RNA extraction, RT and Real-Time PCR (qPCR)
Texto
Imagem

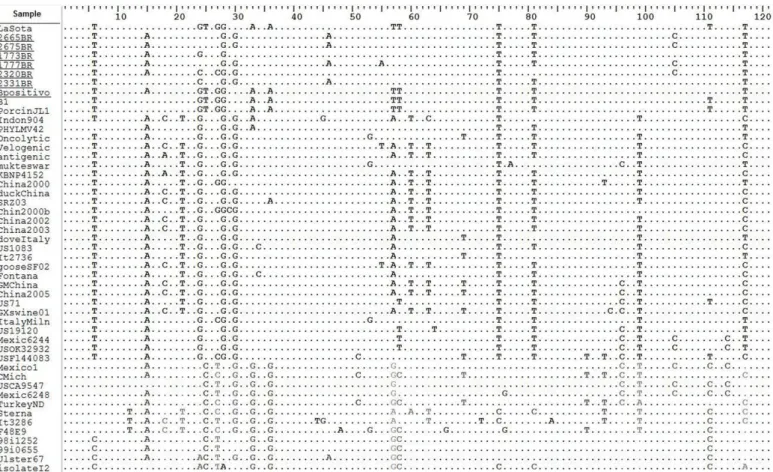
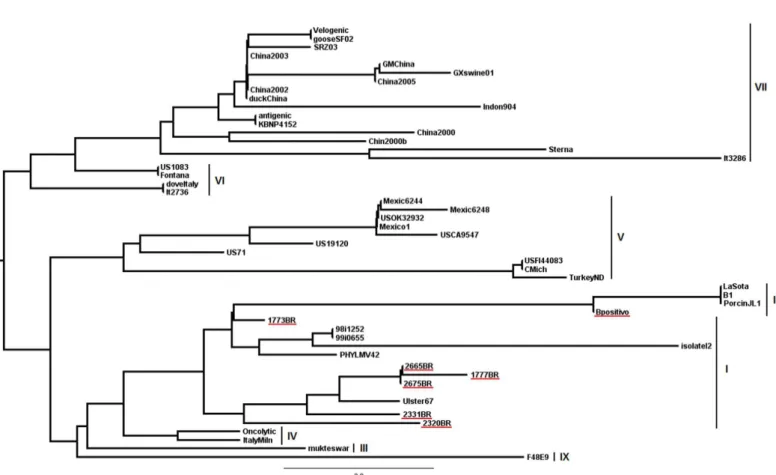
Documentos relacionados
quantified using quantitative RT-PCR technique. After treatment mRNA was isolated from the tissues using RNA extraction kit followed by quantitative RT-PCR. Total extracted RNA
Real-time RT-PCR analyses of genes cryD , wcoA , vvdA and carRA in total RNA samples from the wild type grown for three days in DGasn medium in the dark or after 15 min, 30 min, 1 h,
We evaluated primers and probes to detect Zika virus, which had been published before, and tested sensitivity using serum spiked and patient samples by real-time RT-PCR.. When
Blood samples were collected from 99 domestic dogs from the urban and rural areas of the Lábrea municipality, state of Amazonas, Brazil.. Canine serum samples were tested
Blood and serum samples collected by convenience from dogs in urban areas of five municipalities were analyzed by real-time PCR for the detection of E. canis.. DNA and
Using the kit that showed better performance, two systems of Real- time RT-PCR (RT-qPCR) assays were tested and compared for analytical sensitivity to Canine Distemper Virus
Figure 2 - Effect of drying rate on primary root protrusion (a) , percentage of normal seedlings (%) (b) and germination speed index (GSI) (c) of Campomanesia adamantium
Methods: Oropharyngeal swabs, cloacal swabs, and blood samples from 170 birds captured in two green areas in São Paulo, Brazil were subjected to real time - polymerase chain
