Disparate Effects of Mesenchymal Stem Cells in Experimental Autoimmune Encephalomyelitis and Cuprizone-Induced Demyelination.
Texto
Imagem
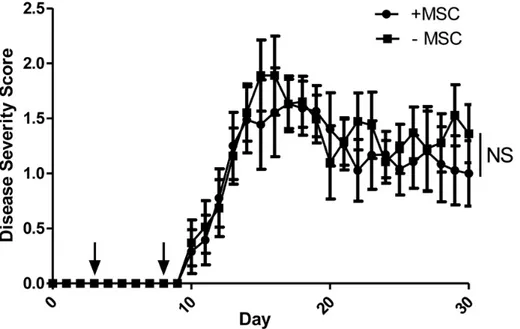
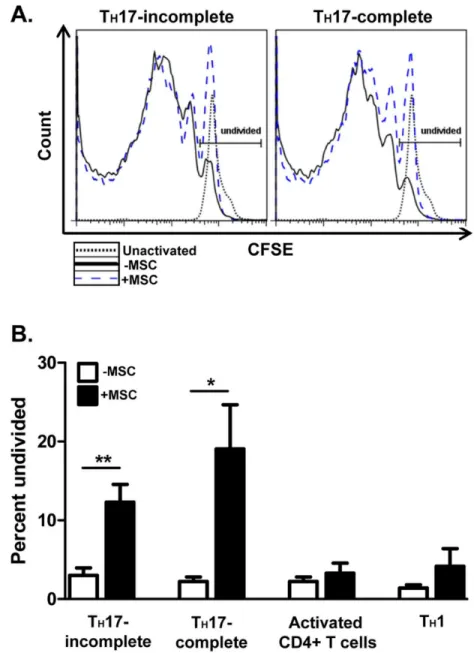
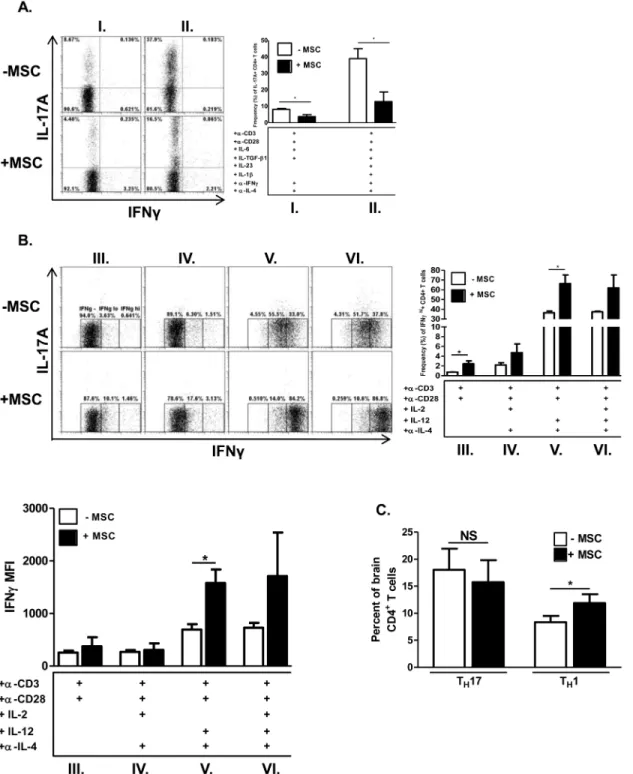

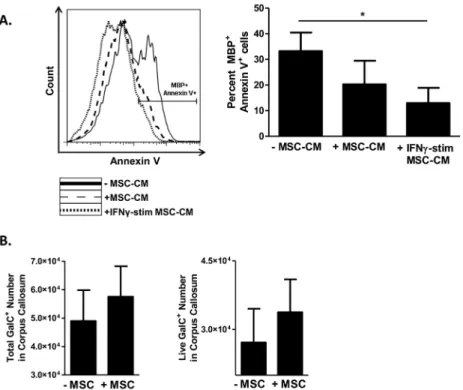
Documentos relacionados
Em suma, poder-se-ia dizer que o têrmo ciência é aplicável à administração pública sempre que alguns de seus métodos possam seguir princípios cinetíficos,
economy since 1980 to 2010, such as: (i) rising skill premium; (ii) increase in income and wealth Gini coefficient; (iii) decrease in the wealth share owned by the bottom 90%
Por ser a renúncia fiscal um ato individual de vontade do poder público de conceder/reconhecer, a determinado contribuinte, o direito ao não cumprimento da obrigação principal,
Ao longo da pesquisa buscou-se compreender se os processos de re-existência de agricultores familiares em áreas de domínio sucro-energético estariam possibilitando a
Dessa forma, os resultados obtidos indi- caram que o total da "Escala de Comporta- mento" discriminou significativamente entre as idades de 4, 5 e 6 anos, em ambos os
Para concluir, apesar do pouco tempo para a realização da investigação, penso que foi uma ótima experiência, não só para o grupo, mas também para mim, pois aprendi de igual
Revela-se de particular interesse para este estudo, O Decreto - Lei 115- A/98, artigo 5º, no que diz respeito às finalidades dos Agrupamentos de escolas, na enunciação
