Vila Real, 2014
Proteogenómica de isolados de Enterococcus spp. e
Escherichia coli com a utilização do MALDI-TOF MS
Dissertação de Mestrado em Genética Molecular Comparativa e Tecnológica
Tiago João Batista Santos
Orientador: Professor Doutor Gilberto Paulo Peixoto Igrejas
Coorientador: Professor Doutor José Luís Capelo Martinez
Proteogenómica de isolados de Enterococcus spp. e
Escherichia coli com a utilização do MALDI-TOF MS
Dissertação de Mestrado em Genética Molecular Comparativa e Tecnológica
Tiago João Batista Santos
Orientador: Professora Doutor Gilberto Paulo Peixoto Igrejas
Coorientador: Professor Doutor José Luís Capelo Martinez
Composição do Júri:
________________________________________________________________ ________________________________________________________________ ________________________________________________________________ ________________________________________________________________As doutrinas apresentadas neste trabalho são da exclusiva responsabilidade do autor.
Orientador científico:
______________________________________________________________________ [Professor Doutor Gilberto Paulo Peixoto Igrejas] Departamento de Genética e Biotecnologia (IBB-CGB) Universidade de Trás-os-Montes e Alto Douro, Portugal
Coorientador científico:
____________________________________________________________________ [Professor Doutor José Luís Capelo Martinez] Departamento de Química, Bioscope Group Universidade Nova de Lisboa, Portugal
Agradecimentos
Ao terminar esta dissertação, desejo manifestar o meu verdadeiro reconhecimento pelo auxílio de diversas pessoas e entidades, que facilitaram o decorrer deste meu projeto científico e pessoal.
Ao anterior e ao atual Magnifico Reitor da Universidade de Trás-os-Montes e Alto Douro (UTAD), nas pessoas do Professor Doutor Carlos Sequeira e do Professor Doutor António Fontainhas Fernandes, pela disponibilidade e facilidades concedidas durante a realização deste trabalho.
Ao anterior e atual Diretores do Mestrado de Genética Molecular Comparativa e Tecnológica, Professor Doutor Valdemar Carnide e Professora Doutora Paula Martins-Lopes, e à Vice-Diretora, a Professora Doutora Raquel Chaves. A todos os Professores que lecionaram no Mestrado em Genética Molecular Comparativa e Tecnlógica e aos meus colegas neste segundo ciclo de estudos.
Ao meu orientador, o Professor Doutor Gilberto Igrejas, por ter prosseguido como meu guia e conselheiro na área científica, após um relatório de estágio bem sucedido. No decorrer desta dissertação, o Professor teve um papel fundamental, desde a ideia de abordar uma nova temática proteómica, até à sua ajuda em todas as etapas deste trabalho. Tenho de relevar a sua integridade e dedicação ao trabalho, um exemplo para seguir na minha carreira científica.
Ao meu coorientador, o Professor Doutor José Luís Capelo Martinez, por me ter acolhido na Universidade Nova de Lisboa e no grupo Bioscope de braços abertos e com toda a disponibilidade para partilhar o seu conhecimento científico e de vida, tendo sido uma grande ajuda nesta etapa tão importante.
À professora Irene, por toda a disponibilidade, paciência e motivação para me ensinar estatística em R, não desistindo de “picar pedra” até alcançar os resultados lapidados.
Ao Hugo, por toda a paciência, amizade e partilha de conhecimentos de química, proteómica e de MALDI-TOF MS, tendo sido essencial para me tornar confortável com esta técnica.
Ao Alexandre, pela amizade e companheirismo sempre presentes, e toda ajuda que sempre dá com um sorriso.
A todos os meus colegas do Centro de Genómica e Biotecnologia (CGB/IBB), pelos almoços, conselhos, gargalhadas, futebóis e espírito de camaradagem.
À Professora Patrícia e ao Nuno, por toda ajuda e conhecimento na Microbiologia e Antibóticos, tendo sido essenciais para o sucesso da publicação destes resultados.
Ao Professor Carlos, ao Eduardo, à Cristina, à Elisabete e a todo o grupo do Bioscope que me receberam com todo o companheirismo, ajuda e espírito académico de um grupo de investigação unido.
Ás três amigas da Caparica, Inês, Belinha e Márcia, por me terem dado um teto, companhia e uma sopa quente todos os dias.
Aos meus pais, Francisco Moreira dos Santos e Maria Arminda Morais Batista Santos. Estou-lhes eternamente grato, por sempre acreditarem em mim e me abrirem as portas ao percurso que quis percorrer, sem nunca colocarem obstáculos. Pelo vosso amor e ajuda, um eterno obrigado.
À minha irmã e o meu cunhado, Daniela e Simão, por serem a família mais próxima, que esteve lá sempre que precisei. Por terem sempre uma porta aberta e um abraço pronto para dar a este vosso irmão, um eterno obrigado.
À mulher da minha vida, Catarina Marinho, por todo o amor e amizade. Por estar lá nos momentos mais felizes e, especialmente, nos mais difíceis, ajudando-me a conseguir alcançar todos os meus objetivos.
Ao Kinder, por todos os olhares e lambidelas, quando me faltava a próxima palavra da dissertação.
Um obrigado a todos as pessoas, que mesmo não mencionadas aqui, me tenham ajudado a completar este trabalho com sucesso.
“Proteogenómica de isolados de Enterococcus spp. e Escherichia coli com a utilização do MALDI-TOF MS”
Resumo
Os Enterococcus spp. e Escherichia coli são microrganismos comensais do trato gastrintestinal da maioria dos animais e humanos, permitindo realizar estudos genéticos sobre a ocorrência e disseminação da resistência aos antibióticos. Estas bactérias apresentam, também, diferenças significativas no seu proteoma, representando bons organismos modelo para a caracterização bacteriana por novas técnicas proteómicas, como o MALDI-TOF MS. Estes dois aspetos estão relacionados com os objetivos deste trabalho: estudar a taxa de resistência a antibióticos em isolados fecais de Enterococcus spp. de aves selvagens do Arquipélago dos Açores e caracterizar, por meio de MALDI-TOF MS, isolados de E. coli e enterococos com a mesma proveniência, a nível proteómico.
A resistência aos antibióticos é um problema emergente em saúde pública pelo que, a avaliação da suscetibilidade aos antibióticos em “bactérias indicadoras”, atua como forma de controlo da disseminação. As bactérias deste estudo são reservatórios de genes de resistência que podem ser transmitidos para outras bactérias patogénicas ou comensais e, assim, representar um problema à escala mundial. A emergência de estirpes de enterococos resistentes à vancomicina (VRE) realça a inexistência de barreiras entre hospitais, população e animais. Os estudos sobre a incidência de enterococos resistentes em animais selvagens são escassos, pelo que, a sua propagação oriunda de outros biossistemas é ainda pouco compreendida. Porém, é reconhecido que os animais selvagens, incluindo as aves, albergam bactérias que atuam como reservatórios de genes de resistência passíveis de ser disseminados.
A prevalência e os mecanismos implicados na resistência aos antibióticos foram avaliados em 138 isolados Enterococcus spp. (59 Enterococcus faecalis, 40 E. faecium, 27 E.
durans e 12 E. hirae), de um total de 218 amostras fecais de aves dos Açores. Encontraram-se
percentagens elevadas de resistência nos enterococos para a tetraciclina (32,6%) e a ciprofloxacina (19,6%), sendo que 46,4% dos isolados apresentaram resistência a pelo menos um antibiótico. Pela técnica de PCR com primers específicos e corrida eletroforética dos amplicões obtidos em gel de agarose, a presença dos genes erm(B), tet(M) e/ou tet(L), e
van(A) foi confirmada, nos isolados resistentes à eritromicina, tetraciclina e vancomicina,
respetivamente. Vários estudos estão de acordo com os resultados obtidos neste trabalho, realizados noutras aves selvagens, como gaivotas e pombas. A informação conseguida é importante para fomentar o conhecimento sobre a disseminação de bactérias resistentes no
ecossistema selvagem e as possíveis implicações resultantes da transferência para outros animais ou humanos.
A técnica “Matrix Assisted Laser Desorption Ionization Time of Flight Mass
Spectrometry” (MALDI-TOF MS) é um método expedito e preciso na identificação de
bactérias intactas, em comparação com os métodos convencionais de microbiologia e/ou biologia molecular. O “fingerprinting” de proteínas por MALDI pode, também, fornecer uma solução para a caracterização das resistências a antibióticos presentes nas bactérias. A caracterização e distinção de espécies bacterianas por esta técnica tem uma potencial aplicação em laboratórios de diagnóstico.
Para a análise proteómica, via MALDI-TOF MS, proteínas solúveis foram extraídas de células bacterianas intactas, provenientes de 60 isolados de Enterococcus spp. e 60 isolados de Escherichia coli, por um método expedito. Desde o crescimento da bactéria, passando pela espetrometria de massa, até à análise bioinformática dos dados obtidos por MALDI-TOF MS, foi possível comparar 1200 espetros de massa. No total, foram obtidos 215 picos distintos, entre eles o pico de massa m/z 4428 ± 3, encontrado exclusivamente em isolados de enterococos, e outros dois picos (m/z 5379 ± 3 e 6253 ± 3) detetados apenas em espetros dos isolados de E. coli, sugerindo que estes picos possam representar biomarcadores específicos destes géneros bacterianos, respetivamente.
Este estudo foi o primeiro a fazer uma separação clara com recurso ao MALDI entre isolados de espécies diferentes do género enterococos com origem animal. Tal foi alcançado com o processamento e análise estatística, via R e MassUp. Este trabalho provou a correta separação dos isolados de espécies diferentes, atráves da análise de componentes principais (PCA), clustering e medidas estatísticas.
Os resultados da análise proteómica neste estudo vêm reforçar a capacidade da espectrometria de massa, com ionização assistida por laser, na caracterização bacteriana por célula intacta, tendo detetado picos de massa específicos dos dois géneros estudados e picos de massa caracterizadores das diferentes espécies do género enterococos. No futuro, é passível que, com uma optimização do protocolo para a obtenção de proteínas intracelulares e o desenvolvimento das ferramentas bioinformáticas, ocorra a deteção de picos de massa característicos de resistências antimicrobianas.
Palavras-chave: Enterococcus spp., Escherichia coli, resistência, antibióticos, aves selvagens,
“Proteogenomic of Enterococcus spp. and Escherichia coli isolates using MALDI-TOF MS”
Abstract
Enterococcus spp. and Escherichia coli are commensal microorganisms of the
gastrointestinal tract of most animals and humans, enabling genetic studies on the occurrence and spread of antibiotic resistance. These bacteria also show significant differences on their proteomes, representing good model organisms for bacterial characterization by new proteomic techniques such as MALDI-TOF MS. These two aspects are related with the objectives of this work: study the rate of antibiotic resistance in fecal isolates of Enterococcus spp. from wild birds of Azores Archipelago and characterize at the proteomic level isolates of
E. coli and enterococci from the same origin, by MALDI-TOF MS.
Antibiotic resistance is an emerging problem in public health and therefore, the evaluation of the susceptibility to antibiotics in "indicator bacteria" acts as a mean of controlling this spread. Bacteria of this study are reservoirs of resistance genes that can be transmitted to other pathogenic bacteria and thus represents a worldwide problem. The emergence of vancomycin-resistant enterococci strains (VRE) highlights the lack of barriers between hospitals, people and animals. Studies on resistant enterococci incidence in wild animals are scarce, so its propagation originating from others biosystems is still poorly understood. However, it's recognized that wildlife animals, including birds, act as reservoirs of resistance genes wich may be disseminated into the environment.
The prevalence and mechanisms involved in antibiotic resistance were evaluated in 138 Enterococcus spp. isolates (59 Enterococcus faecalis, 40 E. faecium, 27 E. durans, 12 E.
hirae), from a total of 218 fecal samples from azorean birds. High percentages of resistance
were found in enterococci resistant to tetracycline (32.6%) and ciprofloxacin (19.6%), whereas 46.4% of the isolates were resistant to at least one antibiotic. Using PCR with specific primers and amplicons obtained by electrophoretic run in agarose gel, the presence of
erm(B), tet(M) and/or tet(L), and van(A) genes were confirmed in isolates resistant to
erythromycin, tetracycline and vancomycin, respectively. Several studies conducted in other wild birds such as gulls and pigeons, are in agreement with the results obtained in this work. The information gained is important to raise awareness of the spread of resistant bacteria in the wild ecosystem and the implications resulting from the transfer to other animals or humans.
The Matrix Assisted Laser Desorption Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS) is a speedy and accurate method for identification of intact bacteria compared with convetional microbiology and/or molecular biology. This protein fingerprinting by MALDI can also provide a solution to the caracterization of antibiotic resistance present in bacteria. The distinguishing and characterization of bacterial species by this technique are potentialy applicable in diagnostic laboratories.
For the proteomic analysis via MALDI-TOF MS, soluble proteins were extracted from intact bacterial cells, from 60 isolates of Enterococcus spp. and 60 isolates of Escherichia coli by an expedient method. Since the growth of bacteria, throug mass spectrometry, to bioinformatic analysis of the data obtained by MALDI-TOF MS spectra it was possible to compare 1200 mass spectra. A total of 215 distinct peaks were obtained, including the mass peak m/z 4428 ± 3, exclusively found in isolates of enterococci and two other peaks (m/z 5379 ± 3 e 6253 ± 3) only detected in the spectra of E. coli isolates, suggesting that these peaks can represent specific biomarkers to these bacterial genera, respectively.
This was the first study that made a clear separation, by MALDI, between isolates from different species of the genus enterococci from animal source. This was achieved with processing and statistical analysis, via R and MassUp. This work proved the correct separation of isolates of different species through principal component analysis (PCA), clustering and statistical measures.
The results of this proteomic analysis confirmed the ability of the mass spectrometry-assisted laser ionization for characterizing whole bacterial cells, sensing specific mass peaks of the two kinds of genera studied and mass peaks that characterize the different species of enterococci. In the future, it is likely that with an optimized protocol for obtaining intracellular proteins and the development of bioinformatic tools, the detection of characteristic mass peaks associated to antimicrobial resistance will occur.
Keywords: Enterococcus spp., Escherichia coli, antimicrobial resistance, wild birds, mass
Graphical Abstract
Carduelis carduelis Erithacus rubecula
Fringila coelebs
Regulus regulus canariaSerinus
Sylvia atricapilla Sturnus
vulgaris
Coturnix
coturnix paraguaiaeGallinago Turdus merula
Enterococcus spp. Escherichia coli
Extração DNA PCR Eletroforese Extração bactéria intacta
!
MALDI-TOF MS - Espécies enterococos - Genes de resistência antimicrobiana- Caracterização de espécies bacterianas - Análise estatística por R e MassUp da identificação de microrganismos via
MALDI-TOF MS
Efeitos da resistência antimicrobiana no ecossistema aéreo
!
MALDI-TOF MS como ferramenta de caracterização microbiana e otimização de análise estatística
- n:138 isolados - E. faecalis 59 - E. faecium 40 - E. durans 27
- E. hirae 12
- E. faecalis- tetraciclina (19): tet(M) 6
tet(L) 4
tet(M)+tet(L) 9 eritromicina (9): erm(B) 6
vancomicina (1): van(A) 1
- E. faecium- tetraciclina (18): tet(M) 10
tet(L) 2
tet(M)+tet(L) 5 eritromicina (6): erm(B) 3
- E. hirae- tetraciclina (2): tet(M)+tet(L) 2 - E. durans- tetraciclina (6): tet(M) 2
tet(L) 1 tet(M)+tet(L) 3
- Classificador weka SMO (cross-validation)
!
Matriz a b c d e <- classificados 15 0 0 0 0 | a = E. coli 0 15 0 0 0 | b = E. durans 0 0 18 0 0 | c = E. faecalis 0 0 0 15 0 | d = E. faecium 0 0 0 0 12 | e = E. hirae Carduelis chlorisArtigo publicado resultante desta investigação científica:
Santos T., Silva N., Igrejas G., Rodrigues P., Micael J., Rodrigues T., Resende R., Gonçalves A., Marinho C., Gonçalves D., Cunha R., Poeta P. 2013. Dissemination of antibiotic resistant
Enterococcus spp. and Escherichia coli from wild birds of Azores Archipelago. Anaerobe.
Comunicações em poster apresentadas em reuniões científicas nacionais:
Santos T., N. Silva, A. Gonçalves, P. Rodrigues, T. Rodrigues, P. Poeta, G. Igrejas. 2012. Molecular characterization of antibiotic resistance in enterococci isolates from wild birds in Portugal. In Book of Abstracts of XXXVII Jornadas Portuguesas de Genética, Faculdade de Ciências e Tecnologia - Universidade Nova de Lisboa, Lisboa, 28-30 Maio, pp. 50.
Santos T., Capelo L. J., Santos M. H., López-Fernández H., Marinho C., Poeta P., Igrejas G. 2013. Bacterial characterization by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry and bioinformatic tools. In Book of Abstracts of XXXVIII Jornadas Portuguesas de Genética, Instituto de Ciências Biomédicas Abel Salazar/ Faculdade de Farmácia - Universidade do Porto, Porto, 4th to the 5th of June, pp. 80.
Santos T., Santos M. H., Capelo L. J., Marinho C., Poeta P., Igrejas G. 2013 Enterococcus spp. and Escherichia coli identification by Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight Mass Fingerprinting. In Book of Abstacts of MicroBiotec' 13, Universidade de Aveiro, Aveiro, 6th to the 8th of December, pp. 32.
Santos T., Capelo L. J., Oliveira I., Santos M. H., Marinho C., Poeta P. and Igrejas G. 2014. Statistical confirmation of MALDI-TOF mass spectra with R and MassUp can be used to distinguish between bacteria. In Book of Abstracts of 13th Human Proteome Organization World Congress (HUPO 2014), Madrid, Spain, October 5th-8th.
Índice
CAPÍTULO 1 – INTRODUÇÃO ... 1
1.1. BACTÉRIAS ... 1
1.1.1. O género Enterococcus ... 1
1.1.1.1. Classificação e Características gerais ... 1
1.1.1.2. Resistência a antibióticos em isolados Enterococcus spp. ... 2
1.1.2. Escherichia coli ... 3
1.1.2.1. Classificação e características gerais ... 3
1.1.2.2. Resistência a antibióticos em isolados de Escherichia coli ... 4
1.2.ANTIBIÓTICOS ... 5
1.2.1. Antibióticos utilizados neste estudo e respetivas características ... 7
1.2.2. Mecanismos de ação dos antibióticos ... 10
1.2.2.1. Inibição da síntese da parede celular ... 10
1.2.2.2. Inibição da síntese de proteínas ... 11
1.2.2.3. Inibição da síntese do ácido fólico ... 12
1.2.2.4. Ação sobre a síntese dos ácidos nucleicos ... 12
1.2.2.5. Ação sobre a permeabilidade da membrana celular ... 12
1.2.3. Mecanismos de resistência aos antibióticos ... 13
1.2.3.1. Alteração da permeabilidade da membrana ... 13
1.2.3.2. Expulsão ativa do antibiótico ... 14
1.2.3.3. Modificação ou proteção do alvo ... 14
1.2.3.4. Modificação ou inibição enzimática do antibiótico ... 14
1.2.4. Resistência a antibióticos: causas, problemas e perspetivas futuras ... 15
1.3.GENÉTICA DA RESISTÊNCIA MICROBIANA ... 17
1.3.1. Elementos genéticos de aquisição e transferência de genes de resistência ... 18
1.3.1.1. Plasmídeos ... 18
1.3.1.2 Sequências de inserção e transposões ... 20
1.3.1.3. Integrões ... 21
1.3.2. Metodologia para a caracterização de marcadores genéticos microbianos: Polymerase chain reaction (PCR) ... 22
1.4.ESPETROMETRIA DE MASSA ... 23
1.4.1. Análises bacterianas via MALDI-TOF MS: Características, Vantagens e Aplicações ... 24
1.4.2. Proteogenómica ... 31
1.4.3. Análise de massa via MALDI-TOF MS ... 32
CAPÍTULO 2 - OBJETIVOS ... 37
CAPÍTULO 3 - MATERIAL E MÉTODOS ... 39
3.1.ESTUDO DE MARCADORES GENÉTICOS (ISOLADOS BACTERIANOS DE ENTEROCOCCUS SPP.) ... 39
3.1.1. Material Biológico ... 39
3.1.1.1. Preparação das amostras para extração de DNA ... 41
3.1.2. Polymerase chain reaction (PCR) ... 42
3.1.3. Eletroforese em gel de agarose ... 43
3.2.ESTUDO PROTEÓMICO (ISOLADOS ESCHERICHIA COLI E ENTEROCOCCUS SPP.) ... 43
3.2.1. Material biológico ... 43
3.2.1.1. Preparação da amostra ... 44
3.2.2. Matrix Assisted Laser Desorption Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS) ... 45
3.2.3. Estudo bioinformático ... 46
3.2.3.1. Sem processamento de espetros: Speclust e mMass ... 46
3.2.3.2. Com processamento de espetros: R, Maldiquant e MassUp ... 47
CAPÍTULO 4 – RESULTADOS ... 51
4.1.CARACTERIZAÇÃO GENOTÍPICA (ISOLADOS ENTEROCOCCUS SPP.) ... 51
4.1.1. Resultados da classificação de espécies enterococos ... 51
4.1.2. Resultados do estudo sobre os genes de resistência ... 51
4.1.ESTUDO PROTEÓMICO (ISOLADOS ENTEROCOCCUS SPP. E ESCHERICHIA COLI) ... 58
4.1.1. Classificação do género Enterococcus spp. e Escherichia (sem processamento dos espetros) ... 58
4.1.2. Análise estatística dos espetros de massa de Enterococcus spp. e Escherichia coli (com processamento dos espetros) ... 64
CAPÍTULO 5 - DISCUSSÃO ... 71
5.1.CARACTERIZAÇÃO GENOTÍPICA:RESISTÊNCIA ANTIMICROBIANA EM ISOLADOS DE ENTEROCOCCUS SPP. ... 71
5.1.ANÁLISE PROTEÓMICA:CARACTERIZAÇÃO DE ESPÉCIES BACTERIANAS ... 74
CAPÍTULO 6 – CONCLUSÕES ... 79
Índice de Tabelas
Página
Tabela 1. Lista de classes de antibióticos e genes codificadores de resistência... 7
Tabela 2. O MALDI-TOF MS comparado com outros métodos de identificação... 28
Tabela 3. Composição do meio de crescimento Brain Heart Infusion... 41
Tabela 4. Componentes e concentrações usadas para a reação de PCR... 42
Tabela 5. Composição da solução matriz para a análise de massa... 45
Tabela 6. Isolados de Enterococcus spp. de aves dos Açores que apresentam fenótipo de resistência e os genes de resistência detetados e a caracterização subespécie... 53
Tabela 7. Resistência aos antibióticos nos isolados de Enterococcus spp. de aves selvagens dos Açores... 55
Tabela 8. Fenótipos de resistência dos isolados de Enterococcus spp. de aves selvagens dos Açores... 55
Tabela 9. Genes de resistência detetados em isolados resistentes de Enterococcus spp. de aves selvagens dos Açores... 57
Tabela 10. Número de isolados com resistência a antibióticos... 58
Índice de Figuras
Página
Figura 1. Esquema dos grupos filogenéticos... 4 Figura 2. Mecanismos de ação dos antibióticos... 10 Figura 3. A inibição da síntese da parede celular bacteriana... 11 Figura 4. Os principais mecanismos que levam à resistência antibiótica... 13 Figura 5. Diagrama da disseminação de bactérias multirresistentes... 16 Figura 6. Plasmídeos bacterianos e diagrama do factor R... 20 Figura 7. Sequência de inserção e transposão complexo... 21 Figura 8. Reação em cadeia por ação da polimerase... 23 Figura 9. Diagrama dos métodos tradicionais de identificação bacteriana... 25 Figura 10. Comparação dos fluxos de trabalho de uma identificação microbiana via MALDI-TOF MS
e a via convencional... 27
Figura 11. Diagrama da ligação e cooperação entre as diferente ómicas na identificação de
microrganismos... 32
Figura 12. Diagrama representativo do princípio de identificação de bactérias por MALDI-TOF
MS... 33
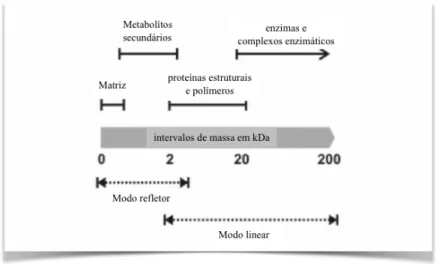
Figura 13. Tipos de compostos celulares detetados, em diferentes intervalos de massa, de célula
intacta em MS... 36
Figura 14. Número de isolados de Enterococcus spp. com fenótipos de resistência a cada antibiótico
testado... 40
Figura 15. Metodologia utilizada para processar os 240 isolados bacterianos...44 Figura 16. A placa de aço inoxidável onde foi depositada a amostra e o MALDI-TOF MS
utilizado... 45
Figura 17. Printscreen do programa de obtenção de espetros ligado ao MALDI-TOF MS... 46 Figura 18. Metodologia para o processamento de espetros e análise estatística...48 Figura 19. Distribuição das diferentes subespécies dos isolados de Enterococcus spp. das aves dos
Açores... 51
Figura 20. Distribuição dos 138 isolados de enterococos de aves selvagens no Arquipélago dos
Açores... 52
Figura 21. Número de isolados de enterococos, com genes de resistência por antibiótico... 57 Figura 22. Dois espetros de massa característicos de cada uma das espécies, Enterococcus spp. e E.
coli... 59
Figura 23. Espetro característico de Enterococcus faecalis... 60 Figura 24. Espetro característico de Enterococcus faecium... 61
Página
Figura 25. Espetro característico de Enterococcus hirae... 61 Figura 26. Espetro característico de Enterococcus durans... 62 Figura 27. Espetros obtidos por MALDI-TOF MS e processados por mMass num “gel de
espetros”... 63
Figura 28. Análise por componentes principais de espetros de massa por MassUp... 64 Figura 29. Análise por componentes principais de espetros de massa por MassUp... 65 Figura 30. Análise por componentes principais de espetros de massa por MassUp... 66 Figura 31. Análise por componentes principais de espetros de massa por R... 67 Figura 32. Análise por agrupamento de espetros numa árvore filoproteómica em MassUp... 67 Figura 33. Análise por agrupamento de espetros numa árvore filoproteómica em MassUp... 68 Figura 34. Análise por agrupamento de espetros numa árvore filoproteómica em R... 68 Figura 35. Printscreen da análise de espetros via MALDI Biotyper para identificação de isolados
Índice de Anexos
Página
Anexo I. Número de amostras fecais de acordo com a ilha do Arquipélago dos Açores e a espécie de
aves de onde foram extraídas... 89
Anexo II. Sequência nucleotídica dos primers de PCR para a identificação das espécies de
Enterococcus spp... 90
Anexo III. Sequência nucleotídica dos primers de PCR para genes codificadores de resistência a
antibióticos em Enterococcus spp... 91
Anexo IV. Número de isolados de Escherichia coli com fenótipos de resistência aos antibióticos
testados...94
Anexo V. Isolados de E. coli de aves dos Açores que apresentaram fenótipo de resistência e os genes
de resistência detetados... 95
Anexo VI. Os isolados bacterianos, e as suas características, utilizados no estudo
proteómico...96
Lista de Abreviaturas β- Beta
σ- Sigma, medida de incerteza %- Percentagem
±- Mais ou menos ºC- Graus Celsius µm- Micrometros µL- Microlitros
µg/ml- Microgramas por mililitro
30S- Subunidade pequena do ribossoma procariota 50S- Subunidade grande do ribossoma procariota
aac- Gene associado à resistência à gentamicina aadA- Gene associado à resistência à estreptomicina
ACN- Acetonitrilo
AFLP- Polimorfismos de comprimento de fragmentos amplificados (Amplified fragment
length polymorphism)
AK- Amicacina
AMC- Amoxicilina + ácidio clavulânico AMP- Ampicilina
aph- Gene associado à resistência à canamicina
ATM- Aztreonam
BHI- Meio de cérebro e coração (brain heart infusion)
bla- Gene codificador de β-lactamases de amplo espetro cat- Gene associado à resistência ao cloranfenicol
CAZ- Ceftazidima CHL- Cloranfenicol
chu(A)- Gene codificador de um recetor da membrana externa usado na identificaçãoo de
CI- Ionização química CIP- Ciprofloxacina
CLSI- Instituto de padrões clínicos e de laboratório (clinical and laboratory standards
institute)
CTX- Cefotaxima Da- Dalton
DNA- Ácido desoxirribonucleico (desoxiribonucleic acid) dNTP’s- Deoxinucleotídeo trifosfato
E. casseliflavus- Enterococcus casseliflavus E. coli- Escherichia coli
E. durans- Enterococcus durans E. faecalis- Enterococcus faecalis E. faecium- Enterococcus faecium E. gallinarum- Enterococcus gallinarum E. hirae- Enterococcus hirae
E.U.A.- Estados Unidos da América
EDTA- Ácido etilenodiamino tetra-acético (ethylenediamine tetraacetic acid) ELISA- Ensaio imunoenzimático (enzyme-linked immunosorbent assay)
erm- Gene associado à resistência à eritromicina
ERY- Eritromicina
ESBL’s- β-lactamases de amplo espetro ESI- Electrospray
FAB- Bombardeamento atómico FOX- Cefotaxima
g/L- Gramas por litro GEN- Gentamicina
IBK- Classificador estatístico IMP- Imipenemo
Intl- Gene codificador da integrase
IFA- Ensaio de imunofluorescência indireta IS- Sequências de inserção
KAN- Canamicina KDa- Kilodalton
LD- Dessorção por laser libsvm- classificador estatístico m- Massa
M- Molar
m/z- Massa por carga
MALDI-TOF MS- Espetometria de massa com ionização/dessorção assistida de laser de uma matriz com medida do tempo de voo (matrix-assisted laser desorption/ionization time of
flight mass spectromety)
MgCl2- Cloreto de magnésio
MLST- Sequênciação multilocus (multilocus sequence typing) mL- Mililitros
mM- Milimolar
mRNA- RNA mensageiro
mzXML- Tipo de ficheiro com espetro de massa/carga n- Número
NAL- Ácido nalidíxico ns- Nanosegundos
PABA- Ácido 4-aminobenzoico
PCA- Análise de componentes principais (principal component analysis) PCR- Reação em cadeia por ação da polimerase (polymerase chain reaction) PFGE- Eletroforese em gel de campo pulsado (pulsed-field gel electrophoresis) pH- Potencial hidrogénico
pmol/µL- Picomole por microlitro
PubMed- Banco de dados de artigos científicos do NCBI QD- Quinupristina-dalfopristina
RAPD- DNA polimórfico amplificado aleatoriamente (random amplified polymorphic DNA) RBF- classificador estatístico
RFLP- Polimorfismo no comprimento dos fragmentos de restrição (restriction fragment
length polymorphism)
RNA- Ácido ribonucleico (ribonucleic acid) RNAses- Ribonucleases
rRNA- RNA ribossómico rpm- Rotações por minuto
RT-PCR- PCR com transcrição reversa RTF- Fatores de transferência de resistência s- Score de coincidência
SB- Meio de Slanetz-Bartley SMO- Classificador estatístico
SNP- Polimorfismo de um único nucleótido (single-nucleotide polymorphism) STR- Estreptomicina
str- Gene associado à resistência à estreptomicina
STX- Sulfametoxazol-trimetroprim
TBE- Solução tampão de Tris, ácido bórico e EDTA TCA- Ácido tricloroacético
TE- Solução tampão de Tris e EDTA TEI- Teicomicina
TET- Tetraciclina
tet- Gene associado à resistência à tetraciclina
TFA- Ácido trifluoroacético Tn- Transposão
TOB- Tobramicina tRNA- RNA transferência
TspE4.C2- Fragmento de DNA, usado na identificação de grupos filogenéticos UTAD- Universidade de Trás-os-Montes e Alto Douro
UV- Ultravioleta U- Unidade
U/µL- Unidade por microlitro V- Volts
VAN- Vancomicina
van- Gene associado à resistência à vancomicina
VNTR- Número variável de repetições em tandem (variable number tandem repeat) VRE- Enterococos resistentes à vancomicina (vancomycin-resistant enterococci)
Nota: Esta dissertação segue as normas do novo acordo ortográfico.
Capítulo 1 – Introdução
1.1. Bactérias
Enterococcus spp. e Escherichia coli são bactérias particularmente importantes no
controlo da evolução da resistência aos antibióticos em diferentes ecossistemas (van den Bogaard e Stobberingh, 2000b). Estas bactérias possuem uma capacidade excelente para adquirir e propagar genes de resistência, mesmo entre bactérias filogeneticamente afastadas e, por estas razões, podem expor um problema de escala mundial, com consequências em saúde pública (Costa et al., 2006a; Martel et al., 2003).
1.1.1. O género Enterococcus
1.1.1.1. Classificação e Características gerais
O nome “entérocoque” foi primeiramente sugerido por Thiercelin em 1899, com o propósito de enfatizar a origem intestinal deste novo diplococos Gram-positivo (Murray, 1990). O género enterococos pertence ao grupo de bactérias acido-lácticas e está presente num largo espetro de animais e produtos derivados. Os Enterococcus spp. são organismos anaeróbios facultativos que têm forma ovóide e, tal como os estreptococos, estas bactérias não têm enzimas citocromo e são, por isso, negativas para a catalase. O habitat intestinal é o mais reconhecido para os enterococos, especialmente em mamíferos e aves. As espécies deste género, quando encontradas fora do intestino, são interpretadas como indicadores de poluição fecal. Os enterococos são notavelmente resistentes a condições ambientais adversas, como por exemplo, à seca (Devriese et al., 1993). Nos últimos anos, os Enterococcus spp. tornaram-se a principal causa de infeções nosocomiais, especialmente devido à sua elevada resistência à maioria dos antibióticos. Enterococcus faecalis e Enterococcus faecium são as duas espécies mais comummente encontradas em infeções de feridas cirúrgicas e do trato urinário. Em ambientes hospitalares, entram frequentemente na corrente sanguínea através de procedimentos invasivos, como processos com cateteres (Tortora et al., 2012).
Os enterococos atuam, também, sobre os alimentos, causando deterioração ou um melhor amadurecimento do produto e processos de aromatização de certos alimentos. As
propriedades favoráveis dos enterococos levaram à inclusão de E. faecalis, E. faecium e E.
durans em culturas iniciais de certos tipos de queijo, possibilitando alcançar bases constantes
em determinadas propriedades. A presença de enterococos no abastecimento de água potável é também monitorizada. Estirpes de E. faecium e E. faecalis sobrevivem muito mais tempo do que outras bactérias na água; a sua presença, na ausência de Escherichia coli, indica fonte de contaminação distante (Devriese et al., 1993).
Certas estirpes de enterococos são utilizadas como probióticos, sendo aplicadas para prevenir ou tratar doenças entéricas em humanos e animais (Devriese et al., 1993; Quintela-Baluja et al., 2013).
1.1.1.2. Resistência a antibióticos em isolados Enterococcus spp.
Os Enterococcus spp. são capazes de causar sérias infeções incluindo bacteremia, endocardite e meningite. A principal razão para a sua proeminência a nível mundial está relacionada com o facto destas bactérias serem intrinsecamente resistentes a um largo número de agentes antimicrobianos e, também, com a sua habilidade para adquirir novos determinantes de resistência (Moellering, 1991). Os enterococos têm a capacidade de adquirir resistência a glicopeptídos, aminoglicósidos, entre outros, colocando um obstáculo às opções terapêuticas (Aarestrup e Seyfarth, 2000).
A resistência a todos os antibióticos beta-lactâmicos disponíveis é uma característica de todos os enterococos de origem natural, incluindo aqueles que nunca foram expostos a agentes antimicrobianos em ambiente hospitalar. Devido a esta resistência intrínseca e à tolerância a antibióticos que inibem a síntese da parede celular, a combinação terapêutica de penicilina e vancomicina tem sido a ferramenta de controlo de várias infeções envolvendo bactérias deste género. A propensão dos enterococos para desenvolverem novos mecanismos de resistência a antibióticos é uma das razões para terem sido os primeiros organismos a adquirir resistência transferível à vancomicina. Os enterococos com resistência à vancomicina foram caracterizados, na maioria, como E. faecium mas, também E. faecalis já apresentou resistência a este antibiótico. A resistência a este tipo de antibióticos é medida por seis tipos de genes van: vanA, B, D, E e G, e também vanC, que é intrínseca a E. gallinarum, E.
casseliflavus e E. flavescens (Griffin et al., 2012). A habilidade dos enterococos para
desenvolverem resistência a uma larga seleção de antibióticos, representa um grande desafio para o desenvolvimento de agentes antimicrobianos efetivos (Moellering, 1991). Esta mesma
resistência natural aos antibióticos conferem-lhes vantagem em ambientes onde o uso destes é frequente e, daí, a sua relevância como agente de infecções adquiridas nos hospitais - infecções nosocomiais (Aarestrup e Seyfarth, 2000).
1.1.2. Escherichia coli
1.1.2.1. Classificação e características gerais
Escherichia coli, identificada por Theodor Escherich em 1885 (Shulman et al., 2007),
tem forma de bacilos Gram-negativos, pertencentes à família Enterobacteriaceae (Sousa, 2006). Esta família é constituída por bactérias aeróbias-anaeróbias facultativas, asporogénicas, imóveis ou móveis, com flagelos perítricos, fermentadoras da glucose, produtoras de catalase e citocromo-oxidase negativa.
Os membros da Enterobacteriaceae são, também, reconhecidos pela sua capacidade de transferência de DNA entre espécies bacterianas, homólogas e não homólogas, leveduras, plantas e células animais (Blake et al., 2003). E. coli é uma bactéria com estrutura celular simples. Contém um nucleóide superenrolado com 3 milhões de pares de base codificando 4700 genes e 2500 proteínas (Bennett, 2008). Este bacilo tem uma parede celular Gram-negativa, sendo assim passível de ser detetado pela técnica de Gram, apresentando uma cor avermelhada (Sousa, 2006).
Esta bactéria é predominantemente comensal, possuindo, por vezes, um potencial patogénico. E. coli é comummente encontrada no trato intestinal de uma grande variedade de humanos e animais (Sorum e Sunde, 2001), onde desempenha um papel protetor do intestino contra infeções, através da competição com outras bactérias intestinais patogénicas (Sokurenko et al., 1998; Souza et al., 1999), e dissemina-se para diferentes ecossistemas através da cadeia alimentar e água (van den Bogaard e Stobberingh, 2000a). É frequentemente isolada em casos de infeção extraintestinal (Duriez et al., 2001) e, por este motivo, E. coli é uma das bactérias mais estudadas, sendo usada como sistema modelo para diversos estudos nas várias áreas da biologia (Bingen et al., 1998; Lecointre et al., 1998). As estirpes patogénicas de E. coli têm diferentes níveis de gravidade para o hospedeiro, algumas com a potencialidade de provocar uma série de doenças e aumento das taxas de mortalidade (Lecointre et al., 1998; Sokurenko et al., 1998; Souza et al., 1999). A nível clínico, esta bactéria é causadora de infeções urinárias, pneumonias, septicemias, abcessos entre outras.
Algumas estirpes de E. coli podem ainda ocasionar perturbações entéricas, como é o caso do serótipo O157:H7 (Sousa, 2006).
Uma técnica rápida e eficiente para determinar o grupo filogenético em isolados de E.
coli baseia-se na deteção de dois genes (chuA e yjaA) e um fragmento de DNA (TspE4.C2),
através da reação em cadeia por ação da polimerase (PCR) (Clermont et al., 2000) (Figura 1). Vários estudos confirmam que as estirpes virulentas se encontram especialmente no grupo B2 e em parte no grupo D; porém, a maioria das estirpes comensais são identificadas no grupo A ou B1 (Bingen et al., 1998; Johnson e Stell, 2000; Picard et al., 1999). Inicialmente, a presença do gene chuA é analisada por PCR com primers específicos. Nos isolados que possuirem o gene chuA é realizada nova PCR, para o gene yjaA: os que apresentarem o gene são incluídos no grupo B2 e os negativos no grupo D. Nos isolados negativos para chuA, é pesquisado o fragmento TspE4.C2: os que apresentarem o fragmento são grupo B1 e os que são negativos grupo A (Clermont et al., 2000).
Figura 1. Esquema dos grupos filogenéticos (Clermont et al., 2000).
1.1.2.2. Resistência a antibióticos em isolados de Escherichia coli
A resistência das bactérias aos antibióticos é um grave problema de saúde, afetando múltiplas regiões do globo (Organization, 2011b). A potencialidade das bactérias para evoluírem e apresentarem resistência aos antibióticos foi prevista por Alexander Fleming em 1945, ao observar que as bactérias sensíveis à penicilina tornavam-se resistentes após várias culturas na presença de concentrações crescentes do antibiótico. No entanto, esta resistência microbiana continua a ser subestimada (Bennett, 2008; Ploy et al., 2000). A utilização de um único antibiótico causa não só uma seleção direta de estirpes resistentes como, também, leva ao desenvolvimento de cross-resistance (resistência a antibióticos relacionados
estruturalmente) e coresistance (resistência a antibióticos estruturalmente não relacionados), através de mecanismos específicos de resistência (Harada e Asai, 2010). A ameaça da resistência antimicrobiana é ainda mais problemática, devido ao potencial da microflora comensal para manter e transmitir genes de resistência a antibióticos, tal como acontece com
Escherichia coli (Blake et al., 2003).
Um dos mecanismos de resistência mais relevante é a atividade das β-lactamases de amplo espectro (ESBL’s) (Pinto et al., 2010). A produção de ESBL’s foi registada em bactérias de diferentes ecossistemas, desde áreas hospitalares, humanos, animais selvagens, animais para produção alimentar até alimentos de diferentes países, disseminando-se através de múltiplos mecanismos (Brinas et al., 2005a; Brinas et al., 2005b; Coque et al., 2008; Costa
et al., 2006b; Jouini et al., 2007; Machado et al., 2008; Poeta et al., 2009). O uso exagerado
de antibióticos em humanos e animais é, mais uma vez, a razão contributiva para a disseminação de ESBL’s e o potencial de ações diretas para controlar esta disseminação global tem de ser mais intensa, pois teme-se atingir um ponto sem retorno (Coque et al., 2008).
1.2. Antibióticos
Após a correlação entre microrganismos e doenças, começou a investigação de substâncias com capacidade de destruir microrganismos patogénicos, sem provocar dano a humanos ou animais infetados. O tratamento de doenças com substâncias químicas é denominado quimioterapia. Os produtos químicos produzidos por bactérias e fungos, que têm ação contra outros microrganismos, são antibióticos. Agentes quimiotrápicos preparados a partir de químicos, em laboratório, são designados drogas sintéticas e o sucesso da quimioterapia é baseado no facto de que, certos químicos são mais tóxicos para os microrganismos do que para os hospedeiros infetados (Willey et al., 2008).
Paul Ehrlich, físico alemão, deu origem à revolução na quimioterapia. Elrlich especulou acerca da “bala mágica” que perseguiria e destruiria um determinado patogénico, sem ferir o hospedeiro. Em 1910, após testar centenas de substâncias, ele descobriu um agente quimiotrápico chamado salvarsan, um derivado do arsénio com sucesso contra a sífilis. Pelo final dos anos 30, vários investigadores desenvolveram outras drogas sintéticas passíveis de destruir microrganismos. Além disso, as sulfonamidas foram sintetizadas por esta altura,
quando foram capazes de controlar infeções de estreptococos em ratos (Tortora et al., 2012; Willey et al., 2008).
Em contraste com as sulfonamidas, que foram desenvolvidas por uma série de químicos industriais, o primeiro antibiótico foi descoberto por acidente. Alexander Fleming, nas suas investigações, reavaliou as colónias presentes em algumas placas contaminadas. À volta do bolor encontrava-se uma área clara onde o crescimento bacteriano tinha sido inibido. Fleming estava perante um bolor que conseguia inibir o crescimento bacteriano. Esse bolor foi, mais tarde, identificado como Penicillium chrysogenum, e o inibidor, apelidado por Fleming em 1928, de penicilina. Em suma, a penicilina é um antibiótico produzido por um fungo (Manten, 1963; Sousa, 2006).
O consumo em massa da penicilina só ocorreu na década de 40. Selman Waksman anunciou em 1944 a descoberta de um novo antibiótico, a estreptomicina, que foi a primeira droga a ser eficaz contra a tuberculose. A designação antibiótico foi usada por Waksman, para denominar todos os compostos naturais produzidos por microrganismos, com capacidade para inibir o crescimento microbiano ou com efeito microbicida (Sousa, 2006). Microrganismos produtores de cloranfenicol, neomicina e tetraciclina foram isolados em 1953 (Jayaraman, 2008; Tortora et al., 2012).
A geração pós segunda guerra mundial, por parte de Howard Flory, foi a primeira a usufruir dos benefícios destes novos tratamentos, que trouxeram o controlo e abolição de certas doenças, pela descoberta e desenvolvimento de múltiplos antibióticos (Bennett, 2008; Chroma e Kolar, 2010).
De forma genérica, os antibióticos podem ser classificados em bactericidas ou bacteriostáticos, dependendo se o fármaco causa diretamente a morte da bactéria ou se inibe a sua replicação, respetivamente. Algumas drogas têm um espetro de atividade antimicrobiana estreito, por exemplo, a penicilina G afeta as bactérias Gram-positivas mas muito poucas negativas. Antibióticos que afetam ambas as bactérias positivas e Gram-negativas são classificados, por isso, como antibióticos de largo espetro. Os agentes antimicrobianos são essenciais no combate a variadas infeções bacterianas em humanos e animais, e são, ainda, usados como promotores de crescimento animal em certos países (Aarestrup, 1999; Schwarz et al., 2001). A descoberta dos agentes quimiotrápicos e o seu desenvolvimento, aumentando a sua eficácia, transformou radicalmente a medicina moderna e aliviou, de forma considerável, o nível de mortalidade associada a determinadas infeções (Willey et al., 2008).
1.2.1. Antibióticos utilizados neste estudo e respetivas características
Na Tabela 1 estão descritas as classes de antibióticos que foram usadas para este trabalho, com os seus respetivos antibióticos e genes codificadores de resistência investigados.
Tabela 1. Lista de classes de antibióticos e genes codificadores de resistência, adaptado de Manual de
Antibióticos Antibacterianos de João Carlos de Sousa (Sousa, 2006).
Mecanismo de Ação
Classe Antibiótico Genes de Resistência
Inibição da síntese da parede celular
(A) Glicopeptidos Vancomicina (VAN) van(A);van(B); van(C1);van(C2)
(A) Glicopeptidos Teicoplanina (TEI)
(B) Penicilinas Ampicilina (AMP)
Inibição da síntese de proteínas
(C) Tetraciclinas Tetraclina (TET) tet(M); tet(L)
(D) Aminoglicósidos Gentamicina (GEN) aac(6')
(D) Aminoglicósidos Canamicina (KAN) aph(3’)-IIIA
(D) Aminoglicósidos Estreptomicina (STR)
(E) Estreptogramina Quinupristina-dalforistina
(QD) vat(D); vat(E)
(F) Macrólidos Eritromicina (ERY) erm(A); erm(B); erm(C)
(G) Cloranfenicol Cloranfenicol (CHL) catA
Inibição da síntese de DNA
(H) Quinolonas Ciprofloxacina (CIP)
(A) Glicopeptidos
A vancomicina é um antibiótico glicopeptídico produzido por Streptomyces orientalis. A porção peptídica da vancomicina bloqueia a reação de transpeptidação de uma porção do peptidoglicano. O antibiótico é bactericida para os Staphylococcus e alguns membros dos géneros Clostridium, Bacillus, Streptococcus e Enterococcus. Porém, estirpes de
Enterococcus resistentes à vancomicina alastraram-se, bem como casos registados de Sthapylococcus aureus resistentes. A teicoplanina é, também, um antibiótico glicopeptídico,
produzido pelo actinomicete Actinoplanes teichomyceticus, com estrutura e mecanismo similar à vancomicina, mas com menores efeitos secundários (Willey et al., 2008).
(B) Penicilinas
A característica mais importante das penicilinas é a presença de um anel beta-lactâmico, essencial para a sua bioatividade. As suas estruturas são semelhantes à cadeia peptidica terminal presente nas subunidades do peptidoglicano. Supõe-se que esta estrutura bloqueie a enzima que catalisa a reação que forma o peptidoglicano. A síntese da parede celular é bloqueada, levando à lise da célula. Este mecanismo confirma a teoria de que as penicilinas atuam, apenas, em bactérias em crescimento. As penicilinas têm a capacidade de se ligar a várias proteínas periplasmáticas, levando à destruição da bactéria pela ativação das enzimas autolíticas. O aumento das resistências às penicilinas naturais levou ao desenvolvimento de análogos semissintéticos, como a meticilina e a oxacilina, que não são destruídos por beta-lactamases (Sousa, 2006).
(C) Tetraciclinas
As tetraciclinas são um grupo de antibióticos de largo espetro, produzidos por
Streptomyces spp. As tetraciclinas interferem com a ligação do tRNA, que transporta os
aminoácidos para a porção 30S do ribossoma, impedindo a adição de aminoácidos na cadeia de polipeptídeos em crescimento. Os ribossomas de mamíferos não sofrem interferência, pois estes compostos não têm capacidade de entrar em células mamíferas intactas. As tetraciclinas são usadas em várias infeções do trato intestinal, pneumonias e clamidíase (Sousa, 2006).
(D) Aminoglicósidos
Os aminoglicósidos ligam-se à subunidade 30S ribossomal, que interfere na síntese proteica, inibindo diretamente o processo de síntese, ou causando erros na leitura do mRNA. Estes antibióticos são bactericidas e tendem a ser mais eficazes contra patogéneos Gram-negativos. A eficácia da estreptomicina tem reduzido, devido ao aumento da resistência a este antibiótico, mas ainda é eficaz contra a tuberculose. A gentamicina é usada contra infeções causadas por Escherichia coli e Klebsiella (Willey et al., 2008).
(E) Estreptograminas
Como anteriormente referido, os patogéneos resistentes à vancomicina são um grave problema médico. Uma das poucas respostas a este problema são os antibióticos do grupo das estreptograminas. O primeiro antibiótico deste grupo a ser produzido foi a quinupristina-dalfopristina. Este tem a capacidade de bloquear a síntese proteica, ligando-se à porção 50S
do ribossoma, tal como outros antibióticos, como o cloranfenicol. A dalfopristina bloqueia uma fase inicial da síntese proteica, enquanto que a quinupristina atua numa fase tardia. A combinação das duas causa a libertação de cadeias peptídicas incompletas. Este antibiótico é eficaz num largo espetro de bactérias Gram-positivas resistentes a antibióticos. Apesar da sua eficácia, este antibiótico tem custos elevados e uma alta incidência de efeitos secundários adversos (Tortora et al., 2012).
(F) Macrólidos
Os antibióticos macrólidos são normalmente bacteriostáticos, um exemplo é a eritromicina. Ligam-se à subunidade 23S rRNA da subunidade ribossomal 50S, por forma a inibir o alongamento da cadeia peptídica durante a síntese proteica. A eritromicina é um antibiótico de largo espetro, sendo eficaz em bactérias Gram-positivas, micoplasmas e algumas bactérias Gram-negativas. Este antibiótico é utilizado em pacientes alérgicos a penicilinas (Willey et al., 2008).
(G) Cloranfenicol
O antibiótico cloranfenicol inibe a formação de ligações peptídicas da cadeia polipeptídica em crescimento, reagindo com a subunidade 50S do ribossoma procariótico. Estes antibióticos têm uma estrutura simples, sendo mais rentáveis de sintetizar quimicamente do que isolar de Streptomyces. O cloranfenicol tem um largo espetro de atividade e, devido à sua massa molecular pequena, difunde-se por todo o corpo do paciente; porém, tem efeitos secundários graves, como a supressão da atividade da medula óssea (Winston et al., 2004).
(H) Quinolonas
As quinolonas são drogas sintéticas que inibem a síntese de ácidos nucleicos e têm atividade bactericida. Estes antibióticos são usados para tratar uma ampla variedade de infeções. A primeira quinolona sintetizada foi o ácido nalidíxico, em 1962. A ciprofloxacina, também pertencente a este grupo e ganhou notoriedade durante os ataques bioterroristas de 2001, nos E.U.A., por ser a primeira escolha de tratamento contra o anthrax. As quinolonas inibem a DNA girase e a topoisomerase II bacteriana. A DNA girase introduz torção negativa ao DNA, ajudando a separar as cadeias pelo que, a sua inibição resulta na interrupção da replicação e reparação de DNA (Willey et al., 2008).
1.2.2. Mecanismos de ação dos antibióticos
Existem vários mecanismos de ação dos antibióticos, havendo diferentes antibióticos atuantes em cada mecanismo (Figura 2): inibição da síntese da parede celular; inibição da síntese da proteína; inibição da síntese do acido fólico; ação sobre a síntese dos ácidos nucleicos e ação sobre a permeabilidade da membrana.
Figura 2. Mecanismos de ação dos antibióticos. Adaptado de (Cummings, 2008).
1.2.2.1. Inibição da síntese da parede celular
A penicilina, o primeiro antibiótico descoberto e utilizado, é um exemplo de inibidor da síntese de parede celular. A parede celular é uma complexa rede macromolecular, chamada de peptidoglicano, e é apenas encontrado em paredes celulares bacterianas. A penicilina e outros antibióticos previnem a síntese de peptidoglicano e, consequentemente, a parede celular é grandemente enfraquecida, levando à lise celular (Figura 3). Visto que estes antibióticos têm como alvo o processo de síntese, estes apenas têm efeito em células em crescimento. Deste modo, o facto das células humanas não possuírem peptidoglicano resulta numa reduzida toxicidade para as células hospedeiras. É um dos grupos mais importantes na
veterinária, pois tem grande eficácia, com baixa toxicidade para os animais. São exemplo as penicilinas, cefalosporinas e a vancomicina (Lima et al., 2013; Sousa, 2006).
Figura 3. A inibição da síntese da parede celular bacteriana. À esquerda a bactéria antes da ação da penicilina; à
direita a lise bacteriana por ação do antibiótico (Tortora et al., 2012).
1.2.2.2. Inibição da síntese de proteínas
Visto que a síntese de proteínas é uma característica comum a todas as células, procarióticas e eucarióticas, a toxicidade seletiva parece ser um alvo pouco provável dos antibióticos que atuam por este método. Porém, uma diferença substancial entre procariotas e eucariotas é a estrutura dos seus ribossomas. As células eucarióticas têm ribossomas 80S, enquanto que os procariotas têm ribossomas 70S. A diferença da estrutura ribosomal é a razão pela seletividade tóxica dos antibióticos que atuam na síntese de proteínas (Tortora et al., 2012). A inibição da síntese proteica é realizada pelos antibióticos via ligação aos alvos ribossomais exercendo antibiose. Os ribossomas são os organelos que fazem a tradução do mRNA para proteínas. Nas bactérias, os ribossomas têm duas subunidades, 30S e 50S, havendo antibióticos que inibem uma ou outra subunidade.
Um dos grupos que atua deste modo são os aminoglicósidos. Deste grupo, à que destacar a amicacina, a gentamicina, a canamicina, a neomicina, a estreptomicina e a tobramicina, todos estes são antibióticos que entram no citoplasma bacteriano, ligando-se à subunidade 30S dos ribossomas, causando alterações na síntese proteica pela incorreta leitura do mRNA (Sousa, 2006). Por sua vez, o cloranfenicol faz a ligação à subunidade 50S, procedendo ao bloqueio da atividade da peptidil transferase, que vai afetar a junção dos novos aminoácidos à cadeia peptídica em formação. A estreptogramina e o grupo dos macrólidos atua na subunidade 50S ribossomal, não permitindo a fase de alongamento das proteínas. São
exemplo os antibióticos aminoglicósidos, tetraciclinas, macrólidos e estreptograminas (Lima
et al., 2013).
1.2.2.3. Inibição da síntese do ácido fólico
Uma atividade particular dos microrganismos pode sofrer competição inibitória, por uma determinada substância que tenha elevada semelhança com a enzima normal. Um exemplo de inibição competitiva é a relação entre a droga sulfa e o ácido para-aminobenzóico (PABA). Em muitos microrganismos, o PABA é o substrato para uma reação química que leva à síntese de ácido fólico, uma vitamina que funciona como coenzima para a síntese das bases purina e pirimidina. Um grupo que causa a inibição do ácido fólico é o das sulfonamidas, onde se encontra o trimetoprim. Assim, a cadeia metabólica é quebrada e não ocorre a formação do ácido di-hidropteorato, mas sendo, na sua ausência, possível a síntese do ácido fólico (Sousa, 2006).
1.2.2.4. Ação sobre a síntese dos ácidos nucleicos
Os antibióticos que interferem com o processo de síntese dos ácidos nucleicos atuam no processo de replicação do DNA e transcrição nos microrganismos. Algumas das drogas deste tipo são de aplicabilidade limitada, pois podem interferir de forma perigosa com o DNA do hospedeiro. A síntese dos ácidos nucleicos pode ser afetada pela ação dos antibióticos, como são exemplo as quinolonas. Estes antibióticos inibem a DNA girase e a topoisomerase IV, enzimas topoisomerases do tipo II intervenientes no superenrolamento do DNA, ocorrendo, desta forma, um impedimento da replicação e transcrição do DNA (Jacoby, 2005; Sousa, 2006).
1.2.2.5. Ação sobre a permeabilidade da membrana celular
Certos antibióticos podem causar alterações na permeabilidade da membrana plasmática que resultam na perda de metabolitos importantes para a célula microbiana. Neste mecanismo, o antibiótico liga-se aos fosfolípidos e, por modificações na estrutura, vai alterar a permeabilidade e, consequentemente, provocar a saída de elementos do interior da célula.
As polimixinas são um exemplo de um antibiótico que atua desta forma (Sousa, 2006; Tortora
et al., 2012).
1.2.3. Mecanismos de resistência aos antibióticos
Os quatro principais mecanismos pelos quais uma bactéria pode ser resistente a um determinado antibiótico estão presentes na Figura 4: alteração da permeabilidade da membrana; expulsão ativa do antibiótico; modificação ou proteção do alvo e modificação ou inibição enzimática do antibiótico.
Figura 4. Os principais mecanismos que levam à resistência aos antibióticos são: mutações genéticas, expressão
de genes de resistência “adormecidos” e aquisição de genes ou segmentos de DNA. Legenda: a- Alteração da permeabilidade da membrana; b- modificação ou proteção do alvo; c- expulsão ativa do antibiótico; d- modificação ou inibição enzimática do antibiótico. Adaptado de (Conly, 2002).
1.2.3.1. Alteração da permeabilidade da membrana
As bactérias Gram-negativas são relativamente mais resistentes aos antibióticos do que as Gram-positivas, isto devido à natureza da sua parede celular. Algumas bactérias mutantes têm os poros de passagem de substratos modificados, impedindo a entrada dos
antibióticos no espaço periplasmático. Antibióticos como os β-lactâmicos, as tetraciclinas e as fluoroquinolonas têm a sua atividade reduzida por este mecanismo (Giedraitiene et al., 2011).
1.2.3.2. Expulsão ativa do antibiótico
Determinadas proteínas transmembranares de transporte nas membranas das bactérias funcionam como bombas que expelem ativamente os antibióticos para fora das células, evitando que os mesmos alcancem concentrações necessárias para serem eficientes. Este mecanismo foi primeiramente observado com as tetraciclinas, porém, concede resistência a várias classes de antibióticos. As bactérias, normalmente, têm este tipo de proteínas para eliminar substâncias tóxicas. Um exemplo da ação deste mecanismo é sobre a tetraciclina, pois quando entra na célula, liga-se à proteína TetR, devido à sua afinidade, que funciona como repressora da bomba Tet, codificada pelo gene tetA, havendo uma sobrexpressão da bomba TetA que, por sua vez, vai levar à expulsão da tetraciclina para o exterior da célula (Chroma e Kolar, 2010).
1.2.3.3. Modificação ou proteção do alvo
A síntese de proteínas envolve o movimento do ribossoma ao longo de uma cadeia de RNA mensageiro. Vários antibióticos aminoglicósidos, tetraciclinas e macrólidos inibem a síntese de proteínas nesta altura do processo. Pequenas modificações no local de tradução podem neutralizar o efeito dos antibióticos, sem afetar significativamente a função celular. Alterações no alvo também são comuns, através da modificação da sua estrutura (Chroma e Kolar, 2010). Esta modificação gera-se por mutações no gene alvo, no entanto, esse mesmo alvo continua a ter a sua atividade celular, mas não é acessível pelo antibiótico. Um exemplo desta redução de sensibilidade para com o antibiótico ocorre com antibióticos β-lactâmicos em Staphylococcus aureus (Giedraitiene et al., 2011).
1.2.3.4. Modificação ou inibição enzimática do antibiótico
A inativação ou destruição de antibióticos por ação de enzimas afeta, principalmente, aqueles que têm origem natural, como é o caso das penicilinas e cefalosporinas. Grupos de
antibióticos totalmente sintetizados quimicamente, como fluoroquinolonas, têm menor probabilidade de serem afetadas por este mecanismo. As penicilinas e cefalosporinas, bem como os carbopenemos, partilham uma estrutura orgânica, o anel beta-lactâmico, que é alvo de enzimas beta-lactamases, que são capazes de o hidrolisar. Nas bactérias Gram-negativas, a produção de lactamases é o principal mecanismo de resistência aos antibióticos β-lactâmicos (Chroma e Kolar, 2010; Sousa, 2006).
1.2.4. Resistência a antibióticos: causas, problemas e perspetivas futuras
A disseminação e aumento de estirpes resistentes foi influenciada por vários fatores: o uso excessivo de antibióticos em humanos, a utilização desprecavida em patologias animais e o uso indevido como aditivo de rações para animais (Harada e Asai, 2010; Manten, 1963; Ploy et al., 2000). A utilização de antibióticos de forma inadequada é um problema presente, especialmente em países menos desenvolvidos, onde técnicos de saúde bem qualificados são escassos e a possibilidade de compra de antibióticos sem prescrição é comum. Mesmo quando a prescrição de antibióticos é adequada, os regimes de toma são curtos demais para erradicar a infeção, intensificando a sobrevivência de estirpes de bactérias resistentes. Também é comum o consumo de antibióticos ultrapassados, adulterados e até falsificados. Porém, os países mais desenvolvidos também contribuem, em muito, para o aumento das resistências antimicrobianas. O centro de controlo e prevenção e doenças dos E.U.A. estima as percentagens em que são utilizados os antibióticos para tratar determinados patógenos de forma desnecessária, em 30% das prescrições para infeções nos ouvidos, em 100% das prescrições para constipações e 50% das prescrições para dores de garganta. Mais de metade das 100 mil toneladas de antibióticos consumidas nos Estados Unidos, a cada ano, não são utilizadas para tratar doenças, mas para alimentação animal, como promotores de crescimento (Tortora et al., 2012).
No mundo inteiro, estima-se que este problema cause prejuízos de 28 mil milhões de euros, anuais (Organization, 2011a). O custo por detrás da resistência aos antibióticos está diretamente relacionado com o elevado custo de desenvolver novas drogas para conseguir substituir o antibiótico que perdeu a eficácia. Existem várias estratégias que podem ser adoptadas, por pacientes e entidades de saúde, para prevenir o desenvolvimento de resistências. Mesmo após a recuperação, os pacientes devem terminar o regime de toma do antibiótico, não devendo deixar sobras de antibiótico para tratar alguma doença posterior.