Mesenchymal stem cells from patients
with chronic myeloid leukemia do not
express BCR-ABL and have absence of
chimerism after allogeneic bone
marrow transplant
Centro Regional de Hemoterapia, Departamento de Clínica Médica, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brasil
R.C.V. Carrara, M.D. Orellana, A.M. Fontes, P.V.B. Palma, S. Kashima, M.R. Mendes, M.A. Coutinho, J.C. Voltarelli and D.T. Covas
Abstract
Bone marrow is a heterogeneous cell population which includes hematopoietic and mesenchymal progenitor cells. Dysregulated he-matopoiesis occurs in chronic myelogenous leukemia (CML), being caused at least in part by abnormalities in the hematopoietic progeni-tors. However, the role of mesenchymal stem cells (MSCs) in CML has not been well characterized. The objectives of the present study were to observe the biological characteristics of MSCs from CML patients and to determine if MSCs originate in part from donors in CML patients after bone marrow transplantation (BMT). We analyzed MSCs from 5 untreated patients and from 3 CML patients after sex-mismatched allogeneic BMT. Flow cytometry analysis revealed the typical MSC phenotype and in vitro assays showed ability to differen-tiate into adipocytes and osteoblasts. Moreover, although some RT-PCR data were contradictory, combined fluorescence in situ hybrid-ization analysis showed that MSCs from CML patients do not express the bcr-abl gene. Regarding MSCs of donor origin, although it is possible to detect Y target sequence by nested PCR, the low frequency (0.14 and 0.34%) of XY cells in 2 MSC CML patients by fluorescence in situ hybridization analysis suggests the presence of contaminant hematopoietic cells and the absence of host-derived MSCs in CML patients. Therefore, we conclude that MSCs from CML patients express the typical MSC phenotype, can differentiate into osteogenic and adipogenic lineages and do not express the bcr-abl gene. MSCs cannot be found in recipients 12 to 20 months after BMT. The influence of MSCs on the dysregulation of hematopoiesis in CML patients deserves further investigation.
Correspondence
D.T. Covas
Centro Regional de Hemoterapia FMRP, USP
Rua Tenente Catão Roxo, 2501 14051-140 Ribeirão Preto, SP Brasil
Fax: +55-16-2101-9309 E-mail: [email protected]
Research supported by FAPESP (Nos. 1998/14247-6 and 01/07308-3).
Received December 14, 2005 Accepted October 9, 2006
Key words
•Mesenchymal stem cells •Chimerism
Introduction
Bone marrow (BM) is composed of at least two systems, i.e., the hematopoietic tissue proper and the stroma (1,2). Mesenchymal stem cells (MSCs) are important constituents of this microenvironment and are character-ized as adult, non-hematopoietic stem cells (HSCs) which, after an adequate stimulus, can differentiate morphologically and function-ally into different cell lines including the stroma, which gives support to hematopoiesis, adipo-cytes, chondroadipo-cytes, myoadipo-cytes, astroadipo-cytes, tenocytes, and hepatocytes (3,4).
In addition, studies by our group on the transcriptome of BM MSCs have shown both differences and similarities compared to CD34+ hematopoietic BM stem cells.
About 1/3 of the most expressed gene prod-ucts were present in both cell types, whereas 2/3 showed exclusive over-expression in one of the cell types (5).
Chronic myeloid leukemia (CML) is a myeloproliferative disease originating in multipotent HSCs which acquire the recip-rocal translocation t(9;22)(q34;q11) charac-terized cytogenetically by the presence of the Philadelphia (Ph1) chromosome. This translocation intercalates portions of the c-abl protooncogene in chromosome 9 with the c-bcr gene in chromosome 22, forming a hybrid bcr-abl gene that codes for the BCR-ABL oncoprotein (6-8).
More recent studies have suggested a more primitive cellular origin for the t(9;22) translo-cation (9,10). Gunsilius et al. (9) identified the bcr-abl fusion gene in a more primitive pro-genitor cell (CD34+ CD31+) than the
hemato-poietic cell which had the characteristics of a hemangioblast when isolated in vitro from the BM of patients with CML.
Fang et al. (10) isolated from the BM of patients with CML a population of even more primitive progenitor cells defined as Flk1+
CD31- CD34- which also carried the bcr-abl
fusion gene, and demonstrated in in vitro stud-ies that, starting from cultures of a single cell,
this population can give origin to both he-matopoietic and endothelial cells, with both populations being malignant and able to pro-duce CML when injected into SCID mice.
The objective of the present study was to determine the possible existence of the bcr-abl fusion gene in the MSCs of the BM of patients with CML. This would permit us to conclude the presence of this mutation in another more primitive progenitor cell popu-lation in addition to hemangioblasts. To this end, untreated MSCs were isolated from the BM of CML patients during the diagnostic phase and characterized phenotypically and cytogenetically.
In view of the controversial literature re-ports about the transplantation potential of MSCs (11-15), the degree of stromal micro-chimerism was evaluated in fractions of MSCs isolated from the BM of CML patients submit-ted to allogeneic bone marrow transplantation (BMT), in order to determine the migration and stromal reconstitution potential of donor MSCs in recipients submitted to the myeloab-lative conditioning regimen. This analysis was carried out by isolating MSCs obtained from 3 female patients who had received a BMT from a male donor, using nested PCR and fluores-cence in situ hybridization (FISH).
Patients, Material and Methods
Patients
Twelve untreated CML patients were re-cently diagnosed Ph+ in the chronic phase, and
3 were female patients with CML who had received an allogeneic BMT from male do-nors at the BMT service of the University Hospital, Faculty of Medicine of Ribeirão Preto. In order to investigate the biological characteristics of MSCs we isolated BM-MSC from 5 untreated patients. The other 7 un-treated patients were used to obtain purified HSCs CD34+ from the BM and peripheral
these cell populations. We also analyzed the MSCs and hematopoietic cells of 3 female patients with CML after allogeneic BMT, and mononuclear cells from peripheral blood. The Ethics Committee of the Institution approved the study and all patients gave written in-formed consent to participate.
Characteristics of the patients
The clinical characteristics of the pa-tients submitted to BMT are presented in Table 1. Mean patient age was 25.7 years (range: 24 to 29 years). The mean age of the untreated and non-transplanted CML patients was 37.4 years (range: 15 to 64 years). The cellularity of the BM of these untreated pa-tients was 11-130 x 103/mm3 and the
expres-sion of the bcr-abl gene was demonstrated in all CML patients by RT-PCR (data not shown).
Characteristics of the donors
All donors were related to the recipients, i.e., HLA-identical brothers as determined by high-resolution molecular typing. One patient (CML 7) received peripheral blood mono-nuclear cells from a donor who was mobilized with G-CSF, 10 µg/kg, sc, for 5 days and who had to be submitted to two leukopheresis pro-cedures for the collection of mononuclear cells. The BM of the other donors was collected by multiple punctures of the posterior iliac bone under general anesthesia.
Conditioning regimen for the patients
All patients received a regimen of mye-loablative conditioning with busulfan, 4 mg kg-1 day-1, for 4 days (days -7 to -4) and
cyclophosphamide, 60 mg kg-1 day-1, for 2
days (days -3 and -2).
Prophylaxis against graft-versus-host disease
Prophylaxis against graft-versus-host
dis-ease was performed with cyclosporine and methotrexate (16). Cyclosporine was first administered intravenously, 3 mg kg-1 day-1,
by continuous infusion starting 24 h before BM or peripheral blood infusion. The dose was maintained until the occurrence of graft-ing and, as soon as the patient was able to swallow, cyclosporine was administered oral-ly at the dose of 7.5 mg kg-1 day-1 divided
into two daily portions and the dose was then adjusted so as to keep a serum level between 200 and 400 ng/mL. Methotrexate was ad-ministered intravenously at the dose of 15 mg/m2 on day +1 and of 10 mg/m2 on days
+3, +6, and +11. The dose of methotrexate was adjusted on the basis of the presence of severe mucositis and renal or hepatic insuf-ficiency according to the protocol of the institution.
Isolation, culture and differentiation of mesenchymal stem cells
BM samples from normal donors and from patients with CML were obtained from the iliac crest. The mononuclear cells were separated on a Ficoll-Paque™ Plus gradient (Amersham Biosciences, Uppsala, Sweden) and 2 x 107 mononuclear cells were cultured
Table 1. Characteristics of the patients, of the donors and of the allogeneic transplant in the present study.
Characteristics CML 6 CML 7 CML 8
Patient’s age/sex 24/Female 24/Female 29/Female
Diagnosis CML CML CML
Cytogenetics of patients 46,XX,t(9;22) 46,XX,t(9;22) 46,XX,t(9;22)
Phase of disease at BMT CP1 CP1 CP1
Conditioning regimen BU/CY2 BU/CY2 BU/CY2
GVHD prophylaxis CSA/MTX CSA/MTX CSA/MTX
Donor’s sex Male Male Male
Donor’s HLA type Related/identical Related/identical Related/identical
WBC count transplanted 3.6 2.4 3.7
(x 108 cells/kg)
BMT = bone marrow transplant; CML = chronic myeloid leukemia; CP1 = chronic phase 1; BU/CY2 = busulfan, 16 mg/kg, and cyclophosphamide, 120 mg/kg; GVHD = graft-versus-host disease; CSA/MTX = cyclosporine, 3 mg/kg iv, and methotrexate, 15 mg/m2 on day +1 and 10 mg/m2 on days +3, +6 and +11; HLA = human leukocyte
in α-MEM (Invitrogen, Carlsbad, CA, USA) containing 15% fetal calf serum (Invitrogen) supplemented with 2 mM L-glutamine, 100 IU/mL penicillin and 100 mg/mL strepto-mycin. After 3-7 days, non-adherent cells were removed and adherent cells were main-tained in culture and expanded with specific medium as previously described (5) after successive passages. After the third passage the cells were submitted to the adipogenic and osteogenic differentiation medium as previously described (17), and then sepa-rated for immunophenotyping, FISH and RT-PCR.
Isolation and purification of CD34+
hematopoietic stem cells
Mononucleated BM and peripheral blood cells from 7 patients with CML were separated by Ficoll-Hypaque centrifugation (Histo-paque®-1077, Sigma, St. Louis, MO, USA).
CD34+ cells were obtained by positive
selec-tion using an immunomagnetic column to col-lect CD34+ cells coupled to microbeads
ac-cording to the manufacturer’s protocol (MACS, Milteny Biotec, Bergisch Gladbach, Germany). After selection, CD34+ HSCs were
immedi-ately labeled with anti-human phycoerythrin-conjugated CD34 (Becton Dickinson, San Jose, CA, USA), anti-human FITC-conjugated CD14 (Becton Dickinson) or its isotype-matched control (anti-γ2a, Becton Dickinson)
in PBS for 30 min at 4ºC in the dark. After several washings, the cells were resuspended in 300 µL 2% BSA/PBS for FACS analysis. Flow cytometry of 75,000 events was per-formed and analyzed with a FACSort (Becton Dickinson) running CellQuest data acquisi-tion and analysis software (Becton Dickin-son). CD34+ HSC populations presenting ≥90% purity were used in the present study.
Mesenchymal stem cell separation and immunophenotyping
The culture-expanded MSCs were
imme-diately labeled with monoclonal antibodies specific for the following surface molecules: CD90-PE, CD73-PE, CD105-PE, CD51/61-FITC, CD29-PE, CD49a-PE, CD49e-PE, CD49d-PE, CD49b-PE, CD54-PE, CD106-PE, HLA-DR-FITC, HLA class I-FITC (Phar-mingen, San Jose, CA, USA); CD13-PE, CD14-PE, CD34-PE, CD44-FITC, CD45-FITC (Becton Dickinson); Ac133-PE (Milteny Biotec) or its isotype-matched control
(anti-γ2a, Becton Dickinson) in PBS for 30 min at
4ºC in the dark. After several washings, the cells were resuspended in 300 µL 2% BSA/ PBS for FACS analysis. For KDR (Sigma-Aldrich) and STRO-1 (kindly provided by Dr. Bervely Torok-Storb - Fred Hutchinson Can-cer Research Center, Seattle, WA, USA) stain-ing, MSCs were incubated with a primary antibody at 4ºC for 1 h. Cells were washed twice with 2% BSA/PBS and incubated with a secondary antibody (goat anti-mouse IgG1 (γ) FITC-conjugated antibody) for 1 h at 4ºC. After washing, cells were resuspended as men-tioned above for FACS analysis. Flow cytom-etry analysis of 10,000 events was performed using a FACSort (Becton Dickinson) running CellQuest data aquisition and analysis soft-ware (Becton Dickinson). Forward scatter and light scatter were used for gating on staining cells and excluding cell debris.
In order to characterize MSCs from CML patients after BMT and to exclude contami-nating hematopoietic cells we performed sorting with a FACS VANTAGE cytometer (Becton Dickinson). For this procedure, first culture-expanded MSCs were stained with anti-human CD90-FITC, CD54-PE, CD29-PE, CD13-CD29-PE, CD14-PE and CD33-CD29-PE, or PE or FITC-conjugated isotype control (BD Biosciences, San Jose, CA, USA). After stain-ing, cells were analyzed using the same cy-tometer apparatus. Sorting was then per-formed on the CD14-/CD33- cell
popula-tion. These CD14-/CD33- MSCs were
experiments the CD90+ cell purity was >96%
and the presence of CD14+/CD33+
contami-nant cells was <0.04%. These purified cell populations were then analyzed by nested PCR for the SRY gene.
Cytogenetic analysis
To identify the presence of the Ph1 chro-mosome in patients with a diagnostic hypo-thesis of CML and to evaluate the degree of post-transplant hematopoietic chimerism during the different treatment phases, BM and peripheral blood samples were collected into heparinized tubes and analyzed by stand-ard cytogenetic procedures. BM hematopoi-etic cells were processed by direct analysis, a method that does not require cell culture to obtain mitotic figures, with the metaphases obtained representing the in vivo situation of the cells at the time of sampling (18). Periph-eral cell metaphases were obtained by tem-porary lymphocyte culture (19). The chro-mosomes were identified by G banding (tryp-sin-Giemsa) and 20 metaphases per sample were analyzed using the 1995 ISCN nomen-clature (20).
Fluorescence in situ hybridization
The LSI BCR-ABL ES Dual Color probe (Vysis, Downer Grove, IL, USA) was used to identify the bcr-abl fusion gene in the 5 MSC samples and in 7 CD34+ HSC samples isolated
from patients with CML Ph+ at diagnosis. The
centromeric probes chromosome X α satellite (DXZ1)andchromosome Y α satellite(DXZ3) (Cytocell, Cambridge, UK) were used to quan-tify the chimerism of donor MSCs. The iso-lated MSC and HSC populations were treated with hypotonic solution (0.075 mM KCl) at 37ºC for 20 min, centrifuged at 200 g for 10 min and fixed with Carnoy solution (3:1 methanol:acetic acid; Merck, Darmstadt, Ger-many) for three cycles of 1-5 min followed by centrifugation at 200 g for 10 min. The nucleus suspension was fixed and placed on slides and
the preparations of best quality were selected by phase microscopy. The selected slides were pretreated with a 2X sodium chloride and sodium citrate solution, pH 7.0, at 37ºC for 10 min, dehydrated in an increasing ethanol se-ries (Merck) (70, 85, and 100%) for 5 min each and air dried. Nuclear DNA was dena-tured and hybridized according to the protocol provided by the manufacturer of the Vysis probes, followed by counterstaining with 10 µL of DAPI-II antifade solution (Vysis). In each case, at least 200 isolated nuclei were examined for the presence of the bcr-abl genes and about 600 nuclei were studied for XX/XY chimerism. The images were acquired with an AxioCam digital camera and analyzed with the AxioVision 3.0 software (Carl Zeiss, Oberkochen, Germany) using the Axioskop2 fluorescence microscope (Carl Zeiss) with the DAPI, FITC and rhodamine filters.
RT-PCR analysis of the bcr-abl gene in the
mesenchymal stem cells of patients with chronic myelogenous leukemia
Total MSC RNA was extracted with Trizol® LS (Gibco BRL®, Grand Island, NY,
using the 2OABL1 oligonucleotides 5' TGA TTA AGC CTA AGA CCC GGA 3' and LICABL2: 5'ACT GAA GCC GCT CGT TGG AAC TCC 3'. In these PCR assays, 2.5 µM of each oligonucleotide, 2 mM dNTPs and 1 unit of Taq polymerase were used in a final volume of 25 µL.
Analysis of donor chimerism by nested PCR
MSC DNA was extracted by digestion with proteinase K starting from 1 x 105 cells.
Briefly, the cell pellet was digested in 40 to 50 µL lysis buffer (100 mM NaCl, 10 mM Tris HCl, pH 8.0, 25 mM EDTA, pH 8.0, 0.5% SDS, and 0.1 mg/mL proteinase K) and incu-bated for 2 h at 55°C. After digestion, the enzyme was inactivated by incubation at 95ºC for 8 min. Approximately 2 to 5 µL of the digestion mixture was submitted to nested PCR. Two PCR cycles with two distinct sets of synthetic oligonucleotides were performed in this procedure. Both reactions consisted of 35 cycles of 30 s at 94°C, 40 s at a specific annealing temperature and 40 s at 72°C. After each cycle, a final 10-min extension at 72°C was performed. The oligonucleotides for the SRY gene were: a) for the first reaction 5'-GAATATTCCCGCTCTCCGGA-3' and 5' GTACAACCTGTTGTCCAGTTG-3' at an annealing temperature of 57°C, resulting in a PCR product of 416 bp, and b) for the second reaction 5'-CAGTGTGAAACGGGAGAA AACAGT-3' and 5'-GACGAGGTCGATACT TATAATTCG-3' at an annealing temperature of 60°C, resulting in a PCR product of 264 bp. As a control, the ß-globin gene was amplified by nested PCR using the primers 5'-CGTAAA TACACTTGCAAAGGAG-3' and 5'-CAG AGAGAGTCAGTGCCTATC-3', resulting in a 510-bp product after the first reaction, and the oligonucleotides 5'-GAAGAGCCAAGG ACAGGTAC-3' and 5'-CAACTTCATCCA CGTTCACC-3', resulting in a PCR product of 267 bp after the second reaction. For this gene, the annealing temperature was 57ºC in both reactions. In these PCR assays, 40 pmol of each oligonucleotide, 0.2 mM dNTPs and 2 Taq polymerase units were used in a final volume of 25 µL.
Results
Isolation, phenotypic analysis and potential for differentiation of the mesenchymal stem cells from the bone marrow of individuals with chronic myelogenous leukemia
The MSC populations of the BM from 5
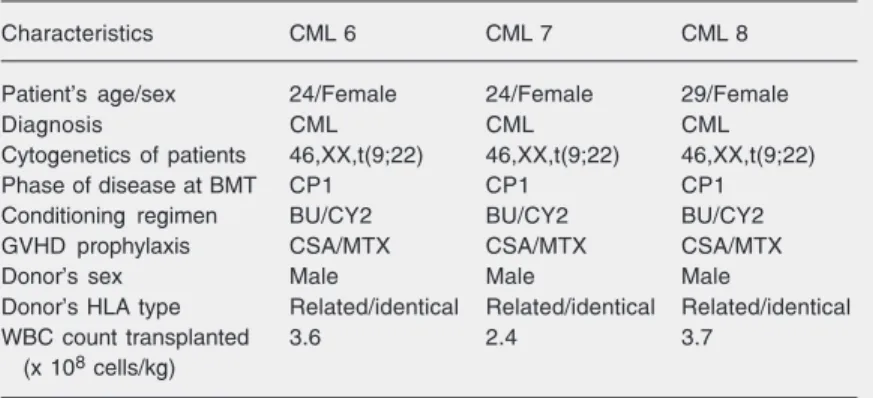
untreated patients in the chronic phase of CML, 3 female patients with CML who had received a BMT from a male donor, and 3 normal individuals were isolated, expanded and immunophenotyped. All samples per-mitted the isolation (based on the ability to adhere to plastic), culture and expansion of MSCs. The MSCs isolated from normal in-dividuals permitted the expansion of at least 20 passages, while the MSCs isolated from patients with CML demonstrated a consider-able reduction of their capacity for ex vivo expansion, which was limited between the 5th and 6th passages. In the 3rd passage, the MSCs obtained from CML patients at diag-nosis and after transplantation presented typi-cal fibroblastoid morphologitypi-cal characteris-tics with a capacity for differentiation simi-lar to that of normal MSCs. Figure 1 illus-trates the morphology of cultured MSCs from an untreated patient with CML (Figure 1A and B) and the morphology of these cultured cells without adipogenic (Figure 1C) and osteogenic medium (Figure 1E) or after differentiation into adipocytes and os-teocytes (Figure 1D and F).
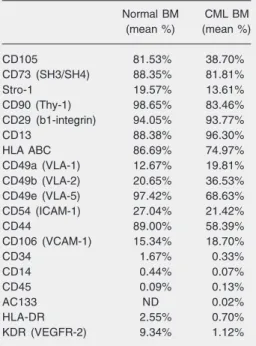
All MSC samples were submitted to im-munophenotyping between the second and fourth passage and were found to express CD105, CD73 (SH3/SH4), CD90, CD29, CD13, HLA ABC, CD49e, CD44, Stro-1, CD49a, CD49b, CD54, and CD106 antigens, but they did not express hematopoietic mark-ers (CD34, CD14, CD45, and CD133) or endothelial markers (KDR) and HLA-DR (Table 2). Our data show that in untreated CML patients the expression pattern for most of these MSC CD markers was similar to that found in normal samples. However, a slight difference was found for some surface anti-gens such as CD105, HLA ABC, CD49e, and CD44. The fact that MSCs comprise a het-erogeneous population containing sub-popu-lations at different stages of differentiation or small variations in antibody specificity when different lots are used can explain these differences in gene expression pattern.
BCR-ABL is not detected in mesenchymal stem cells isolated from the bone marrow of patients with chronic myelogenous leukemia
Primary cultures of adherent MSCs from the BM of 5 untreated patients with CML were evaluated in an attempt to identify the presence of the 9;22 translocation in these cells of the stromal microenvironment. Table 3 shows the results obtained by FISH for the MSC samples from the 5 untreated CML cases and from 1 control case. In two of these samples, CML 1 and CML 2, one BCR-ABL+ nucleus was detected among
approximately 180 nuclei analyzed. These results could not be confirmed by RT-PCR
Table 2. Immunophenotypic characteristics of mesenchymal stem cells obtained from the bone marrow (BM) of normal donors and untreated pa-tients with chronic myeloid leukemia (CML).
Normal BM CML BM
(mean %) (mean %)
CD105 81.53% 38.70%
CD73 (SH3/SH4) 88.35% 81.81%
Stro-1 19.57% 13.61%
CD90 (Thy-1) 98.65% 83.46%
CD29 (b1-integrin) 94.05% 93.77%
CD13 88.38% 96.30%
HLA ABC 86.69% 74.97%
CD49a (VLA-1) 12.67% 19.81%
CD49b (VLA-2) 20.65% 36.53%
CD49e (VLA-5) 97.42% 68.63%
CD54 (ICAM-1) 27.04% 21.42%
CD44 89.00% 58.39%
CD106 (VCAM-1) 15.34% 18.70%
CD34 1.67% 0.33%
CD14 0.44% 0.07%
CD45 0.09% 0.13%
AC133 ND 0.02%
HLA-DR 2.55% 0.70%
KDR (VEGFR-2) 9.34% 1.12%
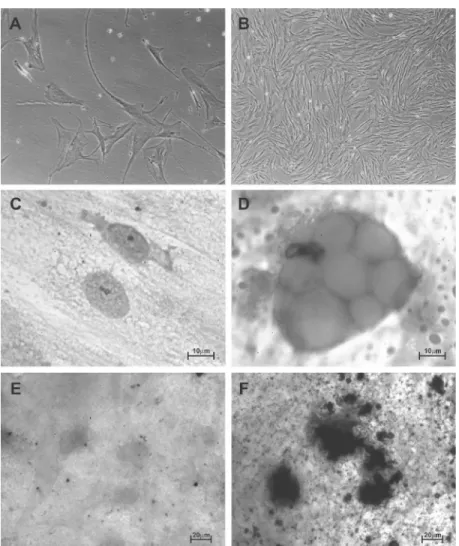
Figure 2. Fluorescence in situ hybridization (FISH) of the mes-enchymal stem cells (MSCs) and hematopoietic stem cells (HSCs) of patients with chronic myeloid leukemia (CML). A, I-FISH BCR-ABL using the LSI BCR-ABL ES Dual Color probe (Vysis) in the MSCs of the CML 1 patient showing two negative BCR-ABL cells (Ph-) and one
positive cell (Ph+) (1000X). B,
Analysis of the bcr-abl fusion gene by I-FISH in the CD34+
HSCs purified from the bone marrow (BM) of a patient with CML Ph1+, without treatment,
showing the presence of one normal nucleus (Ph-) and a
nucleus with t(9;22) (Ph+),
dem-onstrating the biclonal origin of CML in the HSCs (1000X). ES = extra signal. C, I-FISH using centromeric probes for the chro-mosome X α satellite (DXZ1) and Y α satellite (DXZ3; Cytocell) in the positive MSC XY isolated from a female patient after allogeneic transplantation of BM from a male donor (1000X), and D, in the positive MSC XX (1000X).
Table 3. Fluorescence in situ hybridization (FISH) analysis of the bcr-abl fusion gene in the mesenchymal stem cells isolated from the bone marrow of individuals with chronic myeloid leukemia (CML).
Case Passage No. No. of nuclei No. of BCR-ABL- No. of BCR-ABL+
(cytometry/FISH/RT-PCR) analyzed nuclei (%) nuclei (%)
CML 1 4º/4º/5º 195 194 (99.5%) 1 (0.5%)
CML 2 4º/5º/5º 174 173 (99.5%) 1 (0.5%)
CML 3 2º/4º/- 400 400 (100%) 0
CML 4 2º/3º/- 295 295 (100%) 0
CML 5 2º/3º/- 451 451 (100%) 0
Normal 2º/3º/- 211 211 (100%) 0
Table 4. Analysis of X/Y chimerism by nested polymerase chain reaction (PCR) and fluorescence in situ hybridization (FISH) X/Y after bone marrow transplantation (BMT).
Case Age Time BM Nested PCR SRY FISH X/Y
post-BMT (years) post-BMT karyotype MSCs
(days) Post-sorting (post-sorting)
MSCs (CD90+/ MSCs (CD90-/ MNC MNC
CD33-/CD14-) CD33+/CD14+) BM PB
BM BM
CML 6 24 551 94% 46,XY + + + + 0.3% XY (2/654)
CML 7 24 575 68% 46,XY + + ND + 0.1% XY (1/701)
CML 8 29 359 100% 46,XY - + ND +
in the two samples cited above. Curiously, the data for the CML 1 samples were not reproducible in the three reactions, with posi-tivity for the bcr-abl gene being diagnosed in only two of the three reactions performed. In contrast, in the CML 2 sample negativity was confirmed in all three reactions. The possibility of a false-positive result of FISH is very low since the probe used (LSI BCR-ABL ES Dual Color-Vysis) is characterized by giving origin to an extra signal coming from the ASS gene (synthetase argininosuc-cinate) located on the long arm of chromo-some 9 (9q), the derivative of t(9;22), in addition to the signal of bcr-abl fusion, thus increasing the reliability of our results. The fact that the second sample was negative in RT-PCR suggests the presence of a cell con-taminant of hematopoietic origin detected in the samples analyzed by FISH.
As a control, highly purified CD34+ HSCs
of BM and of peripheral blood from un-treated patients with CML were analyzed by FISH in order to quantify the presence of the fusion gene bcr-abl in this cell population (Figure 2). The present results show that the number of bcr-abl-positive CD34+ HSCs
ranged from 92 and 100% in BM and from 94 to 99% in purified peripheral blood samples. This analysis was also performed in mononuclear cell populations isolated from the BM and peripheral blood of a patient with CML, revealing that approximately 92% of the mononuclear cells of BM and 95% of those of peripheral blood were bcr-abl posi-tive (data not shown).
Absence of chimerism in the bone marrow of patients after bone marrow transplantation
XX/YY stromal chimerism was quanti-fied by FISH using centromeric probes for chromosomes X and Y (Figure 2). The anal-ysis was performed on 2 MSC samples that were found to be SRY positive by nested PCR. The percentage of XY MSCs was 0.3% (2/654) in one of the SRY+ sample (CML 6)
and 0.1% in theother (1/701; CML 7). All mononuclear cell samples from BM and pe-ripheral blood evaluated by nested PCR were SRY+, as also were the contaminating
mono-cytes (CD14+ cells) isolated from the MSC
population by flow cytometry (Table 4). As suggested in the analysis of the bcr-abl gene in the MSCs of CML patients, we believe that the small percentage of positive XY nuclei observed in the present study (3/1355 = 0.02%) was due to the presence of taminating hematopoietic cells. As a con-trol, we evaluated the mononuclear cells of the peripheral blood and BM of these pa-tients by nested PCR and confirmed the presence of chromosome Y in all samples (Table 4). To determine the kinetics of en-graftment of donor cells in the hematopoi-etic tissue, a retrospective chromosome anal-ysis of BM samples from the recipients was performed during different phases after BMT. The results indicated that the largest per-centage of hematopoietic metaphases ana-lyzed were from the donor (68 to 100% 46,XY) at the time of sample collection (Table 4).
Discussion
We determined the presence of the bcr-abl gene in the MSCs of BM from CML patients in order to determine the role of MSCs in the genesis of CML. We then in-vestigated whether this mutation might oc-cur in a more primitive progenitor cell popu-lation common to MSCs and HSCs, as ob-served for endothelial cells of patients with CML (9,10). To this end, we characterized phenotypically and cytogenetically a popu-lation of MSCs isolated from the BM of 5 Ph+-untreated CML patients. Analysis of the
sug-gests the presence of hematopoietic con-taminants in the culture of these cells, possi-bly (CD14+) bcr-abl-positive macrophages
that grow adhering to MSCs. Bhatia et al. (1) also described contaminating bcr-abl -posi-tive macrophages in in vitro cultures of stro-mal cells from the BM of CML patients and suggested that the abnormal function of the stroma in CML may be due to the presence of bcr-abl-positive macrophages detected in the marrow microenvironment that may con-tribute to the selective expansion of leuke-mic HSCs. The immunophenotypic analysis of MSCs from CML patients did not show any significant difference compared to MSCs isolated from normal individuals. Our re-sults agree with data reported by Zhao et al. (21) in an article that was published during the preparation of the present manuscript. The authors reported that MSCs in CML patients express the typical MSC CD mark-ers, but they did not show the percentage of positive cells for each surface antigen in comparison with normal BM-MSCs.
Several in vivo studies on animal models have demonstrated that MSCs can migrate and differentiate into various tissues after allo-geneic transplants (22-25). However, the transplantability of MSCs in humans is still controversial. Some studies investigating the degree of MSC chimerism in patients submit-ted to allogeneic BMT from donors of the opposite sex have demonstrated that some MSCs can be from the donor (13,14,26), whereas others did not report this finding (11, 12,15). In the present study, we observed the absence of transplantability of the MSCs isolated from 3 patients with CML submit-ted to allogeneic transplantation with a my-eloablative conditioning regimen. In these analyses we demonstrated the absence of donor MSCs in the stroma of the recipient, suggesting that the type of conditioning used does not seem to damage the stromal mi-croenvironment, a fact that did not provide the engraftment of donor MSCs during the study period (between +359 and +575 days).
Although 2 of the 3 patients analyzed pre-sented 0.1 and 0.3% microchimerism, we believe that the SRY+/XY+ cells respectively
detected by nested PCR and FISH were he-matopoietic contaminants which still per-sisted in small amounts in these samples. Thus, the present study supports previous investigations that did not demonstrate stro-mal microchimerism after allogeneic BMT. These results suggest that the type of stand-ard conditioning used (busulfan + cyclo-phosphamide) does not seem to influence the origin of MSCs after transplantation, although we cannot exclude the possibility of detecting stromal microchimerism in pa-tients submitted to other conditioning regi-mens during more recent post-BMT periods and/or in other hematologic diseases, as de-scribed by Villaron et al. (14, 27). These inves-tigators detected partial chimerism in the MSCs of the donor (26.13 and 60.17%) in 2 patients with multiple myeloma after BMT with a conditioning regimen of reduced intensity (flu-darabine + melphalan). However, it should be pointed out that multiple myeloma can dam-age the stromal microenvironment, possibly explaining the chimerism observed by Villaron et al. (14) in patients with the disease. Taken together, these data agree with a recent study by Polony et al. (28) which demonstrated the transplantability of MSC in 8 of 26 patients analyzed. None of them were CML patients and 2 of them were MM patients.
Therefore, we conclude that MSCs from CML patients express the typical MSC phe-notype, can be differentiated into osteogenic and adipogenic lineages and do not express the bcr-abl gene. However, the influence of MSCs on the dysregulation of hematopoie-sis in CML patients is worthy of further investigation. Moreover, MSCs from healthy donors cannot be found in recipients 12 to 20 months after allogeneic BMT.
Acknowledgments
par-References
1. Bhatia R, McGlave PB, Dewald GW, Blazar BR, Verfaillie CM. Abnormal function of the bone marrow microenvironment in chronic myelogenous leukemia: role of malignant stromal macrophages.
Blood 1995; 85: 3636-3645.
2. Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997; 276: 71-74.
3. Caplan AI, Bruder SP. Mesenchymal stem cells: building blocks for molecular medicine in the 21st century. Trends Mol Med 2001; 7: 259-264.
4. Minguell JJ, Erices A, Conget P. Mesenchymal stem cells. Exp Biol Med 2001; 226: 507-520.
5. Silva WA Jr, Covas DT, Panepucci RA, Proto-Siqueira R, Siufi JL, Zanette DL, et al. The profile of gene expression of human marrow mesenchymal stem cells. Stem Cells 2003; 21: 661-669.
6. Nowell PC, Hungerford DA. A minute chromosome in human chronic granulocytic leukemia. Science 1960; 132: 1497 (Abstract). 7. Kabarowski JH, Witte ON. Consequences of BCR-ABL expression
within the hematopoietic stem cell in chronic myeloid leukemia.
Stem Cells 2000; 18: 399-408.
8. Deininger MW, Goldman JM, Melo JV. The molecular biology of chronic myeloid leukemia. Blood 2000; 96: 3343-3356.
9. Gunsilius E, Duba HC, Petzer AL, Kahler CM, Grunewald K, Stockhammer G, et al. Evidence from a leukaemia model for mainte-nance of vascular endothelium by bone-marrow-derived endothelial cells. Lancet 2000; 355: 1688-1691.
10. Fang B, Zheng C, Liao L, Han Q, Sun Z, Jiang X, et al. Identification of human chronic myelogenous leukemia progenitor cells with he-mangioblastic characteristics. Blood 2005; 105: 2733-2740. 11. Awaya N, Rupert K, Bryant E, Torok-Storb B. Failure of adult
mar-row-derived stem cells to generate marrow stroma after successful hematopoietic stem cell transplantation. Exp Hematol 2002; 30: 937-942.
12. Stute N, Fehse B, Schroder J, Arps S, Adamietz P, Held KR, et al. Human mesenchymal stem cells are not of donor origin in patients with severe aplastic anemia who underwent sex-mismatched allo-geneic bone marrow transplant. J Hematother Stem Cell Res 2002; 11: 977-984.
13. Cilloni D, Carlo-Stella C, Falzetti F, Sammarelli G, Regazzi E, Colla S, et al. Limited engraftment capacity of bone marrow-derived mes-enchymal cells following T-cell-depleted hematopoietic stem cell transplantation. Blood 2000; 96: 3637-3643.
14. Villaron EM, Almeida J, Lopez-Holgado N, Alcoceba M, Sanchez-Abarca LI, Sanchez-Guijo FM, et al. Mesenchymal stem cells are present in peripheral blood and can engraft after allogeneic hemato-poietic stem cell transplantation. Haematologica 2004; 89: 1421-1427.
15. Rieger K, Marinets O, Fietz T, Korper S, Sommer D, Mucke C, et al. Mesenchymal stem cells remain of host origin even a long time after allogeneic peripheral blood stem cell or bone marrow
transplanta-tion. Exp Hematol 2005; 33: 605-611.
16. Storb R, Deeg HJ, Pepe M, Doney K, Appelbaum F, Beatty P, et al. Graft-versus-host disease prevention by methotrexate combined with cyclosporin compared to methotrexate alone in patients given marrow grafts for severe aplastic anaemia: long-term follow-up of a controlled trial. Br J Haematol 1989; 72: 567-572.
17. Covas DT, Siufi JL, Silva AR, Orellana MD. Isolation and culture of umbilical vein mesenchymal stem cells. Braz J Med Biol Res 2003; 36: 1179-1183.
18. Coutinho V, Bottura C. Método direto para o estudo dos cromosso-mos humanos. Rev Paul Med 1968; 72: 9-13.
19. Moorhead PS, Nowell PC, Mellman WJ, Battips DM, Hungerford DA. Chromosome preparations of leukocytes cultured from human peripheral blood. Exp Cell Res 1960; 20: 613-616.
20. Mitelman F. ISCN 1995: An international system for human
cytoge-netic nomenclature. Switzerland: S. Karger AG/Cytogenetics and
Cell Genetics; 1995.
21. Zhao Z, Tang X, You Y, Li W, Liu F, Zou P. Assessment of bone marrow mesenchymal stem cell biological characteristics and sup-port hematopoiesis function in patients with chronic myeloid leuke-mia. Leuk Res 2006; 30: 993-1003.
22. Hayakawa J, Migita M, Ueda T, Shimada T, Fukunaga Y. Genera-tion of a chimeric mouse reconstituted with green fluorescent pro-tein-positive bone marrow cells: a useful model for studying the behavior of bone marrow cells in regeneration in vivo. Int J Hematol 2003; 77: 456-462.
23. Devine SM, Cobbs C, Jennings M, Bartholomew A, Hoffman R. Mesenchymal stem cells distribute to a wide range of tissues follow-ing systemic infusion into nonhuman primates. Blood 2003; 101: 2999-3001.
24. Chapel A, Bertho JM, Bensidhoum M, Fouillard L, Young RG, Frick J, et al. Mesenchymal stem cells home to injured tissues when co-infused with hematopoietic cells to treat a radiation-induced multi-organ failure syndrome. J Gene Med 2003; 5: 1028-1038. 25. Mahmud N, Pang W, Cobbs C, Alur P, Borneman J, Dodds R, et al.
Studies of the route of administration and role of conditioning with radiation on unrelated allogeneic mismatched mesenchymal stem cell engraftment in a nonhuman primate model. Exp Hematol 2004; 32: 494-501.
26. Keating A, Singer JW, Killen PD, Striker GE, Salo AC, Sanders J, et al. Donor origin of the in vitro haematopoietic microenvironment after marrow transplantation in man. Nature 1982; 298: 280-283. 27. Villaron EM, Perez-Simon JA, San Miguel JF, del Canizo C. Bone
marrow mesenchymal stem cells chimerism after allogeneic he-matopoietic transplantation. Exp Hematol 2006; 34: 7.
28. Poloni A, Leoni P, Buscemi L, Balducci F, Pasquini R, Masia MC, et al. Engraftment capacity of mesenchymal cells following hematopoi-etic stem cell transplantation in patients receiving reduced-intensity conditioning regimen. Leukemia 2006; 20: 329-335.
ticipated in this study. We are also very grateful to Amélia Góes Araújo, Ane Rose