo ocorrer, inclusive, variações entre os clones derivados de uma mesma cepa (Gibson & Miles, 1986; Vargas et al., 2004).
Clark & Pung, (1994) estudaram o polimorfismo do gene da subunidade 18S do rRNA, através da técnica de RFLP, e classificaram as cepas de T. cruzi em grupos
denominados ribodemas I, II e III. Foi observada uma forte correlação entre os marcadores genéticos para as subunidades 18S e 24Sα do rDNA, sendo as cepas
identificadas como ribodema I correspondentes ao grupo 1 e as cepas identificadas como ribodema II e III correspondentes ao grupo 2.
Embora análises da variabilidade intraespecífica do T. cruzi demonstrem
claramente uma ampla variabilidade biológica e genética desse parasito, as análises filogenéticas, empregando diferentes alvos no genoma do parasito, também revelam a tendência de agrupamento dos diferentes isolados em duas ou três subdivisões bem estruturadas.
Nesse sentido, na busca de um consenso sobre a classificação genética do T.
cruzi, e na uniformidade da nomenclatura empregada a fim de facilitar as correlações
entre os diferentes estudos, a comunidade científica durante o Encontro Satélite realizado em abril de 1999, no Rio de Janeiro, em comemoração ao 90º aniversário da descoberta da doença de Chagas, decidiu padronizar a nomenclatura dos dois grupos de
T. cruzi empregando a seguinte classificação (Anonymous, 1999):
i) cepas classificadas como Zimodema 1 (Miles et al., 1978), Tipo III (Andrade,
1974) Ribodemas II/ III (Clark & Pung, 1994), Grupo 1 (Tibayrenc, 1995) e linhagem 2 (Souto et al., 1996) foram renomeadas T. cruzi I.
ii) cepas classificadas como Zimodema 2 (Miles et al., 1978), Zimodema A
Figueiredo, L.M. Introdução
8
Grupo 2 (Tibayrenc, 1995) e linhagem 1 (Souto et al., 1996) foram designadas T. cruzi
II.
iii) cepas caracterizadas híbridas ou cuja caracterização seja inconclusiva, dentre as quais o Zimodema 3 (Miles et al., 1978), Zimodema B (Romanha et al., 1979)
Zimodema Chileno 2b (Miles et al., 1984), Tipo 1 (Andrade, 1974) o grupo 1/2 (Souto et al., 1996), genótipo 39 (Tibayrenc, 1995) foram nomeadas T. cruzi.
Entretanto, o desenvolvimento de novos marcadores multilocos para o estudo da diversidade genética do T. cruzi evidenciou a existência de mais subdivisões em T. cruzi
(Barnabé et al., 2000a; Brisse et al., 2000; Brisse et al., 2001; Westerbeng, 2005;
Subileau et al., 2009). Como já citado anteriormente, Tibayrenc, (1993) verificaram
pelo perfil de isoenzimas e RAPD, que o T. cruzi pode ser subdividido em dois grandes
que foram denominados de DTU1 e DTU2 (Discrete Typing).
Posteriormente, a fim de melhor entender a estrutura populacional do T. cruzi II
Brisse et al., (2000) utilizando análise de MLEE (Multilocus Enzyme Electrophoresis) e
RAPD (Randomly Amplified Polimorphic DNA) estudaram um grande número de
isolados do parasito e propuseram a sua subdivisão em cinco sublinhagens discretas denominadas DTUs 2a, 2b, 2c, 2d e 2e. A DTU2b corresponde ao Zimodema 2 (Miles
et al., 1977), as DTU 2c ao Zimodema 3 (Miles et al., 1977) e as DTUs 2d e 2e são
linhagens híbridas estabelecidas após subseqüente propagação clonal. Nesse estudo, os autores ainda sugeriram que as DTUs IId e IIe foram consideradas produtos de um evento ancestral de hibridização entre DTUs IIb and IIc (Brisse et al., 2000a,b; Barnabé et al., 2000, 2003).
Ao estudar esses mesmos subgrupos do T. cruzi, (Westenberger et al., 2005)
Figueiredo, L.M. Introdução
9
Variações intraespecíficas do T. cruzi também tem sido demonstradas por meio
da análise genes que codificam proteínas e rRNA mitocondriais (Freitas et al., 2006,
Westenberger et al., 2006). Avaliando genes mitocondriais, Freitas et al., (2006)
sugeriram a existência de um terceiro grupo ancestral denominado T. cruzi III e que por
eventos de hibridização desse com o T. cruzi II, teria originado os isolados de caráter
híbrido que são observados atualmente. Desta maneira, as populações do T. cruzi seriam
subdivididas em T. cruzi I, T. cruzi II e T. cruzi III (que corresponde ao Zimodema 3 e a
DTU2c e os híbridos (que correspondem às DTU(s) 2d e 2e.
A busca de uma correlação entre a variabilidade genética do T. cruzi e as
diferentes manifestações clínicas na doença de Chagas, tem estimulado os pesquisadores a desenvolverem novas metodologias capazes de detectar alvos do parasito diretamente no sangue ou de tecidos provenientes do hospedeiro infectado
Os microssatélites, que correspondem a pequenas seqüências em tandem de DNA
com motivos de repetição de dois até seis nucleotídeos apresentam um elevado grau de polimorfismo em relação ao número de repetições de um dado loco e, portanto, são bastante úteis para o estudo da variabilidade genética do parasito. A primeira técnica a empregar esse alvo no genoma do parasito foi proposta por Oliveira et al., (1997) e foi
denominada de SSR-PCR (Simple Sequence Repeat-anchored PCR). Os autores
realizaram a amplificação simultânea de vários locos entre as repetições (CA)n de microssatélites, produzindo um perfil de múltiplas bandas. Posteriormente, esses mesmos autores caracterizaram oito locos polimórficos contendo microssatélites em diversas cepas de T. cruzi (Oliveira et al., 1998) e demonstraram sua importância para a
reconstrução filogenética deste parasito. Além disso, Pimenta, (2002) demonstraram que os microssatélites são marcadores com um enorme poder de resolução.
Recentemente, Valadares et al., (2008) demonstraram que as análises de
microssatélites também apresentam boa estabilidade sob condições de laboratório, além de uma elevada sensibilidade. Dessa forma, essa técnica é adequada para detectar o T. cruzi em amostras biológicas mesmo quando esse está presente em baixa quantidade,
dispensando o cultivo e a manipulação do parasito, o que poderia levar a diferenças do isolado original. Entretanto, Martins et al., (2008) observaram que a técnica de
Figueiredo, L.M. Introdução
10
sangue e em tecidos de animais com infecção mista atribuindo esse insucesso à baixa carga parasitária, o que diminuiria a sensibilidade da técnica.
Dessa forma, outra abordagem deve ser empregada para caracterizar o T. cruzi
diretamente de amostras biológicas que apresentam baixa quantidade do parasito. Neste sentido, a técnica de LSSP-PCR (Low-stringency Single Primer PCR), que utiliza um
único iniciador complementar a um sítio específico do DNA molde, ao hibridizar com diversos sítios de maneira independente, é capaz de gerar um padrão complexo de fragmentos que constitui uma assinatura gênica, além de apresentar elevada sensibilidade (Pena et al., 1994). Essa metodologia foi a primeira capaz de detectar a
variabilidade do kDNA do T. cruzi diretamente de tecidos infectados sem necessitar de
isolamento prévio das cepas infectantes, evitando a seleção de uma sobre a outra.
Vago et al., (1996b, 2000) utilizaram essa mesma metodologia e avaliaram o
tropismo tecidual do T. cruzi diretamente no coração e no esôfago de 15 pacientes
chagásicos crônicos. Nesse estudo, foi demonstrado que a caracterização das populações do T. cruzi, presentes nos tecidos do coração e do esôfago, revelou diferentes perfis de
assinaturas gênicas nestes órgãos, sugerindo que a variabilidade genética do parasito pode determinar diferentes tropismos, exercendo assim papel importante na evolução das diferentes formas clínicas da doença de Chagas. Por outro lado, Lages-Silva, (2001) utilizando essa mesma técnica, demonstrou identidade nos perfis de LSSP-PCR das populações do parasito isoladas do sangue por hemocultura e/ou pelo vetor de um mesmo paciente cujos perfis foram altamente representativos das populações teciduais.
Lages-Silva et al., (2006) utilizando a técnica de LSSP-PCR para caracterizar
cepas de T. cruzi obtidas de pacientes chagásicos, demonstraram um alto grau de
variabilidade intra-específica entre os isolados, sendo únicos e específicos para cada paciente. Entretanto, esses autores não puderam correlacionar essa variabilidade genética com as características clínicas apresentadas pelos pacientes.
Uma grande heterogeneidade genética também foi verificada por Salazar et al.,
(2006), ao utilizarem a técnica de LSSP-PCR para caracterizar isolados de T. cruzi
Figueiredo, L.M. Introdução
11
Recentemente, Brito et al., (2008) estudaram a variabilidade genética de cepas
de T. cruzi isoladas de fezes de triatomíneos capturados do nordeste brasileiro através da
técnica de LSSP-PCR. Os autores verificaram a presença de diferentes genótipos pertencentes ao T. cruzi I circulando entre diferentes espécies de vetores.
No entanto, mesmo com os grandes avanços obtidos em relação ao desenvolvimento de metodologias capazes de detectar polimorfismos em diferentes alvos no genoma do T. cruzi, o estabelecimento de correlações entre a genética do
parasito e suas características biológicas, epidemiológicas e clínicas da doença de Chagas ainda é difícil de ser demonstrado. Possivelmente, fatores do hospedeiro devem ser considerados para um maior entendimento desta íntima relação parasito-hospedeiro.
1.3) Correlação entre a diversidade genética do T. cruzi e suas propriedades biológicas
Diversos trabalhos utilizando diferentes modelos experimentais mostraram que o T. cruzi apresenta uma grande variabilidade nas suas propriedades biológicas, inclusive no
tropismo tecidual e na susceptibilidade aos nitroderivados (Chagas, 1909; Vianna, 1911; Brener & Chiari, 1963; Melo & Brener, 1978; Bice & Zeledon, 1970; Andrade, 1974; Brener et al., 1976; Filardi & Brener, 1987; Lenzi et al., 1998).
Considerando que o T. cruzi apresenta estrutura e evolução predominantemente
clonal é esperada uma evolução pareada entre a divergência filogenética de diferentes clones e genes que dirigem importantes propriedades do parasito relacionadas à virulência, patogenicidade e a epidemiologia da doença. Portanto, essa evolução não-independente implicaria em uma forte correlação entre a variabilidade genética do parasito e essas propriedades (Tibayrenc, 2003). Dessa forma, os genótipos de T. cruzi
filogeneticamente próximos ou aparentados, teriam propriedades biológicas ou clínicas semelhantes e os genótipos filogeneticamente distantes apresentariam propriedades mais distintas entre si (Tibayrenc et al., 1986; Tibayrenc & Ayala, 1988).
Figueiredo, L.M. Introdução
12
populações pertencentes aos genótipos 19 e 20, filogeneticamente próximos e correspondentes ao grupo T. cruzi I (Anonymous, 1999) apresentaram maior habilidade
em infectar e completar seu ciclo de vida no inseto vetor (Garcia & Dvorak, et al., 1982;
Garcia & Azambuja, 1991; Lana et al., 1998), maior capacidade de realizar a
metaciclogênese, maior capacidade de multiplicação no hospedeiro vertebrado (Andrade, 1974; Andrade et al., 1983; Sànchez et al., 1990; Gonzalez et al., 1995;
Andrade & Magalhães, 1997; Wallace et al.; 2001; Toledo et al., 2002) e maior
capacidade de multiplicação e diferenciação em meios de cultura celular e acelular (Laurent et al., 1997; Revollo et al., 1998) do que as populações pertencentes aos
genótipos 39 e 32, correspondentes ao grupo T. cruzi II, geneticamente mais
relacionados entre si e distantes dos genótipos 19 e 20 que apresentaram características opostas em relação a todos estes parâmetros analisados.
Com relação à susceptibilidade a fármacos, foi demonstrado em diversos trabalhos, que cepas do grupo T. cruzi I apresentaram elevado grau de resistência, tanto
em cultivo celular como em camundongos, enquanto as cepas do grupo T. cruzi II foram
mais susceptíveis (Andrade et al., 1985a; Andrade & Magalhães, 1997, Revollo et al.,
1998; Toledo et al., 2003). Por outro lado, Villareal et al., (2004) não conseguiram
estabelecer correlação entre susceptibilidade ao BZ e distância filogenética do parasito
in vitro. Estes resultados analisados em conjunto, enfatizam a crucial importância de
considerar a diversidade filogenética dos clones naturais do T. cruzi em todos os estudos
referentes a biologia e susceptibilidade aos quimioterápicos, pois tem sido observada uma grande variabilidade entre os clones de um mesmo genótipo (Laurent et al., 1997;
Revollo et al., 1998; Toledo et al., 2003, 2004; Martins et al., 2006, 2007). Estes
resultados sugerem a necessidade de considerar subgrupos menores do parasito ou a utilização de marcadores moleculares mais discriminantes para melhor avaliar a associação entre a genética do parasito e suas características biológicas, de susceptibilidade a fármacos e clínicas.
Ao investigar a associação entre divergência filogenética e resposta à quimioterapia, Toledo et al., (2003, 2004) demonstraram importantes diferenças na
sensibilidade a quimioterápicos entre essas mesmas populações de T. cruzi. Esses
Figueiredo, L.M. Introdução
13
ao tratamento, enquanto clones pertencentes ao grupo genético T. cruzi II (genótipos 32
e 39) foram susceptíveis ou parcialmente susceptíveis. Foi demonstrado ainda que a resistência observada para T. cruzi I foi devido aos isolados pertencentes ao genótipo 20
(100% de resistência), já que o genótipo 19 inclui clones sensíveis, parcialmente e totalmente resistentes ao BZ. Os diferentes graus de susceptibilidade ao BZ observado para clones pertencentes ao grupo T. cruzi II se deve ao fato que nesse grupo estão
incluídos clones resistentes ou susceptíveis ao tratamento (genótipo 39) e clones que apresentam um perfil típico de susceptibilidade (genótipo 32).
A correlação entre divergência filogenética e propriedades biológicas foi particularmente demonstrada por Toledo et al., (2002) ao estudar estoques clonais do T.
cruzi. Os autores utilizaram camundongos BALB/c inoculados com clones pertencentes
aos genótipos principais do T. cruzi (19, 20, 39 e 32) e avaliaram comparativamente 14
parâmetros durante as fases aguda e crônica da infecção. Os resultados obtidos demonstraram que os genótipos mais relacionados entre si (19x20 ou 32x39) apresentaram um menor número de diferenças significativas do que os genótipos geneticamente distantes (19 ou 20 versus 32 ou 39), exceto para a comparação 19x32.
Clones do genótipo 20 foram os mais virulentos e patogênicos e na fase aguda da infecção, enquanto clones do genótipo 39 foram os únicos nos quais se detectou parasitismo tecidual durante a fase crônica.
Ao estudar o impacto da variabilidade genética de clones pertencentes aos genótipos principais do T. cruzi (19, 20 e 39), Diego et al., (1998) verificaram uma
correlação entre distância genética e achados histopatológicos em camundongos Swiss. Os animais infectados com o genótipo 20 apresentaram comprometimento encefálico correspondendo a áreas de inflamação e necrose. Foi observado também parasitismo no baço e fígado apenas em animais infectados com o genótipo 39. O tropismo para o músculo esquelético foi acentuado para todos os genótipos e mais intenso que o cardiotropismo, que por sua vez apresentou diferenças entre todos os genótipos.
Apesar dos diversos clones de T. cruzi apresentarem um tropismo preferencial
para determinados tipos celulares, a correlação entre a variabilidade genética do T. cruzi
Figueiredo, L.M. Introdução
14
parasito na fase crônica da infecção, a fim de estabelecer uma correlação entre o perfil de RAPD dos isolados e as diferentes formas clínicas apresentadas pelos pacientes. Apesar da grande similaridade entre isolados, indicando a presença de grupos genéticos bem correlacionados, os autores não conseguiram estabelecer essa correlação.
Apesar desses diversos genótipos principais (Tibayrenc e Brenière, 1988) apresentarem características peculiares em diversos parâmetros biológicos, na resposta à quimioterapia, bem como no tropismo tecidual, a associação entre esses grupos genéticos do T. cruzi, além das características genéticas do hospedeiro parecem resultar
em importantes alterações no curso da infecção chagásica (Lana et al., 2000; Martins et al., 2006).
1.4) Genética do parasito e distribuição tecidual do T. cruzi
A distribuição tecidual preferencial de cepas do T. cruzi em humanos e animais
experimentais tem sido discutida por muitos autores desde o início do século. Viannia, (1911) verificou que o T. cruzi poderia ser encontrado em diferentes tecidos do
hospedeiro vertebrado, embora os parasitos apresentassem certa preferência por determinados grupos celulares.
Diferenças no tropismo tecidual foram observadas por Brener, (1977) e Melo & Brener, (1978) em camundongos inoculados com diferentes cepas (Y, Berenice, ABC e CL) quando verificaram parasitismo preferencial para dois sistemas de células, as fagocitárias do baço, fígado e medula óssea (cepas Y e Berenice) e células musculares lisa, esquelética e cardíaca (cepa ABC e Berenice).
Diversos autores têm demonstrado que as populações do T. cruzi são
multiclonais, constituídas por populações heterogêneas (Macedo et al., 2002, 2004) e
Figueiredo, L.M. Introdução
15
inabilidade de competir e se propagar no ambiente humano. Dessa forma, o hospedeiro funcionaria como um filtro biológico, diminuindo a complexidade da cepa infectante. Entretanto, os clones remanescentes poderiam apresentar tropismos para diferentes tipos celulares e, como conseqüência, determinar o curso clínico da doença. Este cenário levou a criação do “Modelo Histiotrópico Clonal” da doença de Chagas (Macedo & Pena,1998).
Essa variação no tropismo tecidual de diferentes populações do T. cruzi também
foi demonstrada por Andrade et al., (1999) que, ao infectarem camundongos BALB/c
simultaneamente com duas diferentes populações do parasito (a cepa JG, isolada de um paciente com megaesôfago e oclone CoL1.7G2, isolado do sangue de um paciente com a forma cardíaca) demonstraram uma clara diferença na distribuição tecidual das populações do parasito tanto nas infecções mistas quanto nas infecções monoclonais. Após três meses de infecção, o clone CoL1.7G2 predominou sobre a cepa JG no reto, diafragma, esôfago e no sangue, enquanto a cepa JG foi encontrada no músculo cardíaco do mesmo animal. Esses resultados não apenas fornecem evidências do tropismo tecidual de cepas na fase crônica da infecção, mas também demonstraram a importância de se considerar a variabilidade genética do hospedeiro na distribuição tecidual do parasito.
Apesar dos diversos clones de T. cruzi possuírem um tropismo tecidual para
órgãos específicos, vários mecanismos dependentes do hospedeiro podem também estar envolvidos no processo de distribuição tecidual diferenciado do parasito (Russo et al.,
1996).
1.5) Genética do hospedeiro e distribuição tecidual do T. cruzi
Ao abordar aspectos genéticos do hospedeiro como um dos fatores determinantes das formas clínicas da doença de Chagas, Andrade et al., (2002) estudaram o papel de
genética de camundongos na distribuição tecidual de populações clonais do T. cruzi. Os
autores compararam as infecções experimentais entre animais das linhagens BALB/c e
DBA-2 (ambos haplótipo 2d), C57BL/6 (haplótipo 2b) e Swiss infectados com o clone Col1.7G2 e com a cepa monoclonal JG. Os camundongos BALB/c e DBA-2 infectados
Figueiredo, L.M. Introdução
16
tecidual do parasito, com a cepa JG predominando no coração e o clone CoL1.7G2 predominando no reto e em outros tecidos. Entretanto, o mesmo padrão de distribuição não foi observado para os camundongos da linhagem C57BL/6 e Swiss. Nos animais da linhagem C57BL/6, o clone CoL1.7G2 foi detectado em quase todos os tecidos analisados, inclusive no coração, semelhante ao observado para os camundongos Swiss, com exceção do sangue e reto, onde predominou a cepa JG, indicando a importância de aspectos genéticos do hospedeiro no padrão de distribuição tecidual de populações do
T. cruzi.
Embora a exata causa dessa variabilidade clínica ainda deva ser elucidada, diversos trabalhos indicam que a região do MHC do hospedeiro modula o dano tecidual, a sua sobrevivência (Wrightsman et al., 1984) e a susceptibilidade/resistência à infecção
pelo T. cruzi no modelo murino (Trischmann et al., 1982), já que a interação do MHC
com o parasito poderia influenciar na resposta humoral contra este parasito (Aguillon et al., 2000) e na indução de apresentação de antígenos à células TCD8+ (Van Overtvelt et al., 2002) através da modulação da expressão de moléculas pelo T.cruzi (Alba Soto et al., 2003).
Apesar de alguns autores terem demonstrado nenhuma ou pouca influência do MHC no desenvolvimento das diferentes formas clínicas da doença de Chagas em pacientes do Brasil (Fae et al., 2000), estudos em indivíduos com infecção pelo T. cruzi
do México sugeriram que alguns alelos do MHC poderiam estar associados com as formas clínicas da doença crônica, especialmente com maior risco de desenvolver a doença cardíaca (Cruz-Robles et al., 2004). Recentemente, Freitas et al., (2009)
estudaram a doença de Chagas no modelo murino e demonstraram que a região do MHC do hospedeiro influencia o padrão de distribuição tecidual de cepas de T. cruzi, o
qual poderia, na infecção humana, contribuir para a determinação das formas clínicas da doença de Chagas. Os autores demonstraram que o clone CoL1.7G2 predominou no coração de animais C57BL/6 (H-2b), enquanto a cepa JG foi a que predominou no coração de camundongos da linhagem C57BLKS/J (haplótipo H2-d).
Outros trabalhos ainda demonstraram que tanto o parasito quanto o hospedeiro têm um importante papel no desenvolvimento da infecção não só na fase crônica, mas também na fase aguda da doença. (Andersson et al., 2003) demonstraram que