w w w. s b f g n o s i a . o r g . b r / r e v i s t a
Original
Article
Fucoidan
induces
G1
arrest
of
the
cell
cycle
in
EJ
human
bladder
cancer
cells
through
down-regulation
of
pRB
phosphorylation
Hye
Young
Park
a,1,
Il-Whan
Choi
b,1,
Gi-Young
Kim
c,
Byung
Woo
Kim
d,e,
Wun-Jae
Kim
f,
Yung
Hyun
Choi
a,e,∗aDepartmentofBiochemistry,DongeuiUniversityCollegeofKoreanMedicine,Busan614-052,RepublicofKorea bDepartmentofMicrobiology,CollegeofMedicine,InjeUniversity,Busan608-756,RepublicofKorea
cLaboratoryofImmunobiology,DepartmentofMarineLifeSciences,JejuNationalUniversity,Jeju690-756,RepublicofKorea
dDepartmentofLifeScienceandBiotechnology,CollegeofNaturalSciences&HumanEcology,DongeuiUniversity,Busan614-714,RepublicofKorea eBlue-BioIndustryRICandAnti-AgingResearchCenter,DongeuiUniversity,Busan614-714,RepublicofKorea
fDepartmentofUrology,CollegeofMedicine,ChungbukNationalUniversity,Cheongju362-763,RepublicofKorea
a
r
t
i
c
l
e
i
n
f
o
Articlehistory:
Received30December2014 Accepted22March2015 Availableonline25April2015
Keywords:
Fucoidan EJcells G1arrest Apoptosis pRB
a
b
s
t
r
a
c
t
Fucoidan,asulfatedpolysaccharidefoundinmarinealgaeandbrownseaweeds,hasbeenshowntoinhibit theinvitrogrowthofhumancancercells.ThisstudywasconductedinculturedhumanbladdercancerEJ cellstoelucidatethepossiblemechanismsbywhichfucoidanexertsitsanti-proliferativeactivity,which untilnowhasremainedpoorlyunderstood.FucoidantreatmentofEJcellsresultedindose-dependent inhibitionofcellgrowthandinducedapoptoticcelldeath.Flowcytometricanalysisrevealedthatfucoidan ledtoG1arrestincellcycleprogression.Itwasassociatedwithdown-regulationofcyclinD1,cyclinE, andcyclin-dependent-kinases(Cdks)inaconcentration-dependentmanner,withoutanychangeinCdk inhibitors,suchasp21andp27.Furthermore,dephosphorylationofretinoblastomaprotein(pRB)bythis compoundwasassociatedwithenhancedbindingofpRBwiththetranscriptionfactorsE2F-1andE2F-4. Overall,ourresultsdemonstratethatfucoidanpossessesanticanceractivitypotentialagainstbladder cancercellsbyinhibitingpRBphosphorylation.
©2015SociedadeBrasileiradeFarmacognosia.PublishedbyElsevierEditoraLtda.Allrightsreserved.
Introduction
Bladdercanceris anyofseveraltypes ofmalignancy arising fromtheepithelial lining ofthe urinary bladder.In theUnited States,bladdercanceristhefourthmostcommontypeofcancer inmenandtheninthmostcommoncancerinwomen,although theincidenceofbladdercancerinAsiaismuchlower(Kakehietal., 2010;Abdollahetal.,2013).Despiterecentadvancesinsurgicaland chemotherapeuticprocedures,long-termsurvivalratesarepoor, andthemostcommoncauseofmortalityisrecurrencewith metas-tasis(Racioppietal.,2012).Therefore,itisimportanttodevelop othereffectivestrategiestoimprovethesurvivalrateforbladder cancerpatients.
Several epidemiological studies have revealed that dietary intakeofmarinealgaeand seaweedsareprotective againstthe riskofvarioustypesofmalignancies(Kimetal.,2011;Parkand
∗ Correspondingauthor.
E-mail:[email protected](Y.H.Choi). 1Theseauthorscontributedequallytothiswork.
Pezzuto,2013;Ahmedetal.,2014).Brownseaweedsareusedas an importanthealthcare foodand a pharmaceutical product in Asiancountries.Fucoidanisamajorsulfatedpolysaccharidefound inbrownseaweed;ithasbeenwellcharacterizedandisknown tohave variousbiologicalfunctions,includingantioxidant, anti-inflammatory,andanticancereffects(Lietal.,2008;Sennietal., 2011;Fitton,2011;Wangetal.,2012;Senthilkumaretal.,2013; ThomasandKim,2014).Wereportedrecentlythatfucoidanmay offer substantial therapeutic potential for treatment of inflam-matoryandneurodegenerativediseasesthatareaccompaniedby microglial activation (Park et al., 2011a). In addition, fucoidan suppresses cancercell proliferation and inhibitsthe growth of transplantedtumorxenograftsbyinducing apoptosisand/orby blockingabnormalcellcycleprogressionattheG1orG2/Mphase (Riouetal.,1996;Fukahorietal.,2008;Parketal.,2011b,2013; Zhangetal.,2011,2013;Hsuetal.,2013;Xueetal.,2013;Yang etal.,2013;Parketal.,2014;Banafaetal.,2013;Chenetal.,2014; SenthilkumarandKim,2014;Choetal.,2014).However,thiseffect isselectiveforcancercells,asnormalcelllinesareresistanttocell cyclearrestand apoptosisbyfucoidan.Moreover,fucoidanalso inhibitsmigrationandinvasionofhighlymetastaticcancercells
http://dx.doi.org/10.1016/j.bjp.2015.03.011
viadown-regulationofmatrixmetalloproteinasesandinhibitionof thephosphoinositide3-kinase/Aktandnuclearfactor-kBsignaling pathways(Saitohetal.,2009;Leeetal.,2012;Wangetal.,2014; SenthilkumarandKim,2014).However,themolecularmechanisms ofitsanti-proliferativeactionsonhumanbladdercarcinomacell growthhave not yetbeen examined.Thus, thepurposeof this studywastoinvestigatetheeffectsoffucoidanoncell prolifera-tionofthehumanbladdercarcinomacelllineEJandtoexplorethe potentialmechanismsoftheeffects.Ourdataindicatethatfucoidan inhibitedthegrowthofEJcellsinaconcentration-dependent man-ner,arrestingthemintheG1phaseoftheircellcycleandinducing apoptosis.
Materialsandmethods
Cellculture
EJcells werepurchased fromAmericanTypeCulture Collec-tion (ATCC, Manassas, VA, USA) and maintained in RPMI-1640 medium(GIBCO-BRL,Gaithersburg,MD)supplementedwith10% fetal bovine serum (FBS) and 2mM of l-glutamine and
peni-cillin/streptomycin.Thecellswereculturedinanincubatorwith 5%CO2at37◦C.FucoidanwaspurchasedfromSigma-Aldrich(St.
Louis,MO,USA)anddissolvedinphosphate-bufferedsaline(PBS) asastocksolutionata200mg/mlconcentration,andthestock solu-tionwasthendilutedwiththemediumtothedesiredconcentration priortouse.
MTTassay
Measurementofcellviabilitywasdeterminedusingthe 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Sigma-Aldrich)assay,whichisbasedontheconversionofMTTto MTT-formazanbymitochondria.Inbrief,cells(2×104cells/well)
wereseededin24-wellplatesandexposedtofucoidanfor48h. Aftertreatment,5mg/mlMTTsolutionwasadded,followedby3h incubationat37◦Cinthedark,andthemediawasthenremoved.
The formazan precipitate was dissolved in dimethyl sulfoxide (Sigma-Aldrich), and absorbance of the formazan product was measured at a wavelength of 540nm with an enzyme-linked immunosorbentassay(ELISA)reader(MolecularDevices, Sunny-vale,CA,USA)(Leeetal.,2014).Forthemorphologicalstudy,cells werephotographeddirectlyusing aninverted microscope(Carl Zeiss,Oberkochen,Germany).
Flowcytometryanalysis
Exponentiallygrowingcellswerecomparedtocellstreatedwith variousconcentrationsoffucoidanfor48h.Aftertreatmentwith fucoidan,thecellswerecollectedandfixedin70%ethanolat4◦C
for30min,and theDNAcontentofcells wasstainedwith pro-pidiumiodide(PI)usingaDNAstainingkit(CycleTESTPLUSKit, BectonDickinson,SanJose,CA,USA)inaccordancewiththe manu-facturer’sinstructions.ThecellswerethensubjectedtoaFACScan flowcytometer(BectonDickinson)withthepercentagesofcellsin differentphasesofthecellcyclecalculatedfromDNAhistograms. Cellswithsub-G1DNAcontentwereconsideredapoptotic cells (Kimetal.,2014).
Morphologicalobservationofnuclearchange
Afterculturewithvariousconcentrationsoffucoidan,cellswere washed twicewith PBS and fixedwith 3.7% paraformaldehyde (Sigma-Aldrich)inPBSfor10minatroomtemperature.Thefixed cellswerewashedtwicewithPBSandstainedwith 4,6-diamidino-2-phenylindole(DAPI,Sigma-Aldrich)solutionfor10minatroom
Table1
OligonucleotidesusedinRT-PCR.
Genename Sequence
CyclinD1 Sense 5′-TGG-ATG-CTG-GAG-GTC-TGC-GAG-GAA-3′
Antisense 5′-GGC-TTC-GAT-CTG-CTC-CTG-GCA-GGC-3′
CyclinE Sense 5′-AGT-TCT-CGG-CTC-GCT-CCA-GGA-AGA-3′
Antisense 5′-TCT-TGT-GTC-GCC-ATA-TAC-CGG-TCA-3′
Cdk2 Sense 5
′-GCTTTCTGCCATTCTCATCG-3′
Antisense 5′-GTCCCCAGAGTCCGAAAGAT-3′
Cdk4 Sense 5
′-ACG-GGT-GTA-AGT-GCC-ATC-TG-3′
Antisense 5′-TGG-TGT-CGG-TGC-CTA-TGG-GA-3′
Cdk6 Sense 5
′-CGA-ATG-CGT-GGC-GGA-GAT-C-3′
Antisense 5′-CCA-CTG-AGG-TTA-GAG-CCA-TC-3′
p21 Sense 5′-CTCAGAGGAGGCGCCATG-3′
Antisense 5′-GGGCGGATTAGGGCTTCC-3′
p27 Sense 5′-AAG-CAC-TGC-CGG-GAT-ATG-GA-3′
Antisense 5′-AAC-CCA-GCC-TGA-TTG-TCT-GAC-3′
GAPDH Sense 5
′-CGG-AGT-CAA-CGG-ATT-TGG-TCG-TAT-3′
Antisense 5′-AGC-CTT-CTC-CAT-GGT-GGT-GAA-GAC-3′
temperature.ThecellswerewashedtwomoretimeswithPBS. Cov-erslipsweremountedonglassslidesandanalyzedbyfluorescence microscopyusingaZeissAxiophotmicroscope(CarlZeiss).
Annexin-Vstaining
Toanalyzeapoptosis,thecellsweretreatedwiththeindicated concentrationsoffucoidanfor48handresuspendedinan annexin-Vbindingbuffercontaining10mMHEPES/NaOH(pH7.4),140mM NaCl,and2.5mMCaCl2.Aliquotsofthecellswereincubatedwith annexin-V fluorescein isothiocyanate(FITC, R&D Systems, Min-neapolis, MN, USA), mixed, and incubated for 15min at room temperatureinthedark.PIataconcentrationof5g/mlwasadded todistinguishnecrotic cells.Theapoptoticcells (AnnexinV+/PI− cells)weremeasuredbyafluorescence-activatedcellsorter analy-sisinaFACScanflowcytometer(ParkandHan,2014).
RNAextractionandreversetranscription-polymerasechain reaction(PCR)
TotalRNAwasisolatedusinganRNeasyminikit(Qiagen,LaJolla, CA,USA)andprimedwithrandomhexamerstosynthesize comple-mentaryDNAusingAMVreversetranscpriptase(AmershamCorp., ArlingtonHeights,IL,USA)followingthemanufacturer’s instruc-tions.PCRwasperformedinaMastercycler(Eppendorf,Hamburg, Germany)withtheindicatedprimers,whichwerepurchasedfrom Bioneer(Seoul,RepublicofKorea),inTable1.Theconditionsforthe PCRreactionswere1×(94◦Cfor3min),35X(94◦Cfor45s;58◦Cfor
45s;and72◦Cfor1min),and1×(72◦Cfor10min).The
amplifica-tionproductsobtainedbyPCRwereelectrophoreticallyseparated ona1.0%agarosegelandvisualizedbyethidiumbromide(EtBr, Sigma-Aldrich)staining(Gongetal.,2014).In aparallel experi-ment,glyceraldehyde-3-phosphatedehydrogenase(GAPDH)was usedasaninternalcontrol.
Totalproteinextraction,immunoprecipitation,andWesternblot analysis
For isolation of total protein fractions, the cells were har-vested and washedoncewithice-cold PBS,and lysed withcell lysis buffer[20mMTris pH7.5,150mM NaCl,1%Triton X-100, 2.5mM sodiumpyrophosphate, 1mM EDTA,0.5g/mlleupeptin, 1% Na3CO4, 1mM phenylmethane-sulfonyl fluoride], and
120
A
B
100
80
60
Cell viability (%)
Cell numbers
40
20
0
0 50
Fucoidan (µg/ml)
DNA contents
G1 G2/M
47.4 79.4
17.1 8.5
91.1 92.7% of G1
% of G2/M % of S
2.8 4.5 3.0
5.9 29.8
22.8
S
Fucoidan (µg/ml)
100 150
0 50 100 150
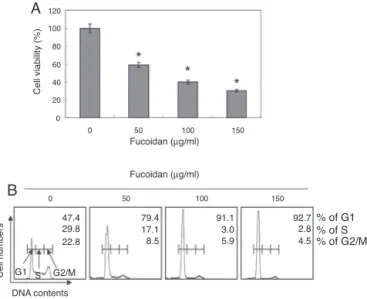
Fig.1. InhibitionofcellviabilityandinductionofG1arrestbyfucoidantreatment inhumanbladdercancerEJcells.(A)Cellsweretreatedwiththeindicated concen-trationsoffucoidanfor48h.Thecellswereharvested,andthepercentageofcell viabilitywascalculatedwithanMTTassay.Dataarepresentedasmean±SDin trip-licate.SignificancewasdeterminedbytheStudent’st-test(*p<0.05vs.untreated control).(B)Thecellswerecollected,fixed,andstainedwithPIforflowcytometry analysis.DNAcontentisrepresentedonthex-axis,andthenumberofcellscounted isrepresentedonthey-axis.Eachpointrepresentsthemeanoftwoindependent experiments.
assay (Bio-Rad Lab., Hercules, CA, USA), following the proce-duredescribedbythemanufacturer.Forimmunoprecipitation,cell extractswereincubatedwithimmunoprecipitatingantibodiesin extractionbufferfor1hat4◦C,andthentheimmunocomplexes
wereprecipitatedwithproteinA-Sepharosebeads(Sigma-Aldrich). ForWesternblotassay,thetotalproteinsandimmunoprecipitated proteinswere subjected to electrophoresis on sodium dodecyl sulphate(SDS)–polyacrylamide gelsandtransferred to nitrocel-lulose membranes (Schleicher & Schuell, Keene, NH, USA) by electroblotting.Blotsweresubsequentlyblockedwith5%non-fat milkandprobedwiththedesiredantibodiesfor1hat4◦C,
incu-bated withdiluted enzyme-linked secondary antibodyat room temperaturefor afurther1h, and thenvisualizedbyenhanced chemiluminescence(ECL,Amersham)accordingtorecommended procedures.Eextracellular-regulatedkinase(ERK)wasusedasan internal control. The primary antibodies were purchased from SantaCruzBiotechnologyInc.(SantaCruz,CA,USA). Peroxidase-labeleddonkeyantirabbitimmunoglobulinandperoxidase-labeled sheepantimouse immunoglobulin were purchased from Amer-sham.
Statisticalanalysis
Thedatawereexpressedasmeans±SD fortriplicate experi-ments.StatisticalanalyseswereperformedusingaStudent’st-test. Apvalue<0.05wasconsideredstatisticallysignificant.
Results
FucoidaninhibitscellviabilityandinducesG1arrestinEJcells
Todeterminewhetherfucoidaninfluencedthecellviabilityof theEJcells,thecellswereculturedinthepresenceoffucoidanfor 48h,andthecellviabilitywasdeterminedwithanMTTassay.As showninFig.1A,fucoidanreducedtheviabilityoftheEJcellsin aconcentration-dependentmanner.Thecellcyclephase distribu-tionofexponentiallygrowingEJcellswasnextcomparedtocells
Fucoidan (µg/ml)
Fucoidan (µg/ml)
Apoptotic cells (%)
0
A
B
C
50 100 150
0 0 5 10 15 20 25 30
50 100 150
Fig.2. InductionofapoptosisbyfucoidantreatmentinEJcells.Thecellsweretreated withvariousconcentrationsoffucoidanfor48h.(A)Themorphologicalchangesof thecellswereimagedusinganinvertedmicroscope(originalmagnification,200×). (B)ThecellswerefixedandstainedwithDAPIsolution.After10minincubationat roomtemperature,thestainednucleiwereobservedwithafluorescentmicroscope (originalmagnification,400×).(C)Thecellswereharvestedfordeterminingthe per-centageofannexin-Vpositive/PInegative(apoptoticcells).Thedataareexpressedas themean±SDofthreeindependentexperiments.Thesignificancewasdetermined bytheStudent’st-test(*p<0.05vs.untreatedcontrol).
treatedwithfucoidan.Comparedwiththeuntreatedcontrol,the fucoidan-treatedcellsaccumulatedintheG1phaseofthecellcycle inaconcentration-dependentmanner(Fig.1B),andtherewasa decreaseinthenumberofcellsintheSandG2/Mphases.Taken together,theseresultssuggestthatthegrowthinhibitoryeffectof fucoidanonEJcellsistheresultofitblockingtheG1phase.
FucoidaninducesapoptosisinEJcells
Asfucoidaninducedmorphologicalchanges,suchasmembrane blebbinganda reductionincellvolumeunder thesame condi-tions(Fig.2A),weassessed theeffectoffucoidantreatmenton theapoptoticcharacteristicsoftheEJcellstodeterminewhether apoptosiswasinvolvedinfucoidan-mediatedantiproliferation[0].
Fig.2Brevealsthatnucleicmorphologicalchangesofapoptosisas indicatedbyrepresentativeimagesofcondensedandfragmented nucleiwereclearlyevidentinfucoidan-treatedEJcellsincontrast totheuntreatedcells.Toquantifyfucoidan-inducedapoptosis,the EJcellstreatedwithfucoidanwerestainedwithannexinV-FITCand analyzedbyflowcytometry.AsshowninFig.2C,fucoidanincreased theannexin-V-FITCstainedapoptoticpopulationofcells,andthese effectsweredosedependent.Theseobservationsindicatethatthe inhibitionofcellviabilityobservedinresponsetofucoidanisalso associatedwiththeinductionofapoptosis.
Effectoffucoidanontheexpressionofcellcyclecheckpoint regulators
0
1 0.6 0.5 0.4
1 0.9 0.9 0.8
1 0.9 0.9 1
1 0.9 0.8 0.5
1 0.8 0.5 0.4
1 0.9 0.8 0.7
1 0.8 0.7 0.4
1 0.7 0.6 0.3
1 1 1 1
1 1 1 1
50
Fucoidan (µg/ml)
A
B
Cycline D1
100 150 0 50
Fucoidan (µg/ml)
100 150
Cycline E
Cdk2
Cdk4
Cdk6
GAPDH
Cycline D1
Cycline E
Cdk2
Cdk4
Cdk6
ERK
Fig.3.Down-regulationofG1-associatedcyclinsandCdksbyfucoidantreatmentinEJcells.(A)Aftertreatmentwithvariousconcentrationsoffucoidanfor48h,thecells werecollected,andtotalRNAwasisolatedandreverse-transcribed.TheresultingcDNAswerethensubjectedtoPCRwiththeindicatedprimers,andthereactionproducts wereseparatedin1.0%agarosegelandvisualizedbyEtBrstaining.GAPDHwasusedasaninternalcontrol.(B)Thecelllysateswereseparatedandequalamountsoftotal celllysatesweresubjectedtoSDS–polyacrylamidegels,transferred,andprobedwiththeindicatedantibodies.ERKwasusedasaninternalcontrol.Therelativeratiosof expressionintheresultsoftheWesternblottingwerepresentedatthebottomofeachoftheresultsasrelativevaluesoftheGAPDHandERKexpression,respectively.
andthenRT-PCRandWesternblotanalysiswereperformed.As illustratedinFig.3,fucoidansuppressedtheexpressionof biomark-ers,suchascyclinD1,cyclinE,cyclin-dependent-kinase(Cdk)2, Cdk4,andCdk6,in thetransitionoftheG1totheSphasein a concentration-dependentmanner.However,fucoidandidnot sig-nificantlyaffectthelevelsofCdkinhibitors,suchasp21andp27 (Fig.4).
0
1 0.9 1 1
1 0.9 1 1
50 Fucoidan (µg/ml)
A
p21
p27
100 150
GAPDH
0
1 0.9 0.5 0.3
1 0.9 1 1.1
50 Fucoidan (µg/ml)
B
p21
p27
100 150
ERK
Fig.4. EffectsoffucoidanonthelevelsofCdkinhibitorsinEJcells.(A)Thecells weretreatedwithdifferentconcentrationsoffucoidanfor48h.TotalRNAwas iso-latedandreverse-transcribed,andtheresultingcDNAsweresubjectedtoPCR.The reactionproductsweresubjectedtoelectrophoresisin1.0%agarosegeland visual-izedbyEtBrstaining.GAPDHwasusedasaninternalcontrol.(B)Afterincubation withfucoidanunderthesameconditions,thecelllysateswereseparated,andequal amountsoftotalcelllysatesweresubjectedtoSDS–polyacrylamidegels,transferred, andprobedwithantibodiesagainstp21andp27.ERKwasusedasaninternal con-trol.TherelativeratiosofexpressionintheresultsoftheWesternblottingwere presentedatthebottomofeachoftheresultsasrelativevaluesoftheGAPDHand ERKexpression,respectively.
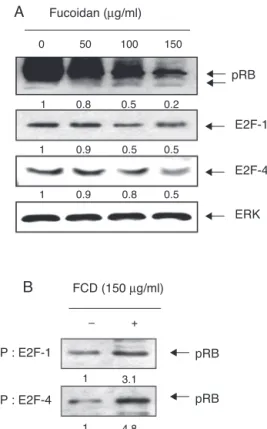
Fucoidandown-regulatespRBphosphorylationandincreasesthe bindingofpRBandE2Fs
As D-type cyclins and cyclin E-stimulatedCdk activity con-verges in hyperphosphorylation of the retinoblastoma protein (pRB), the effect of fucoidan on the phosphorylation status of pRBwasinvestigatedbyWesternblotanalysis.Fucoidancauseda remarkabledecreaseintotallevelsofpRBexpressionandchanged fromahyperphosphorylatedformtoahypophosphorylatedform (Fig.5A).Furthermore,co-immunoprecipitationanalysisindicated thatthereappearedtobenoassociationbetweenpRBandE2Fs, suchasE2F-1andE2F-4,intheuntreatedcontrolcells[0]. How-ever,therewasastrongincreaseinthebindingofpRBandE2Fs inthefucoidan-treatedEJcells(Fig.5B),suggestingthatfucoidan inhibitsthereleaseofE2FsproteinfrompRB.
Discussion
Although fucoidan exerts various pharmacological activities, such as anti-inflammatory, antioxidant, and anticancer effects (Riouetal.,1996;Lietal.,2008;Fukahorietal.,2008;Saitohetal., 2009;Fitton,2011;Sennietal.,2011;Parketal.,2011b,2013;Zhang etal.,2011,2013;Wangetal.,2012;Leeetal.,2012;Liuetal.,2012; Hsuetal.,2013;Xueetal.,2013;Yangetal.,2013;Senthilkumar etal.,2013;Parketal.,2014;Banafaetal.,2013;Chenetal.,2014; SenthilkumarandKim,2014;Wangetal.,2014),theanti-cancer activityoffucoidaninhumanbladdercancercellshasrarelybeen reported.Therefore,weelucidatedtheabilityoffucoidantoinhibit thegrowthofhumanbladdercancerEJcells.Inthisstudy,wefound thatfucoidanshowsconsiderablepotentialtoinhibitthe prolifer-ationofEJcellsviacellcyclearrestintheG1phaseandapoptosis induction.
0
1 0.8 0.5 0.2
1 0.9 0.5 0.5
1
1 3.1
– +
1 4.8
0.9 0.8 0.5
50 Fucoidan (µg/ml)
A
pRB
E2F-1
100 150
E2F-4
ERK
FCD (150 µg/ml)
IP : E2F-1
IP : E2F-4
B
pRB
pRB
Fig.5. InductionofhypophosphorylationofpRBandenhancedassociationofpRB andE2FsbyfucoidantreatmentinEJcells.(A)Aftertreatmentwithvarious concen-trationsoffucoidanfor48h,thetotalcelllysateswerepreparedandseparatedby electrophoresisonan8%or10%SDS–polyacrylamidegel.Westernblottingwasthen performedusinganti-pRB,anti-E2F-1,andanti-E2F-4antibodies.ERKwasusedasan internalcontrol.(B)Thecellswereincubatedwithorwithout150g/mloffucoidan for48h,andthenequalamountsofproteins(0.5mgofprotein)were immunoprecip-itatedwithanti-E2F-1oranti-E2F-4antibody.Immunocomplexeswereseparated by8%SDS–polyacrylamidegelelectrophoresis,transferredtoanitrocellulose mem-brane,andprobedwithanti-pRBantibody.ProteinsweredetectedbyECLdetection. TherelativeratiosofexpressionintheresultsoftheWesternblottingwere pre-sentedatthebottomofeachoftheresultsasrelativevaluesoftheERKexpression oruntreatedcontrol,respectively.
inhibitors,includingp21andp27(Dobashietal.,2003;Paternot etal.,2010).Inthepresent study,theresultsfromRT-PCR and immunoblottinganalysisclearlydemonstratedthatthelevelsof cyclinD1 and cyclin E expression weremarkedly inhibited by fucoidantreatmentatbothtranscriptionalandtranslational lev-els(Fig.3).Inaddition,fucoidan-inducedG1arrestwascorrelated withthedown-regulationofCdks,suchasCdk2,Cdk4,andCdk6. Although,arecentstudyindicatedthatfucoidandown-regulated cyclinE,Cdk2,Cdk4resultinginG0/G1arrestthroughbindingof Cdkinhibitorp21toCdk2andCdk4incellstreatedwithfucoidanin humanbladdercancer5637cells(Choetal.,2014),theexpression ofCdkinhibitorssuchasp21andp27wasremainedunchangedin fucoidan-treatedEJcells(Fig.4).
Ontheotherhand,pRBasatumorsuppressorisalso impor-tantforcellcycleprogressionduringtheG1toSphasetransition. DephosphorylationofpRBinhibitscellcycleprogressionby inter-acting with transcription factors of the E2F family, whereas phosphorylationofpRBresultsininductionofcellcycle progres-sionthroughthebreakingofpRB/E2Fcomplexes(Dobashietal., 2003;Paternotetal.,2010).Therefore,ifthelevelsofeitherprotein aredecreasedortheassociationbetweentheirrespectivebindings partnersisdiminished,aconcomitantdecreaseinthedegreeofpRB phosphorylationwouldbeexpected.In conjunctionwith down-regulationoftotallevelsofpRB,thedatashowedthattreatment withfucoidaninhibitedthephosphorylationofpRBandenhanced theassociationofpRBandE2Fs(Fig.5).TheroleofCdkinhibitors
hasattractedattentionintheinductionofG1phasearrest,which isrelatedtotherestrictionoftheproliferationofcancercellsvia fucoidantreatment.However,accordingtoBooetal.(2012), restric-tionofE2FexpressionplaysanimportantroleininducingG1cell cyclearrestinPC-3prostaticcancercells,alongwiththeincreased expressionof Cdkinhibitor p21.Moreover,a recentstudy con-ductedinourlabrevealedthatphosphorylationinhibitionofpRB wassimultaneouslyinvolvedinadditiontotheexpressionincrease ofCdkinhibitorp21inT24bladdercancercells’G1arrestbrought aboutbyfucoidan(Parketal.,2014).Hence,evenifcancercells’G1 arrestresultingfromfucoidantreatmentdiffersaccordingtotypes ofcancercells,itcanatleastbesaidthatexpressionincreaseof Cdkinhibitorssuchasp21andphosphorylationinhibitionplayan importantfactorsinG1arrest.
Conclusions
In conclusion, the resultspresented here demonstrate that: (i) reduced survival of EJ cells after exposure to fucoidan is associatedwithG1phase cellcyclearrestand apoptosis induc-tion;(ii)fucoidaninhibitscellcycleprogressionintheG1phase bydecreasingG1-related cyclins and Cdks;(iii) treatmentwith fucoidanresultsinincreasedbindingofpRBtoE2Fs.Thesenovel phenomenahavenotbeenpreviouslydescribedinbladdercancer cellsandsuggestthatfucoidanandrelatedcompoundsmayhave significantpotentialinthedevelopmentofcancertreatments.
Authors’contributions
HYP and IWC contributed in running the laboratory work, analysisofthedataand draftedthepaper.GYK, BWKandWJK contributedtocritical readingof themanuscript.WJKand YHC designed the study, supervised the laboratory work and con-tributedtocriticalreadingofthemanuscript.Alltheauthorshave readthefinalmanuscriptandapprovedthesubmission.
Conflictsofinterest
Theauthorsdeclarenoconflictsofinterest.
Acknowledgments
ThisworkwassupportedbyBlue-BioIndustryRegional Innova-tionCenter(RIC08-06-07)atDongeuiUniversityasaRICprogram underMinistryofTrade,Industry&EnergyandBusancityandthe NationalResearchFoundationofKorea(NRF)grantfundedbythe Koreagovernment(MSIP)(No.NRF-2014R1A2A1A09006983).
References
Abdollah,F.,Gandaglia,G.,Thuret,R.,Schmitges,J.,Tian,Z.,Jeldres,C.,Passoni,N.M., Briganti,A.,Shariat,S.F.,Perrotte,P.,Montorsi,F.,Karakiewicz,P.I.,Sun,M.,2013. Incidence,survivalandmortalityratesofstage-specificbladdercancerinUnited States:atrendanalysis.CancerEpidemiol.37,219–225.
Ahmed,A.B.,Adel,M.,Karimi,P.,Peidayesh,M.,2014.Pharmaceutical, cosmeceuti-cal,andtraditionalapplicationsofmarinecarbohydrates.Adv.FoodNutr.Res. 73,197–220.
Banafa,A.M.,Roshan,S.,Liu,Y.Y.,Chen,H.J.,Chen,M.J.,Yang,G.X.,He,G.Y.,2013. FucoidaninducesG1phasearrestandapoptosisthroughcaspases-dependent pathwayandROSinductioninhumanbreastcancerMCF-7cells.J.Huazhong Univ.Sci.Technol.Med.Sci.33,717–724.
Boo,H.J.,Hong,J.Y.,Kim,S.C.,Kang,J.I.,Kim,M.K.,Kim,E.J.,Hyun,J.W.,Koh,Y.S., Yoo,E.S.,Kwon,J.M.,Kang,H.K.,2012.Theanticancereffectoffucoidaninpc-3 prostatecancercells.Mar.Drugs11,2982–2999.
Canavese,M.,Santo,L.,Raje,N.,2012.Cyclindependentkinasesincancer:potential fortherapeuticintervention.CancerBiol.Ther.13,451–457.
Cho,T.M.,Kim,W.J.,Moon,S.K.,2014.Aktsignalingisinvolvedinfucoidan-induced inhibitionofgrowthandmigrationofhumanbladdercancercells.FoodChem. Toxicol.64,344–352.
Dobashi,Y.,Takehana,T.,Ooi,A.,2003.Perspectivesoncancertherapy:cellcycle blockersandperturbators.Curr.Med.Chem.10,2549–2558.
Fitton,J.H.,2011.Therapiesfromfucoidan;multifunctionalmarinepolymers.Mar. Drugs9,1731–1760.
Fukahori,S.,Yano,H.,Akiba,J.,Ogasawara,S.,Momosaki,S.,Sanada,S.,Kuratomi,K., Ishizaki,Y.,Moriya,F.,Yagi,M.,Kojiro,M.,2008.Fucoidan,amajorcomponent ofbrownseaweed,prohibitsthegrowthofhumancancercelllinesinvitro.Mol. Med.Rep.1,537–542.
Gong,F.Y.,Zhang,D.Y.,Zhang,J.G.,Wang,L.L.,Zhan,W.L.,Qi,J.Y.,Song,J.X.,2014. siRNA-mediatedgenesilencingofMexBfromtheMexA-MexB-OprMefflux pumpinPseudomonasaeruginosa.BMBRep.47,203–208.
Hsu,H.Y.,Lin,T.Y.,Hwang,P.A.,Tseng,L.M.,Chen,R.H.,Tsao,S.M.,Hsu,J.,2013. Fucoidaninduceschangesintheepithelialtomesenchymaltransitionand decreasesmetastasisbyenhancingubiquitin-dependentTGFreceptor degra-dationinbreastcancer.Carcinogenesis34,874–884.
Kakehi,Y.,Hirao,Y.,Kim,W.J.,Ozono,S.,Masumori,N.,Miyanaga,N.,Nasu,Y., Yokomizo,A.,2010.BladderCancerWorkingGroupreport.Jpn.J.Clin.Oncol. 40(Suppl.1),i57–i64.
Kim,S.K.,Thomas,N.V.,Li,X.,2011.Anticancercompoundsfrommarinemacroalgae andtheirapplicationasmedicinalfoods.Adv.FoodNutr.Res.64,213–224. Kim,Y.S.,Li,X.F.,Kang,K.H.,Ryu,B.,Kim,S.K.,2014.Stigmasterolisolatedfrom
marinemicroalgaeNavicula incertainduces apoptosisinhumanhepatoma HepG2cells.BMBRep.47,433–438.
Lee,H.,Kim,J.S.,Kim,E.,2012.FucoidanfromseaweedFucusvesiculosusinhibits migrationandinvasionofhumanlungcancercellviaPI3K-Akt-mTORpathways. PLoSONE7,e50624.
Lee,S.J.,Hwang,S.O.,Noh,E.J.,Kim,D.U.,Nam,M.,Kim,J.H.,Nam,J.H.,Hoe,K.L., 2014.Transactivationofbadbyvorinostat-inducedacetylatedp53enhances doxorubicin-inducedcytotoxicityincervicalcancercells.Exp.Mol.Med.46, e76.
Li,B.,Lu,F.,Wei,X.,Zhao,R.,2008.Fucoidan:structureandbioactivity.Molecules 13,1671–1695.
Liu,F.,Wang,J.,Chang,A.K.,Liu,B.,Yang,L.,Li,Q.,Wang,P.,Zou,X.,2012.Fucoidan extractderivedfromUndariapinnatifidainhibitsangiogenesisbyhuman umbil-icalveinendothelialcells.Phytomedicine19,797–803.
Park,E.J.,Pezzuto, J.M., 2013. Antioxidantmarineproducts in cancer chemo-prevention.Antioxid.RedoxSignal19,115–138.
Park,H.S.,Hwang,H.J.,Kim,G.Y.,Cha,H.J.,Kim,W.J.,Kim,N.D.,Yoo,Y.H.,Choi, Y.H.,2013.InductionofapoptosisbyfucoidaninhumanleukemiaU937cells throughactivationofp38MAPKandmodulationofBcl-2family.Mar.Drugs11, 2347–2364.
Park,H.Y.,Han,M.H.,Park,C.,Jin,C.Y.,Kim,G.Y.,Choi,I.W.,Kim,N.D.,Nam,T.J.,Kwon, T.K.,Choi,Y.H.,2011a.Anti-inflammatoryeffectsoffucoidanthroughinhibition ofNF-B,MAPKandAktactivationinlipopolysaccharide-inducedBV2microglia cells.FoodChem.Toxicol.49,1745–1752.
Park,H.S.,Kim,G.Y.,Nam,T.J.,Kim,N.D.,Choi,Y.H.,2011b.Anti-proliferativeactivity offucoidanwasassociatedwiththeinductionofapoptosisandautophagyinAGS humangastriccancercells.J.FoodSci.76,T77–T83.
Park, H.Y., Kim, G.Y., Moon, S.K., Kim, W.J., Yoo, Y.H., Choi, Y.H., 2014. FucoidaninhibitstheproliferationofhumanurinarybladdercancerT24cells
by blocking cell cycle progressionand inducing apoptosis. Molecules 19, 5981–5998.
Park,M.H.,Han,J.S.,2014.Padinaarborescensextractprotectshighglucose-induced apoptosisinpancreaticcellsbyreducingoxidativestress.Nutr.Res.Pract.8, 494–500.
Paternot,S.,Bockstaele,L.,Bisteau,X.,Kooken,H.,Coulonval,K.,Roger,P.P.,2010.Rb inactivationincellcycleandcancer:thepuzzleofhighlyregulatedactivating phosphorylationofCDK4versusconstitutivelyactiveCDK-activatingkinase.Cell Cycle9,689–699.
Racioppi, M., D’Agostino, D., Totaro, A., Pinto, F., Sacco, E., D’Addessi, A., Marangi, F., Palermo, G., Bassi, P.F., 2012. Value of current chemother-apyandsurgeryinadvancedandmetastaticbladdercancer.Urol.Int. 88, 249–258.
Riou, D., Colliec-Jouault, S., Pinczon du Sel, D., Bosch, S., Siavoshian, S., Le Bert,V.,Tomasoni,C.,Sinquin,C.,Durand,P.,Roussakis,C.,1996.Antitumor andantiproliferativeeffectsofafucanextractedfromAscophyllumnodosum againstanon-small-cellbronchopulmonarycarcinomaline.AnticancerRes.16, 1213–1218.
Saitoh,Y.,Nagai,Y.,Miwa,N.,2009.Fucoidan-VitaminCcomplexsuppressestumor invasionthroughthebasementmembrane,withscarceinjuriestonormalor tumorcells,viadecreasesinoxidativestressandmatrixmetalloproteinases. Int.J.Oncol.35,1183–1189.
Senni,K.,Pereira,J.,Gueniche,F.,Delbarre-Ladrat,C.,Sinquin,C.,Ratiskol,J.,Godeau, G.,Fischer,A.M.,Helley,D.,Colliec-Jouault,S.,2011.Marinepolysaccharides:a sourceofbioactivemoleculesforcelltherapyandtissueengineering.Mar.Drugs 9,1664–1681.
Senthilkumar,K.,Kim,S.K.,2014.Anticancereffectsoffucoidan.Adv.FoodNutr.Res. 72,195–213.
Senthilkumar,K.,Manivasagan,P.,Venkatesan,J.,Kim,S.K.,2013.Brownseaweed fucoidan:biologicalactivityandapoptosis,growthsignalingmechanismin can-cer.Int.J.Biol.Macromol.60,366–374.
Sperka,T.,Wang,J.,Rudolph,K.L.,2012.DNAdamagecheckpointsinstemcells, ageingandcancer.Nat.Rev.Mol.CellBiol.13,579–590.
Thomas,N.V.,Kim,S.K.,2014.Fucoidansfrommarinealgaeaspotentialmatrix metalloproteinaseinhibitors.Adv.FoodNutr.Res.72,177–193.
Wang,P.,Liu,Z.,Liu,X.,Teng,H.,Zhang,C.,Hou,L.,Zou,X.,2014.Anti-metastasis effectoffucoidanfromUndariapinnatifidasporophyllsinmouse hepatocarci-nomaHca-Fcells.PLoSONE9,e106071.
Wang,W.,Wang,S.X.,Guan,H.S.,2012.Theantiviralactivitiesandmechanismsof marinepolysaccharides:anoverview.Mar.Drugs10,2795–2816.
Xue,M.,Ge,Y.,Zhang,J.,Liu,Y.,Wang,Q.,Hou,L.,Zheng,Z.,2013.Fucoidaninhibited 4T1mousebreastcancercellgrowthinvivoandinvitroviadownregulationof Wnt/-cateninsignaling.Nutr.Cancer65,460–468.
Yang,L.,Wang,P.,Wang,H.,Li,Q.,Teng,H.,Liu,Z.,Yang,W.,Hou,L.,Zou,X.,2013. FucoidanderivedfromUndariapinnatifidainducesapoptosisinhuman hepa-tocellularcarcinomaSMMC-7721cellsviatheROS-mediatedmitochondrial pathway.Mar.Drugs11,1961–1976.
Zhang,Z.,Teruya,K.,Eto,H.,Shirahata,S.,2011.Fucoidanextractinducesapoptosis inMCF-7cellsviaamechanisminvolvingtheROS-dependentJNKactivationand mitochondria-mediatedpathways.PLoSONE6,e27441.