Association between G316A growth hormone polymorphism
and economic traits in pigs
Danielle Assis de Faria
1, Simone Eliza Facioni Guimarães
1, Paulo Sávio Lopes
1, Aldrin Vieira Pires
2,
Samuel Rezende Paiva
3, Bruna Pena Sollero
1and Amauri Arias Wenceslau
41
Departamento de Zootecnia, Universidade Federal de Viçosa, Viçosa, MG, Brazil.
2
Sistema FAESA de Ensino, Rod. Serafim Derenzi, Vitória, ES, Brazil.
3
Laboratório Genética Animal, Embrapa Recursos Genéticos e Biotecnologia, Brasília, DF, Brazil.
4Departamento de Ciências Agrárias, Universidade Estadual de Santa Cruz, Ilhéus, BA, Brazil.
Abstract
The association between G316A growth hormone polymorphism and quantitative traits was investigated in an F2 population of pigs. Association analyses were performed using a statistical model that included genotype, sex, batch and sex by genotype interaction as fixed effects and sire as random effect. The polymorphism was associated with the number of right teats (p = 0.03), heart weight (p = 0.04), lung weight (p = 0.05), carcass length determined by the Brazilian carcass classification method (p = 0.04), picnic shoulder weight (p = 0.07), jowl weight (p = 0.01), pH 24 h after slaughtering (p = 0.03) and drip loss (p = 0.01). Interaction between genotype and sex was observed for six per-formance traits. The additive effect was significant (p < 0.10) for heart weight, jowl weight and pH 24 h after slaughter-ing. The effect of dominance was significant (p < 0.05) for number of right teats, heart weight, carcass length, picnic shoulder weight and pH 24 h after slaughtering. This study shows that the growth hormone gene is a potential candi-date for investigating the phenotypic variation of quantitative traits in pigs, and suggests its possible application in breeding programs.
Key words:candidate genes, divergent cross, molecular marker, sex x genotype interaction, swine.
Received: June 2, 2005; Accepted: April 4, 2006.
Introduction
Advances in different areas of animal production (management, nutrition, environment, sanitary control and genetic breeding) have led to improvements in the pig pro-duction chain. Genetic breeding of herds have been carried out by selecting animals with high production potential based on their phenotype. One alternative for the selection of the best animals to be used as parents of the next genera-tion is to identify genes or loci controlling economically important traits and to incorporate this information into tra-ditional breeding methods.
In the last few years there have been many important advances in molecular genetics and biotechnology. Pigs represent a good animal model for genetic studies due to their short gestational period, large number of piglets per litter and short generation interval. Most of the markers de-veloped and used by the pig breeding industry have been identified by candidate gene analysis. In these studies, an
appropriate population design is important, with the F2 de-sign being the most widely used (Liu, 1998). The mating design for a reference population should determine the rela-tionships between polymorphic markers and traits of inter-est.
Growth hormone (GH) is involved in the regulation of growth and metabolism of different tissues during ana-bolic and cataana-bolic processes, it may have a direct or indi-rect effect on the target tissues. The effects of growth hormone, mediated by insulin-like growth factor I (IGF-I), on muscle and skeletal tissue growth are similar to those of insulin, whereas its direct action on carbohydrate metabo-lism and lipolysis is antagonistic to that of insulin, once growth hormone, along with cortisol, exerts a synergistic effect on lipolysis and diabetogenesis. Most, if not all the effects of growth hormone on muscle development are me-diated by IGF-I, and the functions performed by GH make this hormone a candidate gene for various production traits. The pig growth hormone gene polymorphism (G316A) was identified by sequencing native Brazilian Piau boars and sows of a commercial line (GenBank acces-sion number DQ339698), corresponding of the parental www.sbg.org.br
Send correspondence to Samuel Rezende Paiva. Embrapa Recur-sos Genéticos e Biotecnologia, 70770-900 Brasília, DF, Brazil. E-mail: [email protected].
generation of the present study population. This polymor-phism is located in the first intron of the gene, 316 bp down-stream according to the reference sequence (GenBank accession number M17704) published by Vize and Wells (1987).
The objective of the study described in the present pa-per was to analyse the G316A polymorphism present in the growth hormone gene sequence using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method and to determine any statistical associations between the G316A polymorphism and quantitative traits in a segregating F2 pig population. This polymorphism has not yet been studied in pigs and it is important to uncover its effects on the quantitative traits in this population. An addi-tional goal in our study was to demonstrate the potential for genetic analyses in naturalized Brazilian pigs, which has not yet been properly explored.
Material and Methods
Experimental population
The F2 pig population was generated by crossing two native Brazilian Piau boars with 18 commercial sows (Lan-drace x Large White x Pietran) selected for growth rate and backfat thickness. The F1 generation consisted of 106 sows and 134 boars (Band et al., 2005; Carmo et al., 2005). Twelve boars from different litters were randomly selected from the 134 F1 boars and mated by natural breeding with 54 F1 sows to produce the F2 generation. The F2 generation consisted of approximately 840 offspring divided into five batches according to the season in which they were born. Ten days after birth, the males were castrated.
Phenotypes and DNA extraction
Sixty days after birth the pigs were transferred to col-lective pens where they were maintained until 77 days of age and then moved to individual cages to measure feed conversion (feed intake/weight gain), average daily gain and feed intake over 28 days (from 77 to 105 days of age). All measurements were performed on the F2 animals and are presented in Table 1, along with standard errors and the total number of pigs analyzed for each trait.
Around 65 kg of living weight (64.71 ± 0.24), the F2 offspring pigs were fasted for 20 h, electrically stunned (300 V/5 s) and slaughtered at the farm’s abattoir and bled by cardiac puncture. Data regarding carcass cut traits were obtained by cooling the right half of each carcass at 4 °C for 24 h, dissected and analyzed as described by Nascimento and Mota (2000). Loin samples (longissimus dorsi) were removed from the animals for the analysis of meat quality, which was performed at the Meat Laboratory of the Depart-ment of Food Technology, Federal University of Viçosa (UFV), Viçosa, Minas Gerais (MG), Brazil. The number of observations, means and standard errors for the analyzed traits are shown in Table 2. All the F2 population results
re-garding performance, carcass yield and meat quality traits were described by Lopeset al. (2002).
Extraction, manipulation and subsequent analysis of genomic DNA were carried out at the Laboratory of Ani-mal Biotechnology (LABTEC), Department of AniAni-mal Science, UFV. Blood samples were collected from paren-tal, F1 and F2 animals by venipuncture of the sinus
orbitalisat 77 days of age. Genomic DNA was extracted
from blood using phenol:chloroform purification after treatment with proteinase K (Sambrooket al., 1989). The DNA samples were diluted to 25 ng/mL in TE buffer con-taining 10 mM Tris-HCl, pH 8.0, and 1 mM EDTA, pH 8.0, and stored at 4 °C. The DNA stock solution (concentrated material) was stored at -20 °C.
PCR and RFLP analysis
The primer pair used for the amplification of the growth hormone gene region was designed from the se-quence published by Vize and Wells (1987), (GenBank ac-cession number M17704) using Primer 3 software (Rozen and Skaletsky, 2000). The F1 - TTATCCATTAGCAC ATGCCTGCC and R1 - CTGGGGAGCTTACAAACTC CTT primer pair was used for amplification of the fragment containing the G316A polymorphism. This primer pair am-plifies a 604-bp fragment comprising the promoter region, first exon and part of the first intron of the growth hormone gene.
Restriction analysis used the (G®A) base
substitu-tion mutasubstitu-tion at posisubstitu-tion 316 recognized by the FokI restriction enzyme (Invitrogen Inc.). The specific DNA fragment was amplified by PCR in a MJ Research thermo-cycler (model PTC-100, Watertown, MA, USA) using a re-action mixture containing 4.0 pmol of each primer, 0.2 mM of each dNTP, 1.0 mM MgCl2, 50 mM Tris-KCl, 1.0 unit
Taq DNA polymerase and 25 ng of genomic DNA in a final
Table 1- Number of F2 offspring (n) analyzed, mean and standard error (SE) with each performance trait.
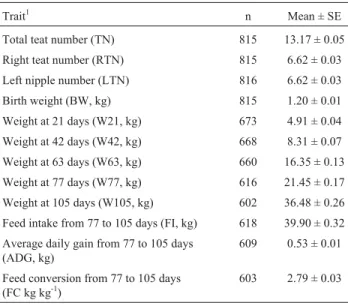
Trait1 n Mean ± SE
Total teat number (TN) 815 13.17 ± 0.05
Right teat number (RTN) 815 6.62 ± 0.03
Left nipple number (LTN) 816 6.62 ± 0.03
Birth weight (BW, kg) 815 1.20 ± 0.01
Weight at 21 days (W21, kg) 673 4.91 ± 0.04
Weight at 42 days (W42, kg) 668 8.31 ± 0.07
Weight at 63 days (W63, kg) 660 16.35 ± 0.13
Weight at 77 days (W77, kg) 616 21.45 ± 0.17
Weight at 105 days (W105, kg) 602 36.48 ± 0.26
Feed intake from 77 to 105 days (FI, kg) 618 39.90 ± 0.32
Average daily gain from 77 to 105 days (ADG, kg)
609 0.53 ± 0.01
Feed conversion from 77 to 105 days (FC kg kg-1)
volume of 20mL. The amplification program consisted of
initial denaturation at 94 °C for 5 min followed by 35 am-plification cycles of strand denaturation at 94 °C for 1 min, primer annealing at 64 °C for 1 min and fragment extension
at 72 °C for 1 min, followed by a final extension step at 72 °C for 5 min. The amplified products were loaded in a 8% (w/v) polyacrylamide gel, stained with silver nitrate. In each gel was also loaded 1mL of a 1-kb ladder (Invitrogen)
as a standard.
After amplification of the region containing the poly-morphism, 7 mL of the PCR product was digested with
about 5 units of theFokI restriction endonuclease in a mix-ture containing 1.5mL buffer, 0.15mL bovine serum
albu-min (10 mg/mL) and 4.85 mL Milli-Q water in a final volume of 15mL. The mixture was incubated for 1 h at
37 °C, followed by inactivation at 65 °C for 20 min. The di-gestion product was also visualized on silver-stained 8% (w/v) polyacrylamide gel.
F1 animals were genotyped by PCR-RFLP to identify the alleles inherited from the parental generation. The mu-tation was first identified by sequencing, when the poly-morphism was confirmed, a total of 402 F2 animals was genotyped.
Statistical analysis
The database consisted of two basic files, one con-taining the phenotypic measurements of all evaluated traits in about 600 animals and the other the marker genotypes coded as GG (homozygous wild-type genotype), AA (ho-mozygous G316A mutant genotype) or GA (heterozygous wild-type/mutant genotype). A total of 402 animals were genotyped for the G316A polymorphism. The association between the genotypes and evaluated traits was analyzed statistically using the SAS program (1998) and the follow-ing model:
yijklm =a+Gi +Sj +GSij+Bk+sl+(Cijklm-C b) +eijklm
whereyijklm= trait observed in pig m, of genotypei, sexj,
from batchk, and sire l;a= constant inherent to all observa-tions;Gi= fixed effect of genotypei(i= 1, 2, 3);Sj= fixed
effect of sexj,j= 1, 2 (1 = castrated males, 2 = females);
GSij= interaction between genotypeiand sexj;Bk= fixed
effect of batchk,k= 1, 2, 3, 4 and 5;sl= random effect of
sire l (l = 1,... );b= linear regression coefficient of theyijklm
trait in relation to the covariable; Cijklm= value of the
covariable observed in animalm, of genotypei, sexj, from batchk, and sirel;C= mean value of the covariable; and
eijklm= random error associated with each observation.
The following covariables were used: litter size at birth for the birth weight trait; litter size at weaning for weight at 21, 42, 63, 77 and 105 days of age and weight at slaughter; weight at 77 days for feed intake, average daily gain and feed conversion; carcass weight for all carcass traits; weight of the cooled right carcass side for cut traits; and age at slaughter for meat quality traits. No covariable was used for the total number of left and right teats.
Covariances between genetic and environmental ef-fects (permanent and temporary) and between
environmen-Table 2- Number of F2 offspring analyzed (n), mean and standard error (SE) of carcass, carcass cut and meat quality traits.
Trait n Mean ± se
Slaughter weight (SW, kg) 529 64.71 ± 0.24
Slaughter age (SA, days) 538 147.81 ± 0.43
Carcass weight (CW, kg) 540 53.60 ± 0.23
Carcass yield (CY, %) 526 81.99 ± 0.09
Right half carcass weight (RHCW, kg) 839 26.84 ± 0.09
Carcass length by the American carcass classification method (MLC, cm)
540 71.67 ± 0.14
Midline backfat thickness at last rib (LR, mm)
549 19.81 ± 0.21
Midline backfat thickness between last and next to last but one lumbar vertebra (LL, mm)
547 28.51 ± 0.25
Midline backfat thickness above the last lumbar vertebra (L, mm)
495 45.01 ± 0.09
Backfat thickness at last rib, 6.5 cm from the midline (P2, mm)
547 16.81 ± 0.16
Loin depth (LD, mm) 495 43.80 ± 0.20
Loin eye area (LEA, cm2) 499 26.30 ± 0.17
Total ham weight (THW, kg) 543 7.28 ± 0.04
Skinless and fatless ham weight (HW, kg)
543 4.99 ± 0.03
Total Boston shoulder weight (TBSW, kg)
549 2.34 ± 0.02
Skinless and fatless Boston shoulder weight (BSW, kg)
544 1.69 ± 0.01
Total picnic shoulder weight (TPSW, kg)
545 4.87 ± 0.03
Skinless and fatless picnic shoulder weight (PSW, kg)
548 2.70 ± 0.02
Total (bone-in) loin weight (TLW, kg) 540 3.47 ± 0.02
Boneless loin weight (LW, kg) 542 1.02 ± 0.01
Bacon weight (BCW, kg) 541 2.69 ± 0.02
Bacon depth (BCD, mm) 539 24.92 ± 0.29
Rib weight (RW, kg) 548 1.52 ± 0.01
Sirloin weight (SLW, kg) 546 0.22 ± 0.02
Small intestine length (SIL, m) 544 18.42 ± 0.08
PH 45 min after slaughter (pH45) 543 6.50 ± 0.01
PH 24 h after slaughter (pHu) 555 5.71 ± 0.01
Drip loss (DL, %) 557 3.19 ± 0.07
Cooking loss (CL, %) 550 32.63 ± 0.11
Total loss (TL, %) 439 34.23 ± 0.13
Intramuscular fat ( IMF, %) 504 1.55 ± 0.03
Shear force (SF, g per 1.2 cm) 431 5550.63 ± 42.10
Redness (A) 485 0.67 ± 0.03
Yellowness (B) 491 6.62 ± 0.02
Chroma (c) 429 6.70 ± 0.03
tal effects of different animals were considered to be zero. Association analyses were evaluated by the F-test between molecular markers and trait measures and between geno-typevs.sex interaction. In both analyses, genotype means were compared by the t-test when the interaction was sig-nificant by the F-test.
The additive effect was determined by regression of the values obtained for each genotype compared to the number of non-mutated alleles. The effect of dominance was determined by linear contrasts between genotype means and the t-test was used to evaluate the significance of the dominance effect.
Results
Digestion of the PCR product and subsequent electro-phoresis revealed three genotypes, with genotype GA showing fragments of 604, 345 and 259 bp, genotype GG fragments of 345 and 259 bp and genotype AA a single 604-bp fragment (Figure 1). In the parental generation both boars and four sows were heterozygous (GA) for the G316A polymorphism and 14 sows had the wild-type ho-mozygous genotype (GG). A total of 402 F2 offspring were genotyped for the G316A polymorphism. The genotype frequency was GG = 0.5672 (228 offsprings), GA = 0.4005 (161 offsprings) and AA = 0.3230 (13 offspring). The gene frequency for the G allele was 0.7674 and 0.2326 for the A allele.
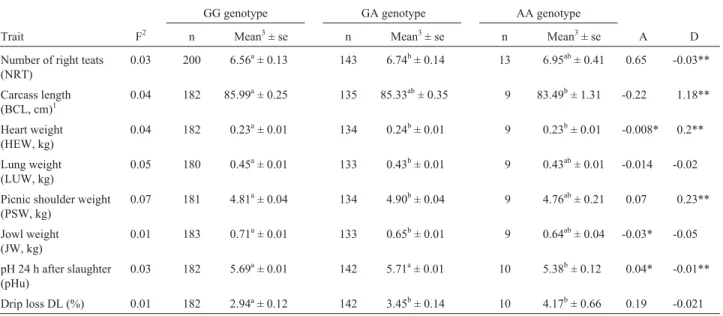
The significance level, mean and standard error, and additive and dominance effects of the genotypes for each significant trait are shown in Table 3. The G316A polymor-phism was found to be associated with the number of right teats, heart weight, lung weight, carcass length measured by the Brazilian carcass classification method, total picnic shoulder weight, jowl weight, pH 24 h after slaughtering (pHu) and drip loss (DL). According to these results, AA animals presented a larger number of teats than the other genotypes. Mean carcass length measured by the Brazilian method was greater in GG and smaller in AA offspring. Mean total picnic shoulder weight was higher in heterozy-gous offspring compared to the other two genotypes.
Re-garding significant meat quality traits (pHu and DL), AA offspring showed lower pHu values and a higher DL mean, whereas a lower DL mean was observed for the GG geno-type and higher pHu values were noted in GG and heterozy-gous offspring.
A significant additive effect of the G316A polymor-phism was present for heart weight (HEW), jowl weight (JW) and pHu (Table 3). The absence of a significant domi-nance effect was only observed for lung weight (LUW), JW and DL.
The genotype (x) sex interaction was significant for some traits in F2 pigs with the polymorphism in theporcine
growth hormone(PGH) gene . Figure 2 shows the level of
significance of the genotype x sex interaction and trait means obtained for F2 males and females within each geno-type class. For the males, the wild homozygous genogeno-type (GG) was only significantly higher than the other geno-types in regard to the carcass length as measured by the Brazilian carcass classification method (BCL) trait. For the females, the homozygous G316A mutant genotype (AA) was significantly higher in respect to backfat thickness af-ter the last rib 6.5 cm from the midline (the P2 trait), a trait related to fat deposition.
Discussion
Three main hypotheses can be postulated to explain the observed association between the change in nucleotides represented by the G316A polymorphism and some quanti-tative traits. One hypothesis is that the G316A polymor-phism is directly responsible for the variation in the observed traits, with this polymorphism being involved in the synthesis of a more or less efficient protein (i.e.PGH) or interfering at various levels of expression such as transcrip-tion, processing, transport or the half-life of the circulating hormone. The G316A polymorphism is located at intron I on a non-expressed region of the gene. Although the proba-bility of exerting biological effects on gene expression is higher for polymorphisms located on exonic regions, it is possible that polymorphisms located on unexpressed re-gions could interfere in gene expression, e.g.
phisms located on enhancer regions consisting of coded or uncoded sequences that physically act on the regulation of gene expression.
A second hypothesis is that the polymorphism is in linkage disequilibrium with another polymorphism which is the true causative agent of the observed variations in the analyzed traits. Other important genes that might be associ-ated with production traits are present on pig chromosome 12 (SSC12), e.g. the acetyl CoA carboxylase gene and
apolipoprotein Hgene. In this case, the polymorphism
de-scribed on the first intron of thePGHgene would be acting as a marker for the true site, which in turn would explain the associations. This hypothesis could be tested by the analy-ses of the frequency of these genes in the population de-scribed in our present paper.
A third hypothesis is the existence of false-positive associations. The statistical model used by us assumed that
only polymorphisms and fixed and random effects influ-ence trait means. However, it is known that phenotypic variation is controlled by many factors, including poly-genic and environmental variations. Another fact that could possibly have resulted in false-positive associations is the limited number of observations for genotype AA (0.3230). In order to minimize the effect of this third hypothesis, we applied the same statistical tests excluding genotype AA and the results were the same as those observed when the three genotypes were analyzed, based on which we main-tained the AA animals in the analyses.
Other studies have shown the association of PGH with economic traits in swine (e.g., Cheng et al., 2000; Knorret al., 1997) and the probability of false positives in this study is small so the false-positive hypothesis can prob-ably be discarded, although further studies are necessary to
confirm which of the remaining hypotheses best explain our results.
Despite the importance for metabolism in general, the effect of the G316A polymorphism on the total variation of traits showing an association was relatively small. No asso-ciation was observed between allelic variants and perfor-mance traits influenced by PGH, such as different feed intake, feed conversion rates or body weights (Table 3).
Our finding that the non-mutant homozygous geno-type (GG) detected in commercial sows of the parental gen-eration was responsible for greater carcass length means, lower drip loss and higher mean pH 24 h after slaughtering can be explained by the fact that commercial animals have been selected over generations for higher meat production, greater carcass length and greater carcass lean meat yield. However, it does not explain the lower drip loss, since mercial pig breeds tend to show higher drip losses com-pared to naturalized breeds. According to correlation analyses (data nor shown), pHu and DL are negatively cor-related (-0.13; p < 0.01), a finding confirmed in our study. Several studies have associated PGH genotypes with traits of interest in some pig breeds. Knorret al. (1997) studied crosses between Pietran and Meishan pigs and found allelic variants of PGH to be associated with eight carcass composition traits, all of which were related to fat deposition. In contrast, in our study the genotypes were only associated with fat deposition traits when the effect of sex was included in the analysis, with the genotypes being associated with three backfat thickness measurements (LL, ETO and BCD; Figure 2).
Divergent results, even in comparison to the present study, have been reported in investigations associating quantitative traits and allelic variations in the PGH gene. Chenget al. (2000), analyzing Duroc, Landrace and Tao-Yuan pigs found positive associations between quantitative traits and PGH allelic variations only for the Tao-Yuan breed, with allelic variants influencing average daily gain and the efficiency of weight gain.
Another important factor is that studies of candidate genes for trait improvement are normally carried out based on variations in quantitative traits that are influenced by various genes and by the environment. Thus, the associa-tions found in experimental populaassocia-tions should not be ex-trapolated before being tested and validated in the target population. Therefore, the detection and confirmation of the G316A polymorphism in a commercial population is important, because it affects different traits included in ani-mal breeding programs. One of these traits is meat quality (pHu and DL) which is difficult to measure, by traditional selection methods. One alternative for the selection of these traits is marker-assisted selection using, for example, the genotypes here described.
Sex hormones (estrogen, testosterone, etc.), along with PGH, may influence traits in castrated males and fe-males in different ways. Few correlation studies about the amount of circulating PGH and the sex of the animal, are available; most investigations associate this parameter with age and animal breed. Dubreuilet al. (1987) reported that sex did not influence basal PGH secretion in growing pigs. In our study there was a positive association between
Table 3- Number of F2 offspring anayzed (n), trait means (mean) with standard errors (SE), additive (A) and dominance (D) effects obtained for each genotype of the G316A polymorphism for significant performance, carcass, carcass cut and meat quality traits in F2 pigs.
GG genotype GA genotype AA genotype
Trait F2 n Mean3± se n Mean3± se n Mean3± se A D
Number of right teats (NRT)
0.03 200 6.56a± 0.13 143 6.74b± 0.14 13 6.95ab± 0.41 0.65 -0.03**
Carcass length
(BCL, cm)1
0.04 182 85.99a± 0.25 135 85.33ab± 0.35 9 83.49b± 1.31 -0.22 1.18**
Heart weight (HEW, kg)
0.04 182 0.23a± 0.01 134 0.24b± 0.01 9 0.23b± 0.01 -0.008* 0.2**
Lung weight (LUW, kg)
0.05 180 0.45a± 0.01 133 0.43b± 0.01 9 0.43ab± 0.01 -0.014 -0.02
Picnic shoulder weight (PSW, kg)
0.07 181 4.81a± 0.04 134 4.90b± 0.04 9 4.76ab± 0.21 0.07 0.23**
Jowl weight (JW, kg)
0.01 183 0.71a± 0.01 133 0.65b± 0.01 9 0.64ab± 0.04 -0.03* -0.05
pH 24 h after slaughter (pHu)
0.03 182 5.69a± 0.01 142 5.71a± 0.01 10 5.38b± 0.12 0.04* -0.01**
Drip loss DL (%) 0.01 182 2.94ª ± 0.12 142 3.45b± 0.14 10 4.17b± 0.66 0.19 -0.021
1Measured by the Brazilian carcass classification method.
2Probability of statistical significance for F test.
3
Means followed by different letters in the same line were different by the t-test (p < 0.05). *Significant by the F-test (p < 0.10).
G316A genotype variation and sex, but since this polymor-phism is located on an intron it was not possible confirm wether or not it causes higher or lower PGH secretion in males or females. However, in our study the males have been castrated and would have shown lower testosterone levels than boars.
In our study, a significant dominance effect was ob-served for many traits (Table 3). It is likely that the lack of significant additive and dominance effects for the drip loss and lung weight traits was a consequence of the fact that in our study only a few pigs had the AA genotype, which might have interfered with the estimates and the signifi-cance of the effects. For example, we analyzed drip loss in 9 AA pigs, 142 GA pigs and 182 GG pigs. Thus, these effects should be tested for using larger commercial populations in order to validate any gene effects.
Since parents transfer their genes and not their geno-types to their offspring, the advantage of a direct marker would be that it presents a higher additive than dominance effect, since effects of dominance are caused by the interac-tion between alleles at the same locus and these interacinterac-tions are rearranged in each generation. Thus, it is important to know whether any polymorphism effects are due to addi-tive action or to dominance deviation.
Positive associations between allelic variants and breed have been reported in investigations analyzing differ-ent breeds. In our study we evaluated a Brazilian natural-ized breed with a different origin from that of the breeds used in European and North American studies. However, this was probably not the main reason for the positive asso-ciations between genotypes and traits here studied. The de-scribed PGH G316A mutation is not exclusive to the Piau breed and has also been detected in commercial females re-sulting from Landrace x Large White x Pietran crosses, breeds widely used in candidate gene analysis.
Our study demonstrates the potential of the PGH gene as a candidate for the investigation of quantitative traits in pigs and suggests that the G316A polymorphism should be tested in commercial populations for its possible applica-tion in marker-assisted selecapplica-tion programs in pigs.
Acknowledgments
The authors wish to thank the Brazilian agencies FAPEMIG, CNPq and CAPES for financial support and Dr. Luiz L. Coutinho (USP/ESALQ) and Dr. Marta Gui-marães (EMBRAPA/Gado de Leite) for help with the gene sequencing and sequence editing.
References
Band GO, Guimarães SEF, Lopes PS, Peixoto JO, Faria DA, Pires AV, Figueiredo FC, Nascimento CS and Gomide LAM (2005) Relationship between the Porcine Stress Syndrome gene and carcass and performance traits in F2pigs resulting
from divergent crosses. Genet Mol Biol 28:92-96.
Carmo FMS, Guimarães EF, Lopes PS, Pires AV, Guimarães MFM, Silva MVGB, Schierholt AS, Silva KM and Gomide LAM (2005) Association of MYF5 gene allelic variants with production traits in pigs. Genet Mol Biol 28:363-369. Cheng WTK, Lee CH, Hung CM, Chang TJ and Chen CM (2000)
Growth hormone gene polymorphisms and growth perfor-mance traits in Duroc, Landrace and Tao-Yuan pigs. Theriogenology 54:1226-1237.
Dubreuil P, Pelletier G, Petitclerc D, Lapierre H, Couture Y, Brazeau P, Gaudreau P and Morisset J (1987) Influence of age and sex on basal secretion of growth hormone (GH) and on GH-induce release by porcine GH-releasing factor pGRF (1-29 NH2) in growing pigs. Dom Animal Endocri 4:229-307.
Knorr C, Moser G and Geldermann H (1997) Associations of GH gene variants with performance traits in F2 generations of European wild boar, Pietran and Meishan pigs. Anim Genet 28:124-128.
Liu BH (1998) Computational tools for study of complex traits. In: Paterson AH (ed) Molecular Dissection of Complex Traits. CRC Press, Nova York, pp 43-80.
Lopes PS, Guimarães SEF, Pires AV, Soares MAM, Carmo FMS, Martins MF, Benevenuto Júnior AA and Gomide LAM (2002) Results of performance, carcass yield and meat qual-ity traits of F2 crosses between Brazilian native and com-mercial pigs for QTL mapping. Proceedings of 7th World Congress on Genetics Applied to Livestock Production, Montpellier, pp 155-158.
Nascimento JD and Mota EO (2000) Dissecção de carcaças de suínos na Agroceres PIC. Informe Técnico, AGROCERES PIC, Rio Claro, 5 pp.
Rozen S and Skaletsky HJ (2000) Primer3 on the WWW for gen-eral users and for biologist programmers. In: Krawetz S and Misener S (eds) Bioinformatics Methods and Protocols: Methods in Molecular Biology. Humana Press, Totowa, pp 365-386.
Sambrook J, Fritsch EF and Maniatis T (1989) Molecular Clon-ing: A Laboratory Manual. 2nd edition. Cold Spring Harbor Laboratory, New York, 564 pp.
SAS (1998) Institut Inc. User’s Guide. Statistical Analysis Sys-tem. Cary, NC, USA.
Vize PD and Wells JRE (1987) Isolation and characterization of the porcine growth hormone gene. Gene 55:339-344.