Gene cloning, expression, and characterization of the Bacillus amyloliquefaciens PS35 lipase
Texto
Imagem

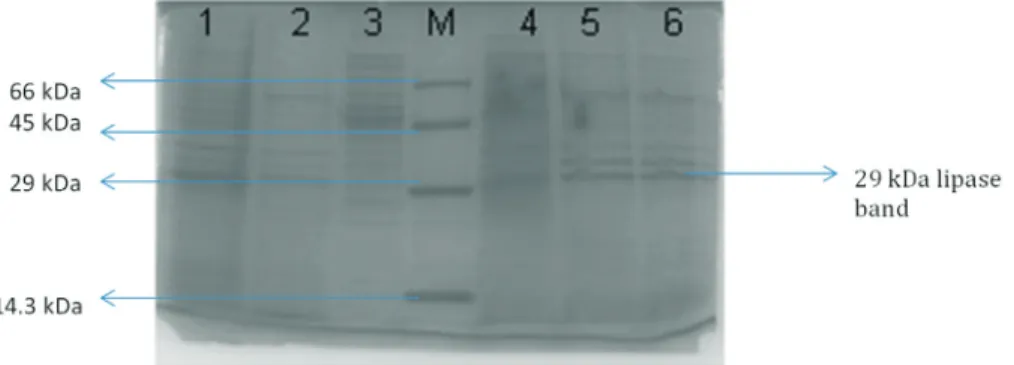
Documentos relacionados
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
O Padroado era pois um direito, um «privilégio oneroso», assente num contrato bilateral, não podendo admitir derrogação, independentemente da existência de domínio político
Although the optimum reaction temperature of the purified Aga21 was 30 °C (Figure 4A), as much as 85% of the maximum activity was observed at 10 °C.. The effect of pH on the
Effect of temperature : The levan formation during sucrose fermentation at different temperatures (25-40 0 C) was studied and the results are shown in Figure3.. but sucrase activity
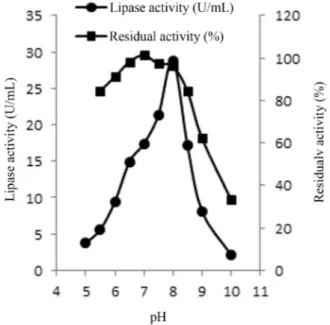
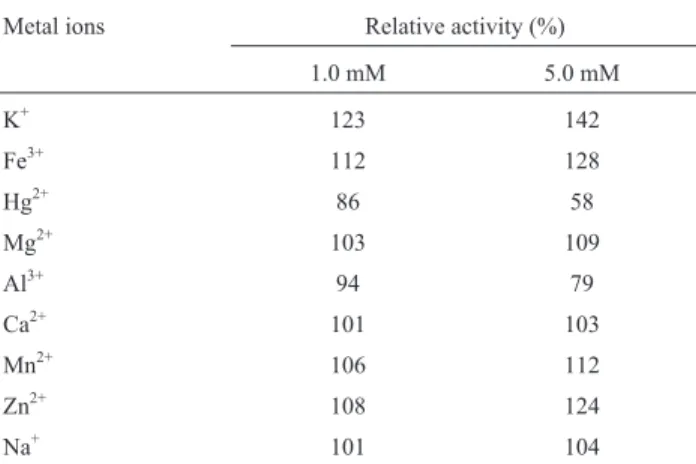
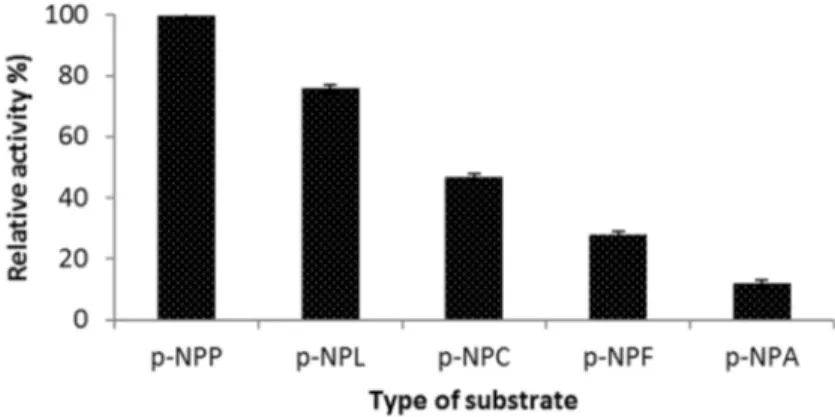
The characters analysed included effect of pH (4 - 10) and temperature (30 - 80 o C) on enzyme activity and stability, effect of NaCl (0 - 20%) on enzyme activity, effect of
a) Sujeitos de direitos – a criança e o adolescente não mais poderão ser tratados como objetos passivos de intervenção da família, da sociedade e do Estado, e sim como sujeitos com
