Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Vegetal
DROUGHTYRUS – PRELIMINARY CHARACTERIZATION OF
THE PHYSIOLOGICAL RESPONSES OF TEN LATHYRUS
GENOTYPES TO WATER DEFICIT
Susana Margarida Duarte Silvestre
Dissertação de Mestrado em Biologia Celular e Biotecnologia
2011
Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Vegetal
DROUGHTYRUS – PRELIMINARY CHARACTERIZATION OF
THE PHYSIOLOGICAL RESPONSES OF TEN LATHYRUS
GENOTYPES TO WATER DEFICIT
Susana Margarida Duarte Silvestre
Dissertação de Mestrado sob a Orientação de:
Professor Doutor Jorge Marques da Silva e
Doutoura Susana de Sousa Araújo
Dissertação de Mestrado em Biologia Celular e Biotecnologia
2011
iii
AGRADECIMENTOS
Tenho a agradecer em primeiro lugar os meus orientadores Professor Doutor Jorge Marques da Silva e Doutora Susana Araújo não só pela possibilidade da realização deste trabalho, mas também por todo o apoio demonstrado ao longo deste ano. A tudo o que me ensinaram, o que me corrigiram e pela liberdade com que me deixarm guiar o meu trabalho. Quero agradecer especialmente à Dra. Maria Carlota Vaz Patto por toda a sua dedicação e entusiasmo no acompanhamento deste projecto. Obrigada pela sua enorme paciência para me ajudar com a estatística dedicando o seu tempo à minha causa.
Tenho a agradecer à Manuela Lucas (ou Manecas) pela ajuda técnica, psicológica e sentimental que me dedicou incansávelmente durante todo este ano e sem deixar que me desanimasse.
Agradeço também à Professora Doutoura Cristina Cruz e Professora Doutoura Otília Correia. pela disponibilização de equipamentos e pelo apoio cientifico na resolução de problemas.
Obrigada a todos os meus amigos biólogos, físicos, matemáticos e economistas.
Aos meus caros colegas de mestrado pelo acompanhamento e amizade destes dois anos.
Às minhas recentes aquisições do ITQB, a piquena Silvestre agradece.
Ao André Alcântara (desculpa mas não tive tempo de pôr aqui um desdobrável com a tua cara, mas fica a intenção) um grande obrigado por toda a ajuda e infinita paciência de me aturar nos meus momentos de irritação e frustração.
Um obrigada muito especial ao meu grande amigo Leo.
Quero agradecer também a toda a minha família, e em especial aos meus pais, que sempre me apoiaram e me deram força.
iv
RESUMO
As leguminosas do género Lathyrus têm grande potencial agronómico não só como espécies produtoras de grão mas também como forrageiras. Salienta-se o Lathyrus sativus considerada como a principal cultura e fonte de proteína vegetal de populações de regiões áridas, nomeadamente por ser a única espécie capaz de crescer em tais condições. No entanto, esta espécie acumula uma neurotoxina (um aminoácido não proteico -ODAP) cuja acumulação em organismos animais resulta em paralisias irreversíveis. Embora alguns estudos tenham demonstrado uma relação entre o teor em -ODAP e as respostas ao deficit hídrico, ainda existe um desconhecimento da sua verdadeira função nas plantas.
Neste trabalho foram utilizados vários genótipos do género Lathyrus, entre eles L. aphaca (L375), L. cicera (L94), L.s clymenum (L388), L. latifolius (L563), L. ochrus (L313), L. odoratus (L17-1), L. sativus (L174-A) and L. sylvestris (L531). Todos eles cedidos pelo Centro de Recursos Fitogenéticos (CRF/INIA), Madrid, Spain pelo Professor Diego Rubiales (IAS-CSIC, Córdoba, Spain). L. sativus (LS 87124), uma variedade com baixas concentrações de -ODAP (0,02%), e L. sativus (Raipur), uma variedade da região de Raipur na India e descrito como tendo concentrações altas de -ODAP (0,54%) foram obtidas pelo Professor Lambein (IPBO, Ghent, Belgium).
Foram usadas dez plantas para cada um dos dez genótipos de Lathyrus sp. caracterizadas em condições de irrigação não-limitante e em condições de deficit hidrico. L. sylvestris e L. sativus (LS 87124) foram as excepções que, por terem mostrado baixas taxas de germinação de 13% e 21% respectivamente, foram representadas apenas por duas e oito plantas respectivamente. Durante o tempo de crescimento, que durou 3 meses e meio, em estufa, as plantas foram regadas sempre que necessário.
A caracterização dos genótipos em condições de irrigação não limitante foi baseada em medições do teor hídrico relativo das plantas (na sigla inglesa RWC), das taxas de evapotranspiração, da clorofila (determinação pelo SPAD), da reflectância foliar e medições da cinética rápida de indução de fluorescência da clorofila a. As condições de stress por deficit hídrico foram induzidas pela supressão da rega a cinco das dez plantas por cada genótipo, em que o último dia de rega, considerado como o dia zero, marca o início deste período. O estudo das respostas fisiológicas correspondentes a este período foi baseado na análise do teor hídrico relativo das plantas, do conteúdo hídrico do solo (na sigla inglesa SWC), da reflectância foliar e da fluorescência da clorofila a.
Em condições de irrigação não limitante o teor hídrico relativo das plantas mostrou-se alto com valores entre 85,8% (± 6,0) a 94,0% (± 5,1), em que as diferenças entre genótipos eram mínimas excepto entre L. cicera (88,4% ± 2,7) e L. odoratus (92,5% ± 4,2). Nas taxas
v
de evapotranspiração (massa de água (g) perdida por hora para cada planta envasada, g H2O.h-1) L. clymenum mostra a maior taxa na ordem dos 1,327 gH2O.h-1 (± 0,356) ao
contrário de L. aphaca (0,580 g H2O.h-1 ± 0,092) e de L. latifolius (0,556 g H2O.h-1 ± 0,091).
Todos os genótipos de L. sativus mostraram também elevadas taxas de evapotranspiração, sugerindo uma menor adaptação a possíveis cenários de aridez.
A análise fotoquímica foi realizada através da medição da cinética rápida de indução da fluorescência da clorofila a, que resulta numa série de parâmetros que traduzem a performance do aparelho fotossintético da planta em várias situações. Estes incluem o rendimento quântico máximo do fotossistema II (Fv/Fm) e o índice de performance (com a single inglesa PI), que reflecte a funcionalidade de ambos os fotossistemas e transmite uma informação quantitativa do desempenho da planta em condições de stress. Incluem ainda a área complementar (na sigla inglesa CAREA) entre a curva de indução de fluorescência e o
valor de fluorescência máximo, que nos dá uma medida do tamanho do pool de quinonas aceitadoras de electrões (na sigla inglesa QA) do fotossistema II. Com valores mais
elevados de Fv/Fm destacam-se L. sativus (L174-A) (0,827 ± 0,012) e L. cicera (0,823 ± 0,015). Por outro lado L. ochrus e L. latifolius apresentam os menores valores com 0,792 (± 0,034) e 0,791 (± 0,066), respectivamente. Em termos de PI podemos agrupar os genótipos em três classes distintas: L. sativus (L174-A) (3,570 ± 1,144), L. cicera (3,053 ± 1,221) e L. latifolius (3,165 ± 2,028) com valores mais altos; L. clymenum (2,506 ±0,883), L. odoratus (2,297 ±0,671) e L. sylvestris (2,704 ±0,978) pertencentes a um grupo intermédio, enquanto que com valores mais baixos encontram-se L. sativus (LS 87124) (3,570 ± 1,144), L. sativus (Raipur) (2,225 ± 1,150) e L. ochrus (1,613 ± 0,768). Em relação à CAREA L. sativus (L174-A)
(44983 ± 10457) e L. ochrus (24085 ± 10683) seguem o mesmo padrão observado para Fv/Fm e PI, enquanto que todos os restantes genótipos mostraram valores intermédios.
A partir da medição da reflectância foliar foram calculados quatro índices de reflectância, o índice de reflectância hídrico (na sigla inglesa WRI) relacionado com o conteúdo hídrico foliar, o normalized difference vegetation index (NDVI) usado como estimativa indirecta da capacidade fotossintética e da produção primária. O índice de reflectância fotoquímico (na sigla inglesa PRI) relacionado com processos de dissipação de excesso de radiação e também o índice de conteúdo de clorofila (na sigla inglesa CHLI) permitindo uma estimativa da concentração de clorofila de forma não destrutiva. Para todos estes índices o genótipo L. sylvestris destacou-se pelos seus altos valores, embora nos casos do NDVI e PRI seja acompanhado por L. cicera (0,826 ± 0,042 and 0,034 ± 0,007, respectivamente) e para o WRI por L. sativus (Raipur) (1,00 ± 0,01). Nos índices CHLI e NDVI destacam-se L. ochrus (2,06 ± 0,66 and 0,584 ± 0,252, respectivamente) e L. aphaca (2,14 ± 0,53 and 0,682 ± 0,201, respectivamente). Para PRI L. ochrus (0,007 ± 0,020) é acompanhado por L. latifolius (0,017 ± 0,019) e para o caso de WRI apenas L. aphaca (1,00 ± 0,01) apresenta os valores
vi
mais baixos. A medição da clorofila pelo SPAD resultou na definição de três grupos e mostrou-se em conformidade com o CHLI, validando-se não só os métodos entre si mas também às diferenças estabelecidas entre genótipos.
Para o estudo das respostas a condições de deficit hídrico definiu-se um ponto máximo de stress por deficit hídrico (na sigla inglesa PMWS) a partir do qual as plantas foram novamente regadas, a definição deste ponto foi feita inicialmente com base na avaliação do RWC até aos 30% fazendo com que alguns dos genótipos atingissem níveis de stress severo a partir dos quais não houve recuperação. Posto isto a avaliação do PMWS foi feita com o auxílio do Fv/Fm para valores iguais ou inferiores a 0,75. A análise dos valores de fluorescência no PMWS mostram diferenças significativas quando comparadas com os valores de controlo. Tendo em conta os baixos valores de PI registados para este ponto podemos ver que para todos os genótipos estudados o stress imposto foi bastante severo resultando não só em danos significativos no sistema fotossintético mas também na perda de vitalidade das plantas.
Para todos os genótipos estudados o Fv/Fm mantém-se estável mostrando valores semelhantes aos de controlo para RWC relativos a um stress hídrico moderado (acima dos 70%). Nos casos de stress severo em que o RWC atinge valores iguais ou inferiores a 30% há um decréscimo acentuado do Fv/Fm. Nos casos do PI e da CAREA o decréscimo dos seus
valores é gradual acompanhando o do RWC, até que mais uma vez atingindo-se os 30% de RWC verifica-se uma diminuição acentuada. Quanto à reflectância foliar os índices calculados não mostraram resultados significativos, não permitindo uma conclusão sobre a sua relação com o RWC.
Os genótipos L. sativus (L174-A) e L. sativus (Raipur) destacaram-se pela sua performance ao longo do decréscimo do RWC, mesmo em classes de stress severo variando entre os 40% e 20% de RWC. Esta performance é destacada pela manutenção de valores significativos de Fv/Fm, PI e CAREA. Estes resultados dão-nos a confirmação
fisiológica da tão reconhecida resistência destas espécies de cultura em zonas áridas. No entanto estudos adicionais são necessários de forma a estabelecer uma linha de base na selecção dos genótipos mais apropriados para cultivo em regiões em que a seca é um problema grave e recorrente.
vii
ABSTRACT
Some species of the Lathyrus genus are important local legume crops providing proteins where products of animal origin are unavailable. Furthermore, some of them are drought resistant rendering them appropriate for areas were irrigation is not available. Nevertheless, the mechanisms underlying water deficit stress resilience of these species are not fully understood, and thereby the main objectives of this work were the characterization of ten Lathyrus sp. genotypes both in well watered conditions and in water deficit stress.
Genotypes studied included L. aphaca (L375), L. cicera (L94), L. clymenum (L388), L. latifolius (L563), L. ochrus (L313), L. odoratus (L17-1), , L. sylvestris (L531), L. sativus (L174-A), L. sativus (LS 87124) and L. sativus (Raipur). Ten plants for each genotype were grown in a greenhouse for 3 ½ months and were watered whenever necessary. Characterization plant and soil water status was performed by the assessment of soil water content (SWC), relative water content (RWC) and evapotranspiration rate. Chlorophyll content (SPAD), foliar reflectance and chlorophyll a fluorescence were used to characterize the photochemical performance. Results shows that at well watered conditions L. sativus (L174-A), L. sativus (LS 87124) and L. sativus (Raipur) have high evapotranspiration rates. Concerning Fv/Fm measurements all genotypes presented values close to 0.8, with the higher values showed by L. sativus (L174-A) (0.827 ± 0.012) and the lowest presented by L. ochrus (0.792 ± 0.034) and L. latifolius (0.791 ± 0.066). Again L. sativus (L174-A) along with L. cicera and L. latifolius showed higher Performance Index (PI) in contrast with both L. sativus (LS 87124) and L. sativus (Raipur). Reflectance indices showed L. sylvestris with the best performance results.
All genotypes showed resistance to mild water stress, maintaining unaltered values of physiological parameters until RWC decreases to 60 – 70%. At severe water stress (20 to 30% RWC) two genotypes L. sativus (L174-A) and L. sativus (Raipur) showed remarkable resilience preserving the functionality of both photosystems and maintaining the PS II efficiency. The foliar reflectance measured had no conclusive results.
viii
ABBREVIATIONS
- ODAP: -N-Oxalyl-L--diaminopropionic acid
CHLI: Chlorophyll Index Chl a: Chlorophyll a DW: Dry Weight Eq.: Equation
Fj: Fluorescence Intensity at J step (at 2 ms) of Kautsky Fluorescence Induction Curves Fo: Minimal Fluorescence
Fm: Maximum Fluorescence Fv: Variable Fluorescence
Fv/ Fm: Maximum Quantum Efficiency of Photosystem II Fig.: Figure
FW: Fresh Weight
H2O: Water
Mo: Initial Slope of Kautsky Fluorescence Induction Curves NDVI: Normalized Difference Vegetation Index
PI: Performance Index PS I: Photosystem I PS II: Photosystem II
PRI: Photochemical Reflectance Index
QA: Primary Quinone Acceptor of PSII
R: Reflectance
RCs: Reaction Centers of PS RWC: Relative Water Content SW: Soil Weight
SWC: Soil Water Content TW: Turgid Weight
Vj: Relative Variable Fluorescence at the step J (at 2 ms) of Kautsky Fluorescence Induction
Curves
ix INDEX AGRADECIMENTOS ... iii RESUMO ... iv ABSTRACT ... vii ABBREVIATIONS ... viii 1. INTRODUCTION ... 1 1.1. Genus Lathyrus ... 1 1.1.1. Lathyus spp. ... 1 1.2. Economic Potential ... 3
1.3. Accumulation of -N-Oxalyl-L--diaminopropionic acid ... 4
1.4. Water Deficit Stress ... 5
1.4.1. General Adaptation Strategies ... 6
2. OBJECTIVES ... 7
3. MATERIALS AND METHODS ... 8
3.1. Plant material and growth conditions ... 8
3.2. Experimental design and water treatments imposed ... 8
3.3. Relative Water Content ... 9
3.4. Evapotranspiration and Soil Water Content ... 9
3.5. Chlorophyll Fluorescence ... 10
3.6. Reflectance... 10
3.7. Chlorophyll Determination ... 11
3.8. Statistical analysis ... 11
4. RESULTS AND DISCUSSION ... 12
4.1. Germination and Growth ... 12
4.2. Physiological Characterization of ten Lathyrus genotypes in well watered conditions ... 14
4.2.1. Relative Water Content (RWC) ... 14
4.2.2. Evapotranspiration Rate ... 15
4.2.3. Chlorophyll Fluorescence ... 16
4.2.4. Reflectance ... 19
4.2.5. Chlorophyll content (SPAD) ... 20
4.3. Physiological Characterization of ten Lathyrus genotypes in water deficit stress conditions 21 5. CONCLUSIONS AND FUTURE PERSPECTIVES ... 38
6. REFERENCES ... 39
1
1. INTRODUCTION
1.1. Genus Lathyrus
Lathyrus genus belongs to the family Leguminosae (Fabaceae), subfamily Pappilionoideae and tribe Viciae and comprises a large number of nearly 187 species and subspecies (Campbell, 1997 and references therein). The first archaeological evidences of Lathyrus appear in the Neolithic Period in the Balkans and in Near East countries of Bulgaria, Cyprus, Iraq, Iran and Turkey (Erskine et al., 1994). However, there are some reports suggesting that it was not grown intentionally and its domestication only occurred by the Bronze Age, until then it was just a tolerated weed in other crops such as lentil crops (Lens culinaris) (Erskine et al., 1994). Lathyrus sativus was also traced in Italy and South of France, and an isolated sample was reported from early Bronze Age in Portugal (Campbell, 1997).
Several Lathyrus species, and specifically L. sativus and L. cicera, have great potential as food crops in areas of major population growth where water shortage is a main concern (Vaz Patto et al., 2006). Lathyrus species are extensively cultivated in Northern Africa countries like Ethiopia, Egypt, Morocco, Algeria and Libya; in some Middle East countries as Iraq, Iran, Afghanistan, Syria and Lebanon; and in less extent in some South European countries like Spain or France (Campbell et al., 1994 and Tadesse and Bekele, 2003).
1.1.1. Lathyus spp.
Lathyrus aphaca is an annual plant with simple leafs and tendrils, its inflorescences are pedunculated, without bracts and reduced to a single yellow small flower. Pods are elliptical, oblong or slightly sickle and have about 4 to 7 seeds. These can be spherical or sub-spherical, smooth and generally brownish (www.floraiberica.es).
Lathyrus cicera or chickling pea, as called, is an annual or perennial shrub and can be glabrous or pubescent, has petiolated leaves with one pair of opposite leaflets, the bottom ones have simple tendrils whereas middle and higher leafs have branched tendrils (usually 3 branches). It has pedunculated inflorescences with bracts, reduced to only one flower. Its seed case holds about 3 to 6 seeds and has a large keel on each side of the ventral and dorsal margins. Seeds are spherical or prismatic, with smooth and brownish faces that can be flat or slightly concave (www.floraiberica.es).
2
Lathyrus clymenum is an annual or biennial climber without leaflets at the bottom leaves but higher leaves topped by long branched tendrils have 4 to 8 alternate leaflets. Its inflorescences are pedunculated, without bracts and have 1 to 2 flowers. Pods have an elliptical slightly obovate shape with a ventral peak and 2 ventral keels that holds 4 to 7 rounded, smooth and brownish seeds (www.floraiberica.es).
Lathyrus latifolius are robust, perennial and climbing shrubs characterized by densely packed stems and ability of vegetative propagation due to its ligneous rhizome which every year develops stems (Hossaert-McKey and Jarry, 1992 and www.floraiberica.es). Leaves are petiolated and have a pair of opposite leaflets; inflorescences are pedunculated with bracts and have 3 to 12 flowers. Fruits are oblong shaped with 3 keels on the ventral margin and can have 13 to 22 seeds, which are brown, sub-spherical with flat and rough surfaces (www.floraiberica.es).
Lathyrus ochrus are climbing, annual or biennial shrubs with simple bottom leaves and tendrils whereas upper leaves have 2 to 3 alternate leaflets and are topped with branched tendrils. Their inflorescences are pedunculated with only one flower and no bracts; fruits are elliptical with 2 ventral wings and have 6 to 8 spherical, smooth and brownish seeds (www.floraiberica.es).
Lathyrus odoratus commonly called “sweet pea” is an annual or biennial climbing shrub with a rich yellowish indumentum, petiolated leaves composed by 2 opposite leaflets and branched tendrils which supports and helps the plant reaching a height of 2 m with proper support (Ochatt et al., 2010 and www.floraiberica.es). It has pedunculated inflorescences with 2 flowers, oblong-elliptical fruits with a small keel on each side of the suture and another one on the dorsal area. Fruits also have a yellowish and glossy indumentum; they hold 5 to 8 rough and sub-spherical seeds (www.floraiberica.es).
Lathyrus sativus also known as grass pea is an annual and climbing shrub with a very penetrating and hardy root system suitable for a wide range of soil types, including very poor soils and heavy clays (Campbell et al., 1994 and www.floraiberica.es). It has petiolated leaves with a pair of opposite leaflets and upper leaves ending in branched tendrils; its inflorescences are pedunculated with bracts and have only one flower. Pods are elliptical with 2 wings in ventral margin, one on each side of the suture. Each pod holds 1 to 3 seeds, which can be white or brownish and have a smooth sub-rectangular or trapezoid shape (www.floraiberica.es). Its ancestor is not known, although there are several Mediterranean species sharing morphological resemblance particularly L. cicera L., L. marmoratus Boiss
3
and L. blepharocarpus Boiss, and its current distribution pattern by Mediterranean regions, Ethiopia and Indian Sub-continent resembling that of lentil (Erskine et al., 1994).
Lathyrus sylvestris is a climbing, perennial shrub found in high-altitude sites in Europe with an extended rhizome that allows vegetative propagation (Hossaert-McKey and Jarry, 1992 and www.floraiberica.es). It has petiolated leaves with one pair of opposite leaflets ending in a branched tendril. Their inflorescences are pedunculated with bracts and hold 2 to 8 flowers. The resulting fruit has an elliptical shape and 3 keels in ventral margin and contains about 10 to 15 seeds, these are sub-spherical, rough and brownish (www.floraiberica.es).
L. sylvestris and L. latifolius share a lot of common characteristics, both are known as “everlasting peas”, invest greatly in growth throughout the year, even when chances of survival are low, and they have a narrow flowering period in summer due to a requirement of a long-day photoperiod for the initiation of floral meristems (Hossaert-McKey and Jarry, 1992).
1.2. Economic Potential
Although Lathyrus is a genus with a large number of species only few have economic potential. Those are mainly food crops for human and livestock feed or forage crops and, less commonly, ornamental plants (Campbell 1997 and Fernández-Aparicio and Rubiales, 2010). L. sativus is grown for green feed, grain and forage for both animal and human consumption, L. cicera is grown primarily as stock feed both as fodder and grain (Hanbury et al., 1999). Other species like L. ochrus, L. latifolius and L. sylvestris are used as forage crops (Campbell 1997). L. odoratus is widely used as ornament in gardens for its flowers’ bright colors and its sweet fragrance having a vast number of cultivars commercially available (Ochatt et al., 2010).
L. sativus has a great agricultural and economic value due to its high resilience to drought stress conditions as it has been grown successfully in areas with a low mean annual precipitation (Campbell et al., 1994). In these areas, where soil quality is poor and extreme environmental conditions prevail, its hardiness and ability to fix nitrogen make this species very suitable for growth (Campbell et al., 1994 and Campbell 1997). L. sativus is also known for withstand floods, like in South Asia where is common to broadcast its seeds in rice crops some weeks before harvesting, so by the time water is allowed to drain for the harvesting of
4
rice L. sativus can germinate and grow with residual water left in field (Campbell 1997). As it can be used as fodder or green manure plants are normally pulled while steel green but only after pods have filled, in this way a biomass maximum nutritional value is achieved allowing good seed yields (Campbell 1997).
Human consumption of L. sativus grains differs greatly between regions, as in India where grains can be boiled or processed in a dal mill to prepare the soup-like dish dal, or can be ground resulting in a flour that is sold as besan, used to make various side dishes (Campbell 1997). In Ethiopia grass pea can be consumed as kitta, an unleavened bread, or its flour used in preparation of a sauce called wott, but it’s widely eaten as boiled grains (Campbell 1997).
1.3. Accumulation of -N-Oxalyl-L--diaminopropionic acid
Approximately 21 species of genus Lathyrus produces the toxic non-protein amino acid
-N-oxalyl-L--diaminopropionic acid ( - ODAP) (Rao et al., 1963 and Addis and Narayan, 1994). The toxicity displayed by this compound puts it as the causative agent of the clinical syndrome neurolathyrism, an irreversible neurodegeneration of motor neurons with symptoms from inability to walk to complete paralysis of the lower limbs (Rao et al., 1963; Roy et al., 1963 and Addis and Narayan, 1994). This syndrome is attributed to a prolonged over-consumption of seeds from L. sativus during drought triggered famines especially in Asia and Africa, where for several months these seeds are the staple food in an unbalanced diet (Getahun et al., 1999 and Lambein et al., 2007).
The rate of biosynthesis and accumulation varies during ontogeny of the plant and between species, but environmental conditions also seem to have a role (Addis and Narayan, 1994). Several studies have been made as an effort to clarify the factors involved in the biosynthesis and accumulation of -ODAP, together with many attempts for improvement of Lathyrus lines with reduced -ODAP concentrations (Campbell et al., 1994; Gengsheng et al., 2001 and Yang et al., 2004). For instance, in India some L. sativus lines that had been selected for reduced ODAP concentrations in a hybridization and selection program in Canada, designed to develop adapted high-yielding material with reduced concentrations of the neurotoxin, were tested (Campbell et al., 1994).
5 1.4. Water Deficit Stress
In natural environments water deficit stress arises usually by the lack of rainfall (drought stress), but is also a component of other stresses like salt or osmotic (Hopkins, 1995b). During periods of drought plants suffer from water deficits that lowers w leading to inhibition
of plant growth and photosynthesis, as we can see by the relationship described in Fig.
1(Taiz and Zeiger, 2006a). By lowering the soil w and the ability of plants to take up water,
drought quickly reduces the rate of cell expansion in growing tissues leading to a decreased photosynthetic leaf area which in turn slows the flow of assimilates to the growing tissues (Munns, 2011). Not only there is a decrease in photosynthetic leaf area, as photosynthesis itself can be affected by water stress by the closure of stomata that cuts off access of the chloroplast to the atmospheric supply of carbon dioxide (CO2) and by the direct effects of low
cellular w on the structural integrity of the photosynthetic machinery (Hopkins, 1995b). At a
cellular level changes resulting from water deficit are related with changes in the cell processes such as cell cycle and division, but also with changes in cell wall architecture and in the endomembrane system (Taiz and Zeiger, 2006b).
The resulting damage is related to detrimental effects of desiccation on protoplasm, such as the increase of non-compatible solute concentration as its volume decreases, and with the disruption of the structure in the membrane bilayer introducing water-filled channels lined with polar phospholipid head groups (Hopkins, 1995b). As with membrane integrity protein structure is also affected leading to a loss of membrane selectivity and loss of function of membrane based enzymes (Hopkins, 1995b).
Fig. 1– Relationship between some
physiological processes and water potential of plants under various growing conditions. The intensity of the bar color corresponds to the magnitude of the process. Adapted from Taiz and Zeiger (2006a).
6 1.4.1. General Adaptation Strategies
In drought-prone environments several strategies concerning physiological traits such as closed stomata, dense trichome layer, high leaf angles or decreased canopy leaf area are adopted by plants to minimize water loss (Chaves et al., 2003; references therein). Although these effects are generalized, plants from different environments have different adaptations to drought. For example in humid regions plants progressively reduce their transpiration closing its stomata, otherwise in dry regions they have deep roots or water-storing tissues avoiding reducing transpiration or like evergreen sclerophyll trees and shrubs in regions with periodic summer drought which can limit their water consumption in 10 to 20% (Larcher, 1983). Decreasing canopy leaf area through reduced growth and shedding of older leaves that contribute to water saving can be viewed as a recycling program within the plant, allowing the reallocation of nutrients stored in older leaves to the stem or younger leaves (Chaves et al., 2003). These adaptations can be divided into two components: drought avoidance, which is the capacity of tissues to resist the formation of damaging water deficits, and drought tolerance which is the ability to function with high water deficits (Bannister, 1986 and Taiz and Zeiger, 2006b). Dehydration avoidance is common to both annuals and perennials and is associated with a variety of adaptive traits involving minimizing water loss and maximizing water uptake by avoiding tissue dehydration while maintaining tissue w as
high as possible, or by tolerating low tissue w (Chaves et al., 2003). In order to maintain
cell expansion, and despite decreasing cell w, there is an accumulation of solutes in the cell
that is able to maintain a positive p needed for the continuation of this process (Taiz and
Zeiger, 2006a). These are just some examples of cellular and physiological plasticity that plants show upon situations of water stress deficit.
7
2. OBJECTIVES
The leading aim of the present work was to contribute to the preliminary characterization of the agronomic potential of ten genotypes of Lathyrus. With this purpose, two main objectives were set:
1) To understand the relative photosynthetic performance at well watered conditions; 2) To understand the relative photosynthetic performance at water deficit stress.
A secondary aim of this work was methodological: to test the capacity of a set of non-invasive physiological and photochemical techniques to detect the effects of water stress and discriminate between genotypes.
8
3. MATERIALS AND METHODS
3.1. Plant material and growth conditions
Seeds of Lathyrus aphaca (L375), Lathyrus cicera (L94), Lathyrus clymenum (L388), Lathyrus latifolius (L563), Lathyrus ochrus (L313), Lathyrus odoratus (L17-1), Lathyrus sativus (L174-A) and Lathyrus sylvestris (L531). All obtained from Centro de Recursos Fitogenéticos (CRF/INIA), Madrid, Spain through Professor Diego Rubiales (IAS-CSIC, Córdoba, Spain). Lathyrus sativus (LS 87124), low Beta-ODAP (0.02%) variety developed by Dr Clayton Campbell from Agriculture and Agri-Food Canada, Morden ResearchCenter in Canada. Lathyrus sativus (Raipur), high beta-ODAP (0.54%) local variety from the Raipur region in India. Both were obtained through Professor Lambein (IPBO, Ghent, Belgium).
Fifteen to twenty seeds per genotype were surface sterilized by immersion in a solution of 50% (v/v) commercial bleach and 50% (v/v) distilled water 3 for 5 minutes, washed three times with sterile distilled water and disinfected again with70% (v/v) ethanol for 2 minutes. After an additional washing step with sterile distilled water, the seeds were left immersed in water for 30 minutes and plated in Petri dishes with sterile absorbent with constant humidity. Seeds were then placed in dark at 4 oC for 48 h (vernalization period), after which they were transferred to 25 oC until the radicle emergence. Ten seedlings for each genotype were planted in 2 L pots with a mixture of a rich soil (Substrato Especial Relvados e PradosTM, Santos & Santos Substratos Biológicos, Portugal, detailed composition in Annex A), sand and vermiculite (1 mm) in a proportion of 1:1:2 (v/v/v). Plants were grown for 3 ½ months in a greenhouse and watered whenever necessary. Environmental conditions were registered with a HOBO® Weather Station (Onset Computer Corporation, USA). To avoid local microclimate effects, pots were randomly placed and moved in the greenhouse bench throughout the experience.
3.2. Experimental design and water treatments imposed
The characterization of the ten Lathyrus genotypes were performed in two different conditions; one in well watered conditions in a pre water deficit stress period and the other in water deficit conditions. The characterization in well watered conditions was assessed with measurements of relative water content (RWC), evapotranspiration rates, leaf chlorophyll determination, chlorophyll fluorescence and reflectance indices. For such ten plants for each genotype were used, all of them with 3 months old, although for L. aphaca leaf chlorophyll
9
determination and chlorophyll fluorescence measurements were not possible due to the small leaf size typical of this genotype.
Water deficit stress was induced by suppression of watering imposed to 5 potted plants randomly selected out of 10 for each genotype. The 5 remaining potted plants were control plants and continued to be watered as before. The last day of watering, when the water stress period began, is referred as day 0 (zero). The days preceding the beginning of the imposition of water stress are referred with a minus (-) signal, and on the other hand the days concerning the period of water stress are referred as “positive” days (ex. Day 14).
3.3. Relative Water Content
Relative water content (RWC) was used as a measure of plant water status and was calculated according to Catsky (1960) (Eq. 1). Fresh weight (FW) was determined immediately after sample collection, turgid weight (TW) was determined after saturating samples with milliQ water for 4h and dry weight (DW) measured after oven drying samples at 80 oC for 48 h.
[ ] (Eq. 1)
3.4. Evapotranspiration and Soil Water Content
Potted plants in well watered conditions were watered and allowed water to drain for 6 h, after which were weighed and then again at 8 h, 12 h, 24 h and 48 h. Weighing was done with an electronic balance KERN (KERN & Sohn GmbH, Germany). The rate of evapotranspiration was determined as the slope of the weight loss (g) plotted against time (h).
For soil water content (SWC) assessment during the water deficit stress period pot weight 6h after irrigation, as stated above, was considered the saturated weight (SW) and fresh weight (FW) was assessed with periodic weighing of stressed pots (Eq. 2). SWC was calculated as follows:
10 3.5. Chlorophyll Fluorescence
Fluorescence measurements were performed with a Handy Plant Efficiency Analyzer (PEA) – Chlorophyll Fluorimeter (Hansatech Instruments, England) by placing samples on leaf clips and allowing 5 min for dark adaptation, after which samples were exposed to a saturating light pulse with the duration of 1 s. The kinetics of the rapid fluorescence rise was recorded and the maximum quantum efficiency of photosystem II (Fv/Fm) (Eq. 3), performance index (PI) (Eq. 4) and complementary area (CAREA) of the chlorophyll
fluorescence induction curve were automatically calculated according to Strasser et al. (2000 and 2007).
(Eq. 3)
(Eq. 4)
Where Fm is the maximum fluorescence obtained when all the PSII reaction centers are open and Fo is the basal fluorescence, obtained when all PSII reaction centers are open. Vj is the relative variable fluorescence at the J step (at 2 ms) of the Kautsky induction curve, calculated as
,
where Fj is the fluorescence intensity also at the J step. Mo is the initial slope of fluorescence kinetics. Five measurements per potted plant per genotype were performed.3.6. Reflectance
Reflectance measurements in vivo were performed with 3 readings per potted plant for each genotype and were always performed during the period of highest sun exposure between 10 a.m. and 3 p.m. Reflectance was measured with UniSpec-SC (Single Channel) Spectral Analysis System (PP Systems, USA) by placing leaf sample in the standard leaf clip connected with a bifurcated fiber optic to the equipment. Water reflectance index (WRI) (Eq.
5), chlorophyll index (CHLI) (Eq. 6), photochemical reflectance index (PRI) (Eq. 7) and
normalized difference vegetation index (NDVI) (Eq. 8) were calculated according to Peñuelas et al. (1994, 1997 and 2004) and Gamon et al. (1997).
11 WI = (Eq. 5) CHL = (Eq. 6) PRI = (Eq. 7) NDVI = (Eq. 8) 3.7. Chlorophyll Determination
Relative Chlorophyll amount was determined with a Chlorophyll Meter SPAD-502 Plus (Spectrum Technologies, Inc., USA) with a measurement area of 2mm x 3mm where leaf samples were placed. Values were calculated based on the amount of light transmitted by the leaf in two specific wavelength regions and showed as “SPAD” values (with a precision of ± 1 SPAD unit, at room temperature and values between 0 – 50.0). A maximum of 15 readings per potted plant per genotype were assessed; with the exception of L. aphaca which at the time did not had leaves large enough to support the dimensions of the used device.
3.8. Statistical analysis
Assessment of data normality distribution was performed using the Kolmogorov-Smirnov Normality test with a Lilliefors Significance Correction. Since our data did not followed a normal distribution, genotype and measuring day effects were analyzed using Kruskal-Walkis non-parametric test. Multiple comparisons between factors were performed with Duncan’s post hoc test in case of homogeneous subsets and for cases that this was not verified Dunnett’s T3 post hoc tests were applied. Homogeneity of variances was assessed with Levene’s statistics. In all cases P 0.05 was considered significant. Statistical analysis was performed with IBM® SPSS® Statistics Version 19 (IBM Corporation, 2010). Correlation analysis was performed with GraphPad Prism version 5.03 for Windows (GraphPad Software, USA). Data from HOBO® Weather Station was analyzed with BoxCar® Pro version 4.3.1.1. (Onset Computer Corporation, 2002).
12
4. RESULTS AND DISCUSSION
4.1. Germination and Growth
Due to the variation in the percentage of germination between genotypes three germination cycles were necessary to achieve the number of seeds needed for the water stress experiments. With the highest percentage of germination, reaching 100%, stands out the genotype L. sativus (Raipur), although L. odoratus, L. ochrus, L. cicera and L. latifolius had percentages ranging from 70 to 100%. L. aphaca, L. clymenum and L. sativus (L 174A) showed values between 30 and 70% and the genotypes with the lowest germination success were L. sylvestris (13%) and the low ODAP variety from Canada of L. sativus (LS 87124) (21%). Due to this low seed viability the number of plants planned for each genotype was not achieved in these two latter cases, and we had to carry on with only two plants for L. sylvestris and eight for L. sativus (LS 87124) genotype. For the remaining genotypes, each one was represented with 10 plants.
The growth period in the greenhouse had the duration of 3 ½ months from early December to mid March, allowing for the proper development of each plant in well watered conditions, with plants being watered whenever necessary according to their stage of development. Weather conditions in the greenhouse were subjected to external environment, so for the growth period mean temperature was 15.1ºC (± 2.3) and relative humidity around 50.3% (± 5.9) (see Annex B). By the end of the growth period the phenological stage of the genotypes was diverse. The phenological stages could be divided in two major classes, with or without flowers. In the first group were L. cicera and the three genotypes of L. sativus, wherein the variety from Raipur (Raipur) and L. cicera had already some pods, and in the second were L. aphaca, L. clymenum, L. latifolius, L. ochrus, L. odoratus and L. sylvestris. Regardless of these two major phenological classes and the implications concerning different water requirements for different developmental stages, measurements and comparisons between genotypes were made throughout this work, though always having it into account. Morphological differences between genotypes were also a concern due to the fact that they were quite evident as showed by Fig. 2. Although this was not a limitation for the majority of genotypes it was for L. aphaca, due to its small sized leaves that were not compatible with measurement devices of fluorescence and chlorophyll (SPAD).
13
Fig. 2 – Photos of all ten genotypes at well watered conditions two days before de beginning of
stress period. (A) L. clymenum, (B) L. sativus (LS 87124), (C) L. sativus (L174-A), (D) L. ochrus, (E) L. latifolius, (F) L. sativus (Raipur), (G) L. aphaca, (H) L. odoratus, (I) L. cicera, (J) L. sylvestris.
A B C
D E F G
14
4.2. Physiological Characterization of ten Lathyrus genotypes in well watered conditions
Lathyrus sp. genotypes are widely known for their proprieties as food or legume forage crops, namely L. sativus (L174-A) and L. cicera, and other species, like L. odoratus, are also used with ornamental purposes (Vaz Patto et al., 2006 and Ochatt et al., 2010). Lathyrus drought resilience is well known, especially in dry land areas where it is widely cultivate, even though there is a lack of information concerning the physiology beyond this resistance. For a comprehensive characterization of the ten genotypes used in this work several physiological parameters were measured and studied in 3 months old plants in well watered conditions.
4.2.1. Relative Water Content (RWC)
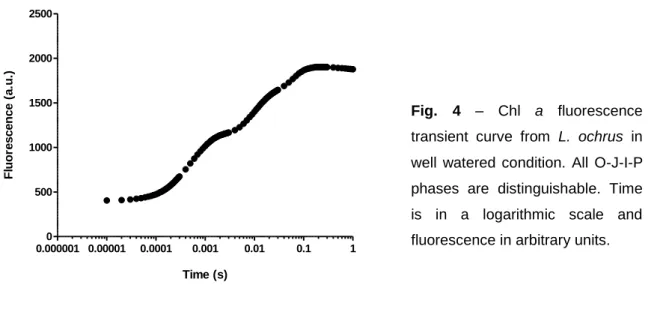
RWC assessment shows that all genotypes were well hydrated with values ranging from 85.8% (± 6.0) to a maximum of 94.0% (± 5.1) where differences among genotypes were minimal, for most of the cases (Fig. 3). Even though differences between genotypes can occur, like the ones observed for L. cicera (88.4% ± 2.7) and L. odoratus (92.5% ± 4.2). At this point of well watered conditions (only two days before the beginning of stress period) the different genotypes were not in the same developmental stage, and in this case L. cicera had flowers and some pods, in contrast with L. odoratus, which could contribute to the differences found. L. a phac a L. c icer a L. c lyme num L. la tifol ius L. o chru s L. o dora tus L. s ativ us L. s ativ us (C anad a) L. s ativ us (I ndia ) L. s ylve stri s 0 20 40 60 80 100 ab a ab ab ab b ab ab ab ab R W C ( % )
Fig. 3 – Relative water content (%) for each
genotype in well watered conditions. Different letters over bars indicate significant differences among genotypes (P< 0.05, Dunnett’s T3 multiple comparison test). Values are mean ± SD
15 4.2.2. Evapotranspiration Rate
Transpiration, the process by which water in plants is lost as water vapor, is one of the major factors concerning the water use efficiency of crop fields. Both the transpiration rate from plants and the evaporation of water from the surface of the soil, which together are known as evapotranspiration, contribute to water use efficiency (Kramer and Boyer, 1995 and Hopkins, 1995c).
Evapotranspiration rate represents the mass of water (g) lost from potted plants per hour in all 10 genotypes (Table 1). As we can see from Table 1 there are significant differences among genotypes. L. clymenum shows the highest value (1.327 g H2O.h-1 ± 0.356) in
contrast with L. aphaca (0.580 g H2O.h-1 ± 0.092) and L. latifolius (0.556 g H2O.h-1 ± 0.091),
suggesting a greater evapotranspiration rate than L. clymenum or even L. cicera since for the same period of time they lose less than half the water. All three L. sativus genotypes had high values of evapotranspiration rate, which suggests that they are less adapted to aridity.
Table 1 – Evapotranspiration rate (g H2O.h
-1
) and relative water content (%) for each Lathyrus sp. genotype measured at well watered conditions. For both cases, different letters indicate significant differences among genotypes (P<0.05, Dunnett’s T3 multiple comparison test). Values are mean ± SD.
Genotype Evapotranspiration Rate ( g H2O.h-1) RWC (%)
L. aphaca 0.580 ±0.092 a 88.3 ±6.5 ab L. cicera 1.305 ±0.527 bc 88.4 ±2.7 a L. clymenum 1.327 ±0.356 c 90.8 ±5.4 ab L. latifolius 0.556 ±0.091 a 92.2 ±4.4 ab L. ochrus 0.941 ±0.482 abc 94.1 ±5.1 ab L. odoratus 0.709 ±0.259 ab 92.5 ±4.2 b
L. sativus (L174-A) 1.187 ±0.463 abc 85.8 ±6.0 ab
L. sativus (LS87124) 1.052 ±0.373 abc 91.4 ±5.0 ab
L. sativus (Raipur) 0.636 ±0.165 ab 92.3 ±4.5 ab
16 4.2.3. Chlorophyll Fluorescence
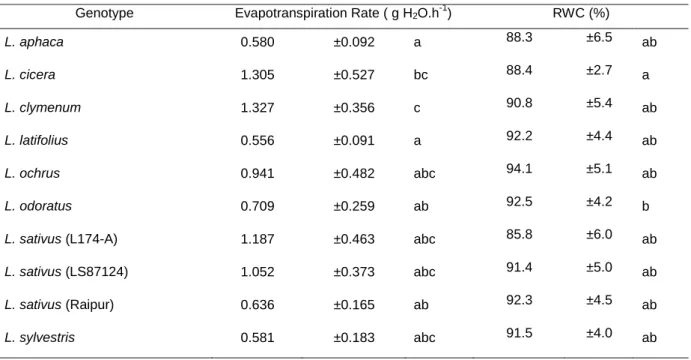
When a dark adapted leaf is exposed to light, large and rapid (in less than a second) changes in chlorophyll fluorescence occur. This process is known as Chl a fluorescence transient or the Kautsky transient (Strasser et al., 2007). On exposure to a strong light, fluorescence rises from the minimal level (Fo), where the primary quinone acceptor of PSII (QA) is maximally oxidized and the reaction centers (RCs) are all open, to a maximal
fluorescence level (Fm) where QA is reduced and RCs are all closed (Baker and Rosenqvist,
2004 and Strasser et al., 2007). This rise is polyphasic, with the steps O (= Fo) at 50 s, J at approximately 2 ms, I at around 30 ms and P (= Fm) at about 300 ms, although the time required for the achievement of these steps is dependent upon the light intensity (Strasser, 1997 and Strasser et al., 2000). Fig. 4shows the Chl a fluorescence transient measured for L. ochrus genotype in well watered control conditions. The steps are related with different phases in the PSII reactions, so the O to J rise is attributed to partial closure of RCs, from J to I represents closure of the remaining RCs and the rise of I to P is the removal of plastoquinone quenching (Strasser, 1997).
Measurements of Chl a fluorescence transient provides a set of parameters very useful to assess the physiological status of leaves and plant photosynthetic performance in a wide range of situations (Baker and Rosenqvist, 2004 and Strasser et al., 2000). Such parameters includes the maximum quantum yield of PSII (expressed as Fv/Fm and calculated as (Fm-Fo)/Fm)), the performance index (PI) and the complementary area (CAREA) between the
fluorescence transient curve and the Fm (Strasser et al., 2000). Dark adapted values of Fv/Fm reflect the potential maximum quantum efficiency of PSII and are used as a sensitive indicator of plant photosynthetic performance (Maxwell and Johnson, 2000). Performance index (PI) is an integrative parameter that comprises the density of fully active reaction centers (RCs), the efficiency of electron movement by trapped exciton into electron transport chain beyond the QA and the probability that an absorbed photon will be trapped by RCs
(Zivcák et al., 2008). CAREA reflects the pool size of the electron acceptors QA on the
17 0.000001 0.00001 0.0001 0.001 0.01 0.1 1 0 500 1000 1500 2000 2500 Time (s) F lu o re s c e n c e ( a .u .)
To assess the behavior and variation of Lathyrus genotypes to well watered conditions five fluorescence measurements were performed in each plant in five separate days. The results show significant variation of some fluorescence parameters along these days, for some genotypes (see Annexes C, D and E). These differences vary with genotype and the studied parameter, with only one exception for the low -ODAP L. sativus (LS87124) genotype which shows no differences at all. L. latifolius sustains steady values of Fv/Fm, while L. ochrus has no differences either in PI values or in the CAREA values, which in this last
case so does L. sylvestris. Although these fluctuations with time in the fluorescence parameters of well irrigated plants were statistically significant, no observable trend was found. Furthermore, no relation with environmental conditions was evident and thereby we could not find any explanation for this behavior. Despite these results we used the pool values to look for differences between genotypes.
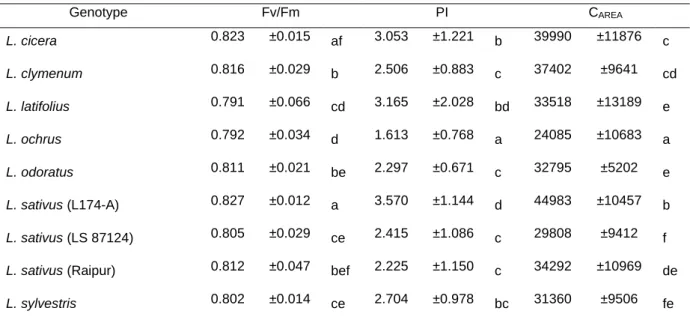
The statistical analyses performed to the fluorescence parameters studied shows significant differences between genotypes although these differences vary with each parameter (Table 2). Fv/Fm is a very steady parameter with values described by Björkman and Demmig (1987) in 37 C3 species of around 0.832. Our results show a tendency similar
to the described for several species, including legumes Glycine max and Phaseolus vulgaris (Björkman and Demmig, 1987). Although this great stability our results show some significant differences, where L. sativus (L174-A) (0.827 ± 0.012) and L. cicera (0.823 ± 0.015) stands out with the highest values, even though L. cicera cannot be distinguished from the variety from Raipur of L. sativus (0.812 ± 0.047). On the other hand with the lowest Fv/Fm value there is L. ochrus (0.792 ± 0.034) and L. latifolius (0.791 ± 0.066), although this last genotype also cannot be set apart from L. sativus (LS87124) (0.805 ± 0.029) and L.
Fig. 4 – Chl a fluorescence
transient curve from L. ochrus in well watered condition. All O-J-I-P phases are distinguishable. Time is in a logarithmic scale and fluorescence in arbitrary units.
18
sylvestris (0.802 ± 0.014) probably, due to its high standard deviation that does not allow them to be statistically different.
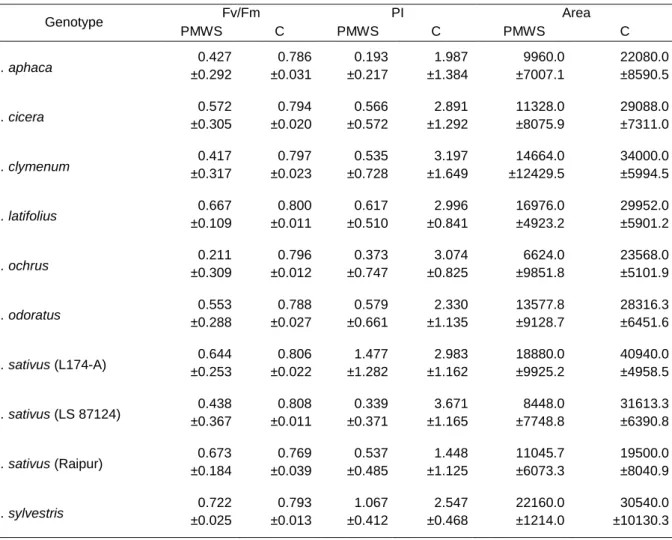
Table 2 – Values of Fv/Fm, PI and CAREA for each genotype measured in well watered conditions.
Different letters aside values shows significant differences among genotypes (P<0.05, Dunnett’s T3 multiple comparison test). Values are mean ± SD.
Genotype Fv/Fm PI CAREA L. cicera 0.823 ±0.015 af 3.053 ±1.221 b 39990 ±11876 c L. clymenum 0.816 ±0.029 b 2.506 ±0.883 c 37402 ±9641 cd L. latifolius 0.791 ±0.066 cd 3.165 ±2.028 bd 33518 ±13189 e L. ochrus 0.792 ±0.034 d 1.613 ±0.768 a 24085 ±10683 a L. odoratus 0.811 ±0.021 be 2.297 ±0.671 c 32795 ±5202 e L. sativus (L174-A) 0.827 ±0.012 a 3.570 ±1.144 d 44983 ±10457 b L. sativus (LS 87124) 0.805 ±0.029 ce 2.415 ±1.086 c 29808 ±9412 f L. sativus (Raipur) 0.812 ±0.047 bef 2.225 ±1.150 c 34292 ±10969 de L. sylvestris 0.802 ±0.014 ce 2.704 ±0.978 bc 31360 ±9506 fe
More distinctive differences were found between genotypes for the PI as shown by table 2. There is a distinct set with the highest values for PI which comprises L. sativus (L174-A) (3.570 ± 1.144), L. cicera (3.053 ± 1.221) and L. latifolius (3.165 ± 2.028) although also L. sylvestris (2.704 ± 0.978) could belong to this group but due to its convergence of values with several of the other genotypes it was not included. L. ochrus (1.613 ± 0.768) stands out has the one with the lowest PI values. We can group the genotypes in three classes: the one with the highest values includes L. sativus (L174-A) (3.570 ± 1.144), L. cicera (3.053 ± 1.221) and L. latifolius (3.165 ± 2.028), a group with intermediate values is formed by L. clymenum (2.506 ± 0.883), L. odoratus (2.297 ± 0.671) and L. sylvestris (2.704 ± 0.978) and both L. sativus (LS 87124) (3.570 ± 1.144) and L. sativus (Raipur) (2.225 ± 1.150) and L. ochrus (1.613 ± 0.768) showed distinctively lower values. Values for CAREA follow the same
pattern of both Fv/Fm and PI when it comes to L. sativus (L174-A) (44983 ± 10457) and L. ochrus (24085 ± 10683), although in this case L. sativus (LS 87124) showed similar low values (29808 ± 9412). The other genotypes presented intermediate values.
19 4.2.4. Reflectance
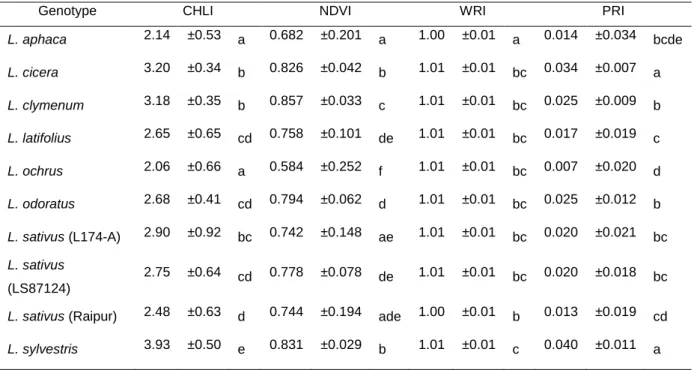
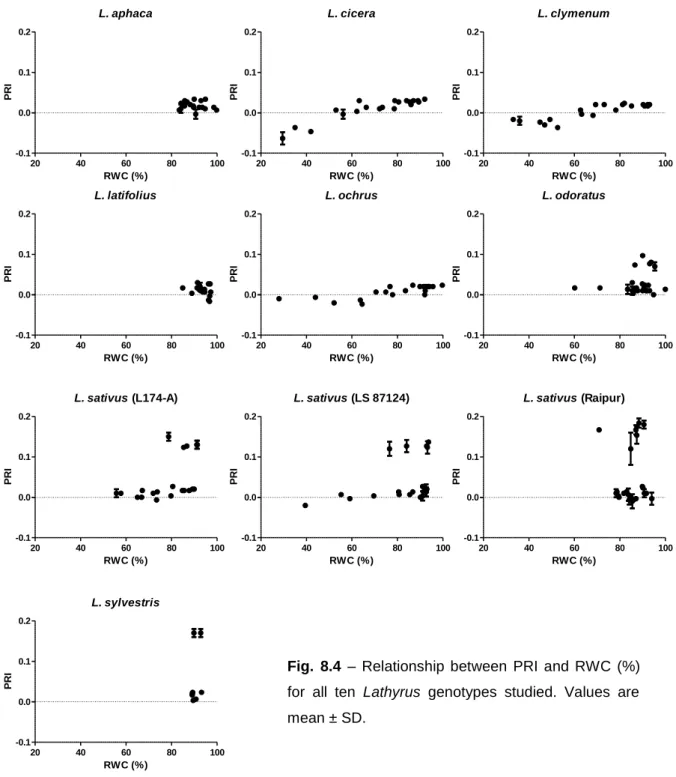
Assessing plant water status based on water absorbance of light at certain wavelengths in the near-infrared portion of the electromagnetic spectrum is a very simple non-destructive method. If a beam of light at the wavelength of the water absorption bands interacts with plant material the reflectance of this light is reduced to varying extent depending on tissue water content (Peñuelas et al., 1993). For the Water Reflection Index (WRI) wavelengths between 900 and 970 nm were chosen due to the strong relationship with RWC (Peñuelas et al., 1993). Normalized Difference Vegetation Index (NDVI) is a broadband index related to green biomass that is widely used for indirect estimates of photosynthetic capacity and net primary production (Peñuelas et al., 1994). The Photochemical Reflectance Index (PRI) calculated based in wavelengths of 531 nm which are related to several processes of dissipation of excess radiation, like the xanthophyll cycle, provides a way of estimating photosynthetic performance using spectral reflectance (Gamon et al., 1997 and Peñuelas et al., 2004). Leaf chlorophyll concentration is an indicator of plant N status (Daughtry et al., 2000). Chlorophyll Reflectance Index (CHLI) detects changes in leaf chlorophyll concentrations constituting a simple and non-destructive method for leaf chlorophyll assessment.
Reflectance was measured in four distinct days to assess any fluctuations of values along well watered conditions. The analysis of variance performed showed significant differences between measurement days for the four indices studied, although none of these indices revealed a consistent pattern of variation or any relationship with any other parameter in this study. Therefore, and just like what happened with fluorescence, we could not explain such variation and used the pool values to look for differences between genotypes.
For all the four indices calculated from reflectance measurements L. sylvestris stands out with the highest values, although for NDVI and PRI is accompanied by L. cicera (0.826 ± 0.042 and 0.034 ± 0.007, respectively) and for WRI by L. sativus (Raipur) (1.00 ± 0.01) (Table 3). On the other edge with the lowest values for CHLI and NDVI are L. ochrus (2.06 ± 0.66 and 0.584 ± 0.252, respectively) and L. aphaca (2.14 ± 0.53 and 0.682 ± 0.201, respectively), for PRI L. ochrus (0.007 ± 0.020) is accompanied by L. latifolius (0.017 ± 0.019) and for WRI only L. aphaca (1.00 ± 0.01) stands out with the lowest value. All other genotypes presented an intermediate behavior with some fluctuations not to relevant to mention. NDVI values obtained by Peñuelas et al. (1994) in sunflower were in the range of 0.75 and 0.77 for the correspondent day time as used in this work. Values of PRI for the same period ranged between 0.04 and 0.08.
20
Table 3 – Values of CHLI, NDVI, WRI and PRI for each genotype measured in well watered
conditions. Different letters aside values shows significant differences among genotypes (P<0.05, Dunnett’s T3 or Duncan’s multiple comparison test, when valid). Values are mean ± SD.
Genotype CHLI NDVI WRI PRI
L. aphaca 2.14 ±0.53 a 0.682 ±0.201 a 1.00 ±0.01 a 0.014 ±0.034 bcde L. cicera 3.20 ±0.34 b 0.826 ±0.042 b 1.01 ±0.01 bc 0.034 ±0.007 a L. clymenum 3.18 ±0.35 b 0.857 ±0.033 c 1.01 ±0.01 bc 0.025 ±0.009 b L. latifolius 2.65 ±0.65 cd 0.758 ±0.101 de 1.01 ±0.01 bc 0.017 ±0.019 c L. ochrus 2.06 ±0.66 a 0.584 ±0.252 f 1.01 ±0.01 bc 0.007 ±0.020 d L. odoratus 2.68 ±0.41 cd 0.794 ±0.062 d 1.01 ±0.01 bc 0.025 ±0.012 b L. sativus (L174-A) 2.90 ±0.92 bc 0.742 ±0.148 ae 1.01 ±0.01 bc 0.020 ±0.021 bc L. sativus (LS87124) 2.75 ±0.64 cd 0.778 ±0.078 de 1.01 ±0.01 bc 0.020 ±0.018 bc
L. sativus (Raipur) 2.48 ±0.63 d 0.744 ±0.194 ade 1.00 ±0.01 b 0.013 ±0.019 cd L. sylvestris 3.93 ±0.50 e 0.831 ±0.029 b 1.01 ±0.01 c 0.040 ±0.011 a
4.2.5. Chlorophyll content (SPAD)
A rapid chlorophyll measurement was performed with a Chlorophyll Meter SPAD-502 Plus (Spectrum Technologies, Inc., USA) and the results were assessed only in relation to variations between genotypes, so they are expressed as SPAD units. As showed by figure 5 three separated groups can be distinguished given the SPAD values of chlorophyll. The highest ranging 40 SPAD units are referred to genotypes L. cicera (39.2 SPAD ± 3.8), L. sativus (L174-A) (39.4 SPAD ± 5.2) and L. sylvestris (36.5 SPAD ± 7.7), an intermediate group statistically different from the other two with values fluctuating around 30 SPAD units is composed by L. clymenum (33.5 SPAD ± 4.1), L. latifolius (31.8 SPAD ± 5.0), L. odoratus (31.0 SPAD ± 4.7) and both L. sativus (LS87124) (33.8 SPAD ± 6.8) and L. sativus (Raipur) (32.0 SPAD ± 5.4) and with the lowest range is L. ochrus (23.8 SPAD ± 5.8) genotype. L. aphaca does not have any values for this parameter due to the nature of the measuring device that was to large for the small leaves characteristic of this genotype.
When these results are compared with the CHLI from reflectance the outcomes are quite similar. Although no values of SPAD for L. aphaca are presented and L. sativus in CHLI analysis does not stands out, which can be explained by the relatively high standard deviation, the comparison between these two parameters applies. Results from both
21
methods are consistent with regard to distinguished groups, not only validating each other but also settling differences between genotypes.
L. c icer a L. s ativ us L. s ylve stri s L. c lyme num L. la tifol ius L. o dora tus L. s ativ us (C anad a) L. s ativ us (I ndia ) L. o chru s 0 10 20 30 40 50 c a a a b b b b b C h lo ro p h y ll ( S P A D )
4.3. Physiological Characterization of ten Lathyrus genotypes in water deficit stress conditions
The water stress period began at day zero with the last watering. Several parameters were assessed throughout the experiment not only to allow a complete characterization of the several stages but also to gain knowledge of when each genotype reached the point of maximum water stress (PMWS) to allow a recovery period afterwards. For this purpose RWC and Fv/Fm, in a combined analysis, were the factors chosen. Initially the PMWS was allowed to reach RWC close to 30% resulting in some genotypes reaching severe stress levels. Although due to the absence of recovery afterwards the assessment of PMWS was performed with the assistance of Fv/Fm. Values equal or less than 0.75 were considered as the threshold for considering that plants had reached the PMWS. At this point stressed plants were rewatered to study the physiological recovery.
In this work the soil used was a mixture of a rich soil with vermiculite and sand (see section 2). Lathyrus plants are legumes which are known for their mycorrhization process and the ability to fix nitrogen (N) and so growth is potentiated in a poor soil. Utilization of inert materials, like sand and vermiculite allowed not only the impoverishment of the soil but also causing it to be less dense or compact, so appropriate conditions would form for processes like mycorrhization and N fixation. Furthermore, vermiculite has a high water
Fig. 5 – Relative chlorophyll content (in SPAD
units) for each genotype in well watered conditions. Different letter over bars indicate significant differences among genotypes (P< 0.05, Dunnett’s T3 multiple comparison test). Values are mean ± SD.
22
retention capacity and by itself it would retain too much water and stress would be installed at a very slow pace, so in order to balance the water retention capacity of the soil sand was added.
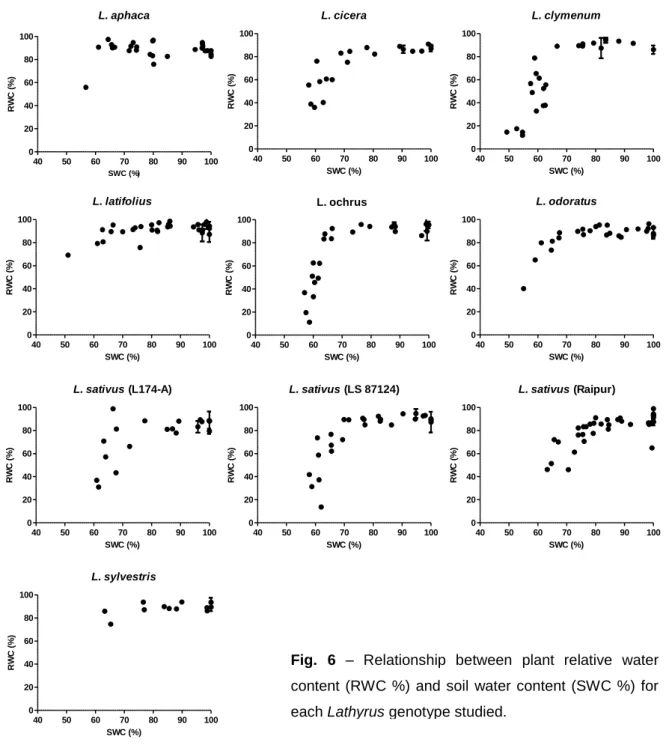
Fig. 6 shows RWC in function of SWC for each genotype, where it is visible a gradual loss of soil water and a gradual response of plant RWC. At values higher that 80% of SWC all genotypes were able to maintain high RWC, when SWC began to drop below the 80% threshold so did RWC, although for L. aphaca, L. latifolius and L. sylvestris high values of RWC were still maintained. Values lower than 70% of SWC resulted in a pronounced decreased of RWC; this behavior is specially marked in genotypes L. cicera, L. clymenum, L. ochrus and L. sativus that had a shorter duration of their stress periods. On the contrary we cannot see this relationship in L. latifolius or in L. sylvestris where with a SWC of 60% the RWC maintained its values between 70% and 80%, making these two genotypes unique. This assessment in relation to SWC shows that the factor soil is an important one when it comes to water deficit stress experiments.
The several genotypes studied presented different velocities to reach PMWS. These differences may be partly related with the developmental stage of each genotype, since plants with flowers and pods have more water needs than the ones that do not possess them. Although this is a viable hypothesis we can also discuss the different water loss rates and stress threshold based on physiological parameters and it is this hypothesis that is discussed throughout this section.
The first genotypes reaching critical stress (taken as a decrease of Fv/Fm below 0.75) were L. sativus, L. clymenum and L. ochrus at the end of 23 days of stress (i.e., water withholding), followed by L. cicera that hold for 30 days. Genotypes L. sylvestris, L. aphaca, L. odoratus and the low -ODAP variety from Canada of L. sativus (LS 87124) only reach PMWS in average 37 days after water withholding. Genotypes that withstood longer the stress period were the variety from Raipur of L. sativus and L. latifolius with respectively PMWS at 43 and 60 days of water withholding. Based upon these differences we can distinguish L. latifolius and the Raipur variety of L. sativus apart. Although developmental stage of each genotype is a factor to be considered it is not the main cause for the differences found, since the variety from Raipur of L. sativus belonged to the flowered group.
23 L. aphaca 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. cicera 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. clymenum 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. latifolius 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. ochrus 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. odoratus 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. sativus (L174-A) 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. sativus (LS 87124) 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. sativus (Raipur) 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % ) L. sylvestris 40 50 60 70 80 90 100 0 20 40 60 80 100 SWC (%) R W C ( % )
Table 4 shows mean values for fluorescence parameters studied at the PMWS when compared with control values for each genotype. There are significant lower values for stressed plants for all genotypes and in all three parameters measured. The analysis of the low Fv/Fm values in the PMWS when compared with control values shows that all genotypes were subjected to a severe stress. If PI values are subject to a careful observation there is an obvious difference between PMWS and control values greater than for Fv/Fm. Taking into account that PI reflects the functionality of both photosystems and gives quantitative
Fig. 6 – Relationship between plant relative water
content (RWC %) and soil water content (SWC %) for each Lathyrus genotype studied.