1
UNIVERSIDADE DO ESTADO DE SANTA CATARINA
–
UDESC
CENTRO DE CIÊNCIAS DA SAÚDE E ESPORTES
–
CEFID
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DO MOVIMENTO
HUMANO
LILIAN CARDOSO VIEIRA
EFEITOS DA SUPLEMENTAÇÃO DE TAURINA SOBRE OS
PARÂMETROS DE ESTRESSE OXIDATIVO MUSCULAR INDUZIDOS
POR EXERCÍCIO EXCÊNTRICO
FLORIANÓPOLIS
–
SC
2009
LILIAN CARDOSO VIEIRA
EFEITOS DA SUPLEMENTAÇÃO DE TAURINA SOBRE OS
PARÂMETROS DE ESTRESSE OXIDATIVO MUSCULAR INDUZIDOS
POR EXERCÍCIO EXCÊNTRICO
Dissertação de Mestrado apresentada à banca examinadora como requisito para obtenção do título de Mestre no Programa de Pós-Graduação Stricto Sensu em Ciências do Movimento Humano, da Universidade do Estado de Santa Catarina.
Orientador: Profº Dr. Magnus Benetti
3
LILIAN CARDOSO VIEIRA
EFEITOS DA SUPLEMENTAÇÃO DE TAURINA SOBRE OS
PARÂMETROS DE ESTRESSE OXIDATIVO MUSCULAR INDUZIDOS
POR EXERCÍCIO EXCÊNTRICO
Dissertação de Mestrado aprovada como requisito para obtenção do título de Mestre no Programa de Pós-Graduação Stricto Sensu em Ciências do Movimento Humano, da Universidade do Estado de Santa Catarina.
Banca examinadora:
Orientador: _________________________________________ Profº. Dr. Magnus Benetti
UDESC - SC
Membro: _________________________________________ Profº. Dr. Ricardo Aurino de Pinho
UNESC - SC
Membro: _________________________________________ Profº. Dr. Fabrizio Caputo
UDESC - SC
Membro: _________________________________________ Profª. Drª. Patrícia Faria Di Pietro
UFSC – SC
Florianópolis - SC, 30/10/2009.
5
AGRADECIMENTOS
Chegou a hora... Dizer obrigada é um gesto complexo, muitas vezes temos dificuldade de reconhecer que simples atos nos levaram a conquista tão grande. E nesses simples atos estão envolvidas pessoas que passaram tão sutilmente por nossas vidas que cometemos injustiças de não as agradecê-las. Mas, na realidade, elas estarão sempre presentes no decorrer das nossas histórias.
Um dos aprendizados que adquiri, neste processo, foi de que nada nós conseguimos por acaso. Nossas conquistas são feitas de esforços! Agradeço por todos os momentos difíceis que passei, que chorei, que me desesperei. Sei que esses momentos foram decisivos para eu atingir esta meta e contribuíram para me tornar cada vez mais forte para agüentar até aqui. Muitas vezes pedi auxílio a forças divinas, rezei e tenho certeza de que fui atendida, que existe Deus por trás de todos nós, olhando todos os nossos passos, inclusive colocando pessoas maravilhosas, outras nem tanto, em nossos caminhos. Mas tudo é aprendizado!
Não conseguiria atingir os meus objetivos sem o alicerce da minha família que sempre acreditou no meu potencial. O apoio psicológico da minha mãe e a racionalidade de meu pai foram essenciais nesse caminho. Os exemplos dos meus irmãos me incentivaram a lutar e a buscar integralmente o que realmente quero.
Em certo momento esta minha entrega integral gerou conflitos entre o meu trabalho e a minha vida acadêmica. Agradeço ao meu orientador e a Profª. Thais pelos conselhos e por terem me orientado para a escolha que fiz!
Meus agradecimentos ao meu orientador, Profº. Dr. Magnus Benetti, pela oportunidade de realizar este sonho, pelas suas idéias (que ele tem muitas), pela paciência e pelo incentivo em dar continuidade à minha vida acadêmica. Suas aulas eram verdadeiras viagens fisiológicas as quais me marcaram e me fizeram reconhecer um dos melhores professores que eu tive na vida!
(Laboratório de Fisiologia e Bioquímica do Exercício) da UNESC, Criciúma; um maravilhoso mundo de pesquisas! Também as pessoas do LAFIBE, as bolsistas (Meri, Débora, Karol) que me ensinaram muito sobre as análises e aos momentos passados no laboratório. Um obrigada do fundo do coração ao Luciano, Doutorando que foi de extrema importância neste processo.
Ao Profº. Dr. Fabrizio Caputo pelo aprendizado adquirido em suas aulas, a ajuda no projeto e a oportunidade de fazer docência orientada.
À Professora Patricia Faria Di Pietro, muito especial em minha vida por ter me orientado no meu primeiro estágio e pelas oportunidades.
Não poderia de deixar de citar aqui a administração, funcionários, jogadores e grandes amigos do Figueirense Futebol Clube que estiveram comigo em um ano e meio de mestrado, sempre me dando forças. Um agradecimento especial ao Dr. Sérgio Eduardo Parucker, Anderson Barros e Alexandre Gallo pela oportunidade de ingressar nesta profissão pela qual eu me apaixonei.
Aos colegas do Mestrado, como a Raffa e a Adri, que indiretamente fizeram parte dessa caminhada. Gilmar, pelas aulas e momentos de estudo na disciplina de Estatística. Cléber, pelas dúvidas sanadas e companheirismo.
Ao Claudio, pessoa muito especial em minha vida. Trazia a minha paz, me acalmava. Muitas vezes, longe, mas tenho certeza de que seu coração estava sempre comigo!
Minhas amigas Camilinha, Cris, Patty e Aline que estavam sempre ao meu lado nos momentos difíceis. Um agradecimento especial à Monique pela paciência, pelas longas conversas no telefone e o coleguismo até mesmo na hora de escrever o projeto.
7
MENSAGEM
Tua caminhada ainda não terminou.... A realidade te acolhe dizendo que pela frente o horizonte da vida necessita de tuas palavras e do teu silêncio.
Se amanhã sentires saudades, lembra-te da fantasia e sonha com tua próxima vitória. Vitória que todas as armas do mundo jamais conseguirão obter, porque é uma vitória que surge da paz e não do ressentimento.
É certo que irás encontrar situações tempestuosas novamente, mas haverá de ver sempre o lado bom da chuva que cai e não a faceta do raio que destrói.
Tu és jovem. Atender a quem te chama é belo, lutar por quem te rejeita é quase chegar a perfeição. A juventude precisa de sonhos e se nutrir de lembranças, assim como o leito dos rios precisa da água que rola e o coração necessita de afeto.
Não faças do amanhã o sinônimo de nunca, nem o ontem te seja o mesmo que nunca mais. Teus passos ficaram. Olhes para trás... mas vá em frente pois há muitos que precisam que chegues para poderem seguir-te.
RESUMO
VIEIRA, Lilian C. Efeitos da suplementação de Taurina sobre os parâmetros de Estresse Oxidativo Muscular induzidos por Exercício Excêntrico. 2009. Dissertação (Mestrado em Ciências do Movimento Humano) – Universidade do Estado de Santa Catarina. Programa de Pós-Graduação em Ciências do Movimento Humano, Florianópolis, 2009.
O objetivo deste estudo foi investigar os efeitos da suplementação de taurina sobre biomarcadores do estresse oxidativo após o exercício excêntrico. Vinte e quatro ratos da raça Wistar machos foram divididos nos seguintes grupos (n = 6): controle (C); exercício excêntrico (EE); exercício excêntrico com a suplementação de taurina (EET); exercício excêntrico com a suplementação de salina (EES). A quantidade de taurina administrada foi de 300mg/Kg de peso corporal / dia em solução de 1 ml de água deionizada por gavagem oral, por duas semanas antes e 2, 12, 24 e 48 horas após a sessão de exercício. Os animais foram submetidos a uma sessão única de corrida em declive com duração de 90 minutos e velocidade constante de 16,6 m.min-1. Quarenta e oito horas após a sessão de exercício, os animais foram sacrificados e os músculos do quadríceps removidos cirurgicamente. Produção do ânion superóxido, lipoperoxidação, carbonilação das proteínas, conteúdo total de tióis e atividade de enzimas superóxido dismutase (SOD) e catalase (CAT) foram analisados. Os resultados mostram que o grupo EET apresentou uma redução significativa (p<0,05) na produção do ânion Superóxido (0,9 0,2 nmol/min/mg proteína) quando comparado com o grupo EE (1,62 0,2 nmol/min/mg proteína) e o grupo EES (1,55 0,3 nmol/min/mg proteína). A lipoperoxidação foi diminuída já que EET apresentou quantidade de espécies reativas ao ácido tiobarbitúrico (TBARS) de 0,13 0,001nmol/mg proteína, significativamente menor (p<0,05) aos outros grupos (EES - 0,18 0,03 nmol/mg proteína e EE - 0,18 0,05 nmol/mg proteína). O conteúdo total de tióis apresentou-se aumentado (p<0,05) no grupo EET (18,8 0,3 nmol TNB/mg proteína) em comparação com EES (13,8 1 nmol TNB/mg proteína) e EE (12,2 1 nmol TNB/mg proteína). O grupo EET demonstrou um valor menor (p<0,05) na carbonilação de proteínas (0,20 0,2 nmol/mg proteína) em relação aos demais grupos (EES - 0,30 0,03 nmol/mg proteína; e EE - 0,30 0,025 nmol/mg proteína). Entretanto, a atividade das enzimas antioxidantes SOD e CAT não teve diferença significativa (p<0,05) com a suplementação da taurina. Os valores da atividade da SOD nos grupos foi de: EE – 1,22 0,25 U de SOD/mg proteína; EET –
1,18 0,15 U de SOD /mg proteína; EES – 1,22 0,12 U de SOD /mg proteína; e da CAT foi: EE - 6,12 0,19 U de CAT/mg proteína; EET- 5,85 0,62 U de CAT/mg proteína; EES – 5,61 0,45 U de CAT/mg proteína. Em conclusão, o estudo demonstra que a suplementação de taurina diminuiu o dano oxidativo, mas não afetou a atividade das enzimas antioxidantes após exercício excêntrico.
9
ABSTRACT
VIEIRA, Lilian C. Taurine Supplementation Effects on Muscle Oxidative Stress Induced by Eccentric Exercise. 2009. Dissertação (Mestrado em Ciências do Movimento Humano) – Universidade do Estado de Santa Catarina. Programa de Pós-Graduação em Ciências do Movimento Humano, Florianópolis, 2009.
This study was to investigate the effects of taurine supplementation on biomarkers of oxidative stress after eccentric exercise. Twenty four male rats Wistar were divided into the following groups (n=6): control (C); eccentric exercise (EE); eccentric exercise plus taurine (EET); eccentric exercise plus saline (EES). Taurine was administered with solution of 1ml water 300mg/Kg body weight (BW)/day by oral lavage, for two weeks before and 2, 12, 24 and 48 hours after the exercise session. The animals were submitted to one downhill run session with duration of 90 minutes and velocity constant of 16,6 m.min-1. Forty-eight hours after the exercise session, the animals were killed and the quadriceps muscle were surgically removed. Production of superoxide anion, lipoperoxidation, carbonylation, total thiol content and antioxidant enzyme were analyze. The results show the EET group had a significant reduction (p <0.05) in the production of superoxide anion (0,09 + 0.2 nmol / min / mg protein) when compared with the EE group (1.62 0.3 nmol / min / mg protein) and the EES group (1,55 0,3 nmol/ min/ mg protein). Lipid peroxidation was diminished as EET showed 0,13 0,001nmol/ mg protein of tiobarbituric acid reactive species (TBARS) significantly lower (p < 0,05) than other groups (EES - 0,18 0,03 nmol/mg protein and EE - 0,18 0,05 nmol/mg protein). The total thiol content were higher (p < 0,05) in EET (18,8 0,3 nmol TNB/ mg protein) when compared with EES (13,8 1 nmol TNB/ mg protein) and EE (12,2 1 nmol TNB/ mg protein). The EET group showed a lower value (p < 0.05) of proteins carbonyl (0,20 0,2 nmol/ mg protein) compared to other groups (EES - 0,30 0,03 nmol/ mg protein and EE - 0,30 0,025 nmol/ mg protein). The activity of antioxidant enzymes superoxide dismutase (SOD) and catalase (CAT) had no significant difference (p <0.05) with supplementation of taurine. The values of SOD activity in the groups were: EE – 1,22 0,25 U of SOD/ mg protein; EET – 1,18 0,15 U of SOD / mg protein; EES – 1,22 0,12 U of SOD / mg protein; and the values of CAT were: EE - 6,12 0,19 U of CAT/ mg protein; EET- 5,85 0,62 U of CAT/ mg protein; EES – 5,61 0,45 U of CAT/ mg protein. In conclusion, the present study demonstrates that taurine supplementation decreased damage oxidative but did not affect antioxidant enzyme activity after eccentric exercise.
LISTA DE ILUSTRAÇÕES
Figura 1: Desenho do estudo...43
Figura 2: Produção de ânion superóxido (O2-) no músculo do quadríceps de ratos, após uma sessão de exercício excêntrico...48
11
LISTA DE TABELAS
LISTA DE ABREVIATURAS, SIGLAS E SÍMBOLOS
µL – Microlitro µmol – Micromol
AMP – Adenosina Difosfato ATP – Adenosina Trifosfato C – Grupo Controle
Ca2+–Íon Cálcio CAT – Catalase CoQ – Ubiquinona CoQH – Semiquinona CoQH2– Ubiquinol Cu2+ - Íon Cuproso Cu3+ - Íon Cúprico
DMT – Dor Muscular Tardia e- - Elétron
EE – Grupo Exercício Excêntrico
EES – Grupo Exercício Excêntrico + Salina EET–Grupo Exercício Excêntrico + Taurina EPM – Erro Padrão Médio
ER – Espécies Reativas
ERO – Espécies Reativas de Oxigênio
FADH – Flavina Adenina Dinucleotídeo Reduzido Fe2+ - Íon Ferroso
Fe3+ - Íon Férrico
GPX – Glutationa Peroxidase GSH – Glutationa
H+ - Íon Hidrogênio H2O – Água
13
K Ca2+– Potássio dependente de Cálcio Kg – Quilo
m - Metro M – Molar
MDA – Malondialdeído mg – Miligrama
min – Minuto ml – Mililitro mM – Milimolar
NAC – N-acetil cisteína
NADH – Nicotinamida Adenina Dinucleotídeo Reduzido NADPH – Nicotinamida Adenina Dinucleotídeo Fosfato nm – Nanômetro
O2– Oxigênio
O2-– Ânion Superóxido O2H– Radical Hidroperoxil OH – Radical Hidroxil PGE – Prostaglandinas
PMN – Células polimorfonucleadas RL – Radicais Livres
RLO – Radicais Livres de Oxigênio
SOD – mt – Superóxido Dismutase mitocondrial SOD – Superóxido Dismutase
TBA – Ácido Tiobarbitúrico
TNFα –Fator de Necrose Tumoral α
SUMÁRIO
1 INTRODUÇÃO ... ....17
1.1 PROBLEMA ... 17
1.2 JUSTIFICATIVA ... 18
1.3 OBJETIVOS ... 20
1.3.1 Objetivo Geral ... 20
1.3.2 Objetivos Específicos ... 20
1.4 HIPÓTESES ... 21
1.4.1 Hipótese geral ... 21
1.4.2 Hipóteses Específicas ... 21
1.5 DELIMITAÇÕES DO ESTUDO ... 21
1.6 LIMITAÇÕES DO ESTUDO ... 21
2 REFERENCIAL TEÓRICO ... 23
2.1 RADICAIS LIVRES ... 23
2.2 PRINCIPAIS ESPÉCIES REATIVAS DE OXIGÊNIO ... 23
2.3 SISTEMA DE DEFESA ANTIOXIDANTE ... 25
2.3.1 Antioxidantes enzimáticos ... 26
2.3.1.1 Superóxido Dismutase (SOD) ... 27
2.3.1.2 Catalase (CAT) ... 28
2.3.1.3 Glutationa Peroxidase (GPX) ... 28
2.3.2 Antioxidantes não enzimáticos ... 29
2.4 TAURINA ... 30
2.4.1 Regulação dos canais de cálcio ... 32
2.4.2 Equilíbrio osmótico celular ... 32
2.4.3 Neutralização direta de ERO ... 33
2.5 EXERCÍCIO FÍSICO E ESTRESSE OXIDATIVO ... 34
2.5.1 Formação de ERO durante o met. do oxigênio na cadeia respiratória ... 35
2.5.2 Formação de ERO durante isquemia-reperfusão ... 35
15
2.7 LESÕES NO EXERCÍCIO EXCÊNTRICO ... 38
3 MÉTODOS ... 40
3.1 CARACTERIZAÇÃO DA PESQUISA ... 40
3.2 AMOSTRA ... 40
3.3 TRATAMENTO EXPERIMENTAL ... 41
3.4 PROTOCOLO DE ADAPTAÇÃO AO EXERCÍCIO ... 41
3.5 PROTOCOLO DE EXERCÍCIO EXCÊNTRICO DE LONGA DURAÇÃO ... 41
3.6 SUPLEMENTAÇÃO ... 42
3.7 EUTANASIA ... 42
3.8 DESENHO DO ESTUDO ... 43
3.9 ENSAIOS BIOQUÍMICOS ... 43
3.9.1 Produção de Espécies Reativas de Oxigênio ... 43
3.9.1.1 Ânion Superóxido ... 43
3.9.2 Marcadores de Danos Oxidativos... 44
3.9.2.1 Espécies Reativas ao Ácido Tiobarbitúrico (TBARS) ... 44
3.9.2.2 Carbonilação de proteínas ... 44
3.9.2.3 Sulfidrilas ... 45
3.9.3 Atividade de enzimas antioxidantes ... 45
3.9.3.1 Superóxido dismutase (SOD) ... 45
3.9.3.2 Catalase (CAT) ... 46
3.9.4 Determinação da Proteína ... 46
3.10 CÁLCULO E TAMANHO DA AMOSTRA ... 47
3.11 TRATAMENTO ESTATÍSTICO ... 47
4 RESULTADOS ... 48
4.1 PRODUÇÃO DE ESPÉCIES REATIVAS DE OXIGÊNIO... 48
4.1.1 Ânion Superóxido ... 48
4.2 MARCADORES DE DANOS OXIDATIVOS ... 49
4.2.1 Espécies Reativas ao Ácido Tiobarbitúrico (TBARS) ... 49
4.2.2 Conteúdo total de tióis ... 50
4.2.3 Carbonilação de proteínas ... 50
4.3.1 Superóxido dismutase (SOD) ... 51
4.3.2 Catalase (CAT) ... 52
5 DISCUSSÃO ... 54
6 CONCLUSÃO ... 59
REFERÊNCIAS ... 60
17
1 INTRODUÇÃO
1.1 PROBLEMA
Segundo Albert (2002), nos últimos anos nenhum outro aspecto de carga muscular (tensão específica aplicada à unidade músculo tendínea) tem sido mais descrito, estudado, discutido e investigado na literatura científica e prática clínica do que o movimento excêntrico. Garret Jr. (1996) afirma que muitos são os danos e/ou alterações musculares provocados pelas contrações excêntricas. Entretanto, este tipo de contração não é utilizado apenas em laboratório para testes, está presente nos movimentos diários, bem como nos frequentadores de academias e praticantes de esportes como basquete, futebol e rúgbi (CLEBIS e NATALI, 2001).
O exercício excêntrico é uma ação muscular dupla de alongamento e contração (força) simultânea, ocasionando efeitos deletérios acentuados em indivíduos treinados e destreinados (APPEL et al., 1992; LEE et al., 2002; LEE e CLARKSON, 2003). Esses efeitos deletérios ocasionam fadiga, dor muscular tardia, redução da força, inflamação e diminuição de performance (AVERY et al., 2003; CROSIER et al., 1996; KONNING et al., 2001; LI et al., 2002; PETERSEN et al., 2001; SHAFAT et al., 2004). Estes fatores possivelmente estão associados à formação de espécies reativas (ER), o que provavelmente dificulta o processo de recuperação e agrava ainda mais a diminuição de performance em atletas (BYER et al., 2006; CLEBIS e NATALI, 2001; PASCHALIS et al., 2006; STUPKA et al., 2000).
As lesões causadas pelos radicais livres nas células podem ser prevenidas ou reduzidas por meio da atividade de antioxidantes (KÖNIG e BERG, 2002). Os antioxidantes podem agir diretamente na neutralização da ação dos radicais livres ou participar indiretamente de sistemas enzimáticos com essa função (POWERS e LENNON, 1999).
O antioxidante denominado Taurina é um aminoácido não essencial sulfuroso, metabólico da metionina e com efeito antagonista ao da homocisteína, presente em alguns alimentos (CHANG, 2004; TANG, 2000; WILLIAMS, 2005). Apesar de não participar da estrutura de proteínas e enzimas (WILLIAMS, 2005), essa também tem importante função reguladora nos fluxos de cálcio, sódio e potássio e de proteção ao DNA (LOURENÇO e CAMILO, 2002), na estabilização da estrutura de membranas celulares (TANG, 2000) e na metabolização de gorduras (YOGOSHI et al., 1999). Além disso, a taurina vem demonstrando um poderoso efeito antioxidante em alguns estudos (DAWSON et al., 2002; ROIG-PÉREZ et al., 2004; ZEYBEK et al., 2006).
Por fim, considerando o pressuposto potencial deletério dos exercícios físicos excêntricos, a sua relação oxidante e inflamatória no músculo esquelético e a ação antioxidante da Taurina, formulou-se a seguinte questão a ser investigada neste estudo: quais são os efeitos da suplementação do antioxidante taurina sobre os parâmetros de estresse oxidativo muscular induzidos por exercício excêntrico?
1.2 JUSTIFICATIVA
A relação entre o estresse oxidativo e o exercício físico tem sido amplamente investigada nos últimos 30 anos (FISCHER-WELLMAN e BLOOMER, 2009). O interesse acerca dos mecanismos de geração e adaptação das ER ao exercício aumentou significativamente a partir da demonstração de sua relação com o consumo de oxigênio (CHILDS et al., 2001) e a inflamação no tecido muscular (GLEESON et al., 1998; KAURANEN et al., 2001; KÖNIG e BERG, 2002; MACINTYRE et al., 2000; MACINTYRE et al., 2001).
19
defesas antioxidantes andrógenas não conseguem neutralizar (ARMSTRONG et al., 1991; BALNAVE et al., 1993; CLEBIS e NATALI, 2001; MCBRIDE et al., 1998; SAXTON et al., 1994).
Além disso, o aumento das taxas metabólicas como resultado de exercício físico intenso pode aumentar excessivamente o consumo de oxigênio em até 20 vezes em relação aos valores de repouso (CARMELI et al., 2000). Esse aumento é seguido por um concomitante aumento na produção de radicais livres (ALESSIO e GOLDFARB,, 1998). Embora os benefícios do aumento no VO2máx sejam bem estabelecidos, um paradoxo bioquímico é verificado. O aumento no consumo máximo de oxigênio (O2) é essencial para a aptidão cardiovascular e performance, porém o aumento no consumo durante o exercício pode ser prejudicial em nível sérico e bioquímico (KÖNIG e BERG, 2002),gerando o estresse oxidativo.
No exercício excêntrico, o estresse oxidativo contribui para o desenvolvimento de lesão muscular (ARMSTRONG et al., 1991; BALNAVE et al., 1993; CLEBIS e NATALI, 2001; MCBRIDE et al., 1998; SAXTON et al., 1994). Este desequilíbrio bioquímico torna-se um poderoso comprometimento para o sistema muscular, pois as ER podem alterar os sistemas orgânicos por meio de sua ação tóxica.
O estresse oxidativo tem sido associado com a diminuição da performance, fadiga, dano muscular e excesso de treinamento - overtraining. Por essa razão, alguns pesquisadores (CARMELI et al., 2000; POLIDORI et al., 2000; POWERS et al., 1999; RÁDAK et al., 1999) sugerem que reduzir o estresse oxidativo pode melhorar a tolerância ao exercício, bem como a performance física. Os danos celulares ocasionados pelo estresse oxidativo podem ser revertidos pela ação de defesas antioxidantes (enzimáticas e não-enzimáticas), restabelecendo o equilíbrio entre a produção de ER e o sistema de defesa antioxidante (SILVA, 2006).
deletérios nas miofibrilas, após sessões de exercícios físicos em ratos e obteve resultados positivos.
Dessa forma, os achados desta pesquisa pretendem contribuir para o entendimento do mecanismo da ação antioxidante da taurina em exercícios físicos, especificamente em treinamento excêntrico. Assim, espera-se que esses resultados sejam de relevância para a área da Atividade Física e Saúde, objetivando a busca de benefícios entre a relação da suplementação e exercício físico.
1.3 OBJETIVOS
1.3.1 Objetivo Geral
Avaliar os efeitos da suplementação de taurina sobre os parâmetros de estresse oxidativo muscular após exercício excêntrico.
1.3.2 Objetivos Específicos
Verificar as respostas da suplementação de taurina sobre a produção de superóxido no músculo esquelético, após exercício excêntrico.
Verificar os efeitos da suplementação da taurina sobre os marcadores de danos oxidativos no músculo esquelético, após exercício excêntrico.
21
1.4 HIPÓTESES
1.4.1 Hipótese geral
A suplementação de taurina causará uma diminuição nos parâmetros de estresse oxidativo após uma sessão de exercício excêntrico em ratos.
1.4.2 Hipóteses Específicas
A produção de superóxido no músculo será atenuada com a suplementação de taurina.
Os marcadores de danos oxidativos no músculo serão atenuados com a suplementação de taurina.
A atividade enzimática no músculo será mantida com a suplementação de taurina.
1.5 DELIMITAÇÕES DO ESTUDO
A amostra foi composta por ratos Wistar machos, do biotério da Universidade do Extremo Sul de Santa Catarina (UNESC). O treinamento e as análises foram realizados nas dependências do Laboratório de Fisiologia e Bioquímica do Exercício (LAFIBE) da UNESC, localizado em Criciúma, SC.
1.6 LIMITAÇÕES DO ESTUDO
23
2 REFERENCIAL TEÓRICO
2.1 RADICAIS LIVRES
A definição de radical livre (RL) pode ser expressa como uma molécula altamente reativa ou um fragmento molecular que contém pelo menos um elétron ímpar em seu orbital externo. Tende a extrair elétrons de outras moléculas para alcançar um estado quimicamente mais estável (HALLIWEL e GUTTERIDGE, 2007).
Os radicais livres de oxigênio (RLO) são produzidos naturalmente no organismo durante a respiração e pelos processos metabólicos oxidativos para produção de energia (THOMAS, 2000). Quando se encontram em baixa a moderada concentração, eles exercem efeitos benéficos na defesa contra agentes infecciosos, em sistemas celulares de sinalização e na função mitogênica (VALKO et al., 2007). No entanto, um aumento desequilibrado na produção dessas moléculas pode lesionar componentes celulares indispensáveis para a vida da célula, tais como DNA, lipídios, proteínas e carboidratos (KÖNIG e BERG, 2002). Durante o exercício físico, a produção elevada dos RLO pode promover disfunção contrátil do músculo esquelético, resultando em fadiga muscular (POWERS e JACKSON, 2008).
2.2 PRINCIPAIS ESPÉCIES REATIVAS DE OXIGÊNIO
As espécies reativas (ER) são definidas como moléculas orgânicas altamente reativas e instáveis, podendo ter ou não um elétron desemparelhado na última camada de sua estrutura (HALLIWEL, 1994). O termo ERO não engloba somente as espécies reativas derivadas do oxigênio, mas também as RLO (POWERS e JACKSON, 2008). Apesar de existirem ER derivadas de outras moléculas orgânicas como as Espécies Reativas de Nitrogênio (ERN) e Espécies Reativas de Enxofre (ERX), as ERO estão em maior quantidade no organismo (FINAUD et al., 2006).
A maioria das ERO são produzidas na cadeia de transporte de elétrons mitocondrial durante a respiração sob condições fisiológicas normais. De acordo com
Halliwell e Gutterigde (2007), o oxigênio respirado é metabolizado sendo que aproximadamente 90 a 95% são utilizados pela mitocôndria, por meio da cadeia de transporte de elétrons e reduzidos a água durante a produção de energia. Na parte terminal da cadeia de transporte de elétrons, a enzima citocromo oxidase remove um elétron (e-) de cada uma das quatro moléculas reduzidas de citocromo C, oxidando-as e adiciona os quatro elétrons ao O2 para formar água (H2O) - Reação 1 (MATSUO e KANEKO, 2000).
Reação 1:
O2 + 4e- + 4H+→ 2 H2O + energia
De 2 a 5% do oxigênio respirado são reduzidos univalentemente em ERO (HALLIWEL e GUTTERIDGE, 2007). Durante o processo de formação de trifosfato de adenosina (ATP), o elétron da nicotinamida adenina dinucleotídeo reduzida (NADH) e da flavina adenina dinucleotídeo reduzida (FADH) é doado (via complexo I e II) para ubiquinona (CoQ) (complexo III) é reduzida formando a semiquinona (CoQH ) que em contato com o oxigênio forma o superóxido (O2-) – Conjunto de Reações 2 (HALLIWEL e GUTTERIDGE, 2007; NELSON e COX, 2007).
Reações 2:
CoQH2 + O2 CoQH + O2- CoQH + O2 CoQ + H+ + O2-
25
(THOMAS, 2000). Nos neutrófilos, a produção de O2- catalizada pela nicotinamida adenina dinucleotídeo fosfato (NADPH) oxigenase gera o burst respiratório necessário para para a destruição de bactérias (VALKO et al., 2006).
Reação 3:
2O2- + 2H+ H2O2 + O2
O H2O2 ao receber mais um elétron ou um íon de hidrogênio forma então o radical hidroxil (OH ), que é a ERO mais reativa dos intermediários, pois pode reagir e alterar qualquer estrutura celular que esteja próximo a ele, influenciando enzimas, lipídeos da membrana ou ácidos nucléicos (BIESALSKI, 2000; VANCINI et al., 2005). O radical hidroxil pode ser formado por mais duas reações conhecidas como reação de Fenton (FENTON, 1894) e Haber- Weiss (HABER, WEISS, 1934). Primeiro, o OH é formado quando o H2O2 reage com íons de ferro (Fe3+ e Fe2+ ) ou cobre (Cu3+ e Cu2+ ). Segundo, quando os íons de ferro e cobre catalisam a reação entre o O2- e o H2O2(Reações 4). Como características comuns, estas espécies apresentam grandes reatividades e possuem uma meia vida muito curta, o radical hidroxila, por exemplo, tem 10-9 segundos de vida, já o ânion superóxido decai a peróxido de hidrogênio em 4,5 x 105 milissegundos (SIES e CADENAS, 1989).
Reações 4:
O2- + H+ O2H O2H + O2- + H+ H2O2 + O2
Fe3+ + O2 - Fe2+ + O2
Fe2+ + H2O2 Fe3+ + OH + OH- (Reação de Fenton)
2.3 SISTEMA DE DEFESA ANTIOXIDANTE
Estas moléculas trabalham num complexo processo bioquímico e agem com o objetivo de prevenir a produção demasiada de radicais livres e reduzir um possível efeito deletério (BIESALSKI, 2000).
Os antioxidantes podem agir diretamente na neutralização da ação dos radicais livres ou participar indiretamente de sistemas enzimáticos com essa função (BARREIROS e DAVID, 2006). Especificamente, os antioxidantes podem atuar na prevenção da propagação de RL, na hidrólise enzimática de ligações estéricas para remoção de ácidos graxos peroxidados, no sequestro de íons metálicos e na redução catalítica-enzimática dos peróxidos (THOMAS, 2000).
O processo de neutralização dos ER por antioxidantes pode acontecer em três condições. A primeira previne a formação das substâncias agressoras. A segunda é a varredura, em que o antioxidante intercepta diretamente a ação da ER, neutralizando-o. A última condição é o reparo que ocorre quando as duas condições anteriores não foram bem sucedidas, e a ação do ER acaba desenvolvendo dano à estrutura celular. O antioxidante nesta condição tem a função de ajudar na recuperação da estrutura danificada (KONG e LILLEHEI, 1998).
Segundo Pinho (2005), as defesas antioxidantes podem atuar de forma associada ou independente por duas vias: enzimáticas; sistema composto pelas enzimas superóxido dismutase (SOD), a catalase (CAT) e a glutationa peroxidase (GPX) as quais são ativadas normalmente durante o metabolismo celular, porém suas atividades podem aumentar em função da presença de ER; e não enzimáticas, incluem as vitaminas E, C e betacaroteno, glutationa (GSH), taurina entre outros. Grande parte dos antioxidantes não enzimáticos é encontrada na alimentação e eles podem ser suplementados por uma dieta alimentar.
2.3.1 Antioxidantes enzimáticos
27
em condições fisiológicas e sob exercício físico leve (BANERJEE et al., 2003). As principais enzimas antioxidantes estão apresentadas na tabela 1 abaixo.
Tabela 1 – Principais antioxidantes enzimáticos (Adaptado de: POWERS e JACKSON, 2008)
Antioxidantes
enzimáticos Localização Função
Superóxido
Dismutase (SOD) Matriz mitocondrial, citosol e meio extracelular Scavenger de O2
-Catalase (CAT) Citosol, principalmente dos
peroxissomos Scavenger de H2O2
Glutationa
Peroxidase (GPX) Citosol, membrana mitocôndria, e meio extracelular
Scavenger de H2O2
2.3.1.1 Superóxido Dismutase (SOD)
A SOD constitui a primeira linha de defesa enzimática contra a produção intracelular de radicais livres, catalisando a dismutação do O2- (HOLLANDER et al., 2000; MOOREN e VÖLKER, 2004). Embora o O2- não seja altamente danoso, é um precursor do OH e pode extrair elétrons de alguns componentes celulares, causando reações em cadeia de radicais livres (HALLIWELL e GUTTERIDE, 2007).
O produto resultante da reação catalisada pela SOD é o H2O2 (Reação 5) que deve ser retirado do meio o mais rápido possível. É estimado que mais de 80% de O2 -formado na mitocôndria seja reduzido pela SOD (MOOREN e VÖLKER, 2004).
Reação 5:
2.3.1.2 Catalase (CAT)
A enzima CAT catalisa a degradação do H2O2, é uma hemeproteína (PRYOR e GODBER, 1991) e está localizada principalmente no peroxissoma celular (FINAUD et al., 2006). Também está presente em outras organelas como as mitocôndrias e retículo endoplasmático (MOOREN e VÖLKER, 2004). Na reação, uma das moléculas de peróxido de hidrogênio é oxidada a oxigênio molecular e a outra é reduzida à água –
Reação 6 (CHANGE et al., 1979).
Reação 6:
2 H2O2 2 H2O + O2
A CAT pode também usar o H2O2 para reagir com alguma substância tóxica, via reação catalisada pela peroxidase (Reação 7). Esta reação necessita de um substrato como fenol, álcool (A) ou ácido fórmico para reagir com H2O2 (FINAUD et al., 2006).
H2O2 + H2A (substrato) 2 H2O + A (Reação 7)
A catálise do H2O2 é muito importante, pois na presença de Fe2+ ou Cu2+ leva à formação de radical hidroxil (OH ) (reação de Fenton- Conjunto de reações 4) (CHANGEet al., 1979).
2.3.1.3 Glutationa Peroxidase (GPX)
29
Reação 8:
H2O2 +2 GSH GSSH (glutationa oxidada) + 2 H2O
2.3.2 Antioxidantes não enzimáticos
Incluem as substâncias encontradas nos alimentos como as vitaminas (C e E), minerais (zinco, selênio), provitaminas (alguns carotenóides), aminoácidos (taurina), flavonóides e polifenóis conforme mostra a tabela 2. Substâncias orgânicas como proteínas com grupo tiol (Glutationa - GSH), e várias outras substâncias de baixo peso molecular orgânicas como a ubiquinona e o ácido úrico também têm ação antioxidante e consideradas defesas exógenas (KÖNIG e BERG, 2002; BIESEK et al., 2005).
Tabela 2 – Principais antioxidantes não-enzimáticos (Fonte: KÖNIG e BERG, 2002) Antioxidantes
não-enzimáticos Localização Função
Vitamina C Ambiente aquoso da
célula Scavenger Recicla Vit. E de RL
Vitamina E Membranas
celulares lipoperoxidação Impede a Reduz vários ERO para a forma menos reativa
Carotenóides Membranas
celulares Scavenger Protege de RL da liperoxidação
Glutationa Em todo o ambiente
celular Scavenger Remove de RL H+ e peróxidos orgânicos na reação catalizada por GPX
Recicla Vit. C e E.
Flavonóides/
Polifenóis celulares Membranas Scavenger Quelador de metais de RL
Ubiquinonas Membranas
Alguns estudos mostram que a suplementação de alguns antioxidantes não-enzimáticos diminuiu o dano oxidativo em diferentes tecidos e doenças. Pinho et al. (2005) observou que a administração de N-Acetilcisteína (NAC) reduziu a resposta inflamatória e os parâmetros de estresse oxidativo pulmonar em ratos expostos à poeira de carvão. No estudo de Jain et al. (2009), os autores suplementaram vitamina C e E em coelhos diabéticos e verificaram uma diminuição no dano do DNA plasmático. Ramirez-Tortosa et al. (2008) comprovou que a suplementação de coenzima Q 10 reduziu o estresse oxidativo em mitocôndrias do fígado de coelhos com aterosclerose.
No exercício físico, várias pesquisas demonstram os efeitos positivos de alguns antioxidantes. Silva et al. (2008) demonstrou que o tratamento com NAC pode ter alguns efeitos anti-inflamatórios e age sobre a regulação de citocinas pró-inflamatórias após o exercício excêntrico. Araújo (2008) suplementou licopeno aos pacientes com Doença Arterial Coronariana e confirmou que a diminuição do dano oxidativo ocorreu naqueles pacientes que estavam submetidos a um treinamento de exercício físico intenso e controlado. No estudo em que Sureda et al. (2008) suplementou vitamina C e E em corredores amadores e a suplementação com níveis moderados desses antioxidantes reduziu o dano oxidativo, sem bloquear a adaptação celular ao exercício. Já Zoppi et al. (2006) verificou que a suplementação dessas duas vitaminas em jogadores profissionais de futebol pode reduzir a peroxidação lipídica e o dano muscular durante o período de treinamento mas não tem nenhum efeito sobre a performance.
2.4 TAURINA
31
descarboxilase para sintetizar a hipotaurina que é oxidada à taurina (BOUCKENOOGHE et al., 2006).
É um dos aminoácidos intracelulares mais abundantes do organismo e não faz parte da estrutura de proteínas. Uma pequena quantidade está presente nos peptídoes do cérebro (LOURENÇO e CAMILO, 2002). Nos tecidos, encontra-se uma maior concentração de taurina nos músculos e no cérebro (GOODMAN et al., 2009). Faz parte da estrutura do ácido taurocólico, um ácido biliático, produzido no fígado que ajuda na digestão de lipídeos (NELSON e COX, 2007).
A taurina faz parte de vários processos metabólicos como contração cardíaca, atividade anti-oxidante, permeabilidade e proteção da membrana e contração muscular (WILLIAMS, 2005). Cuisinier et al. (2000) relatou em um estudo que o miofilamento cálcio-sensível e as características mecânicas são dependentes da taurina. Ela pode controlar a homeostase do íon cálcio (Ca2+) intracelular e ajudar no controle de excitação-contração muscular (CONTE-CAMERINO et al., 2004).
A suplementação de taurina tem mostrado em estudos benefícios à saúde. Nonaka et al. (2001) relatou que um maior consumo de taurina pode prevenir a progressão de aterosclerose e isquemia cardíaca.
Durante o exercício, Zhang (2004) reportou que sete dias de suplementação de taurina induziram significantemente o aumento do VO2max e o tempo para chegar à exaustão na pedalada. O autor sugeriu que os efeitos ergogênicos foram ocasionados pela propriedade antioxidante da taurina e proteção das membranas celulares. No estudo feito por Galloway et al. (2008), o mesmo tempo de suplementação de taurina não alterou a concentração de carboidratos e oxidação de lipídeos no músculo esquelético, mas causou impacto na metabolização, metabolismo ou transporte de indispensáveis aminoácidos no músculo durante exercício prolongado.
2.4.1 Regulação dos canais de cálcio
A taurina pode diminuir o dano oxidativo, regulando os Ca2+ que estão envolvidos na patogênese do dano celular mediados por RL na rota da xantina oxidase (XO) (Chang et al., 2004).
Em um estudo em que os autores examinaram as propriedades contráteis do músculo esquelético de ratos e a quantidade de taurina presente no músculo, observou-se a redução de força no grupo que apreobservou-sentou o nível de taurina muscular inferior ao grupo controle. Os autores atribuíram esta situação à relação que a taurina tem com o controle excitação-contração muscular que influencia na liberação de Ca2+ celular e na produção de força (HAMILTON et al., 2006). Os resultados alcançados por Bakker e Berg (2001) mostram que a taurina é um modulador da contração do músculo esquelético pelo aumento do acúmulo do Ca2+ do retículo sarcoplasmático e liberação de Ca2+ da célula.
Pelo ponto de vista farmacológico, a suplementação de Taurina pode ser benéfica, pois esse aminoácido pode diminuir a hipercalemia observada durante a reperfusão pelo bloqueio de canais de potássio dependentes do cálcio (KCa2+). Este processo pode estar relacionado com a produção demasiada de moléculas reativas de óxido nítrico e consequentemente, estresse oxidativo (CONTE-CAMERINO et al., 2004).
2.4.2 Equilíbrio osmótico celular
33
ativação do canal KATP possa ser a chave do mecanismo para a adaptação para a hipóxia tecidual, apoptose celular e proteção cardíaca contra a isquemia (MURZAEVA et al., 2008).
Pode existir, ainda, uma relação entre a atividade da Aquaporin-4 (canal de água da célula) e o conteúdo de taurina muscular no momento em que há a necessidade de estabelecer um equilíbrio de água para amenizar os efeitos adversos do lactato formado durante o exercício anaeróbio (CONTE-CAMERINO et al., 2004).
2.4.3 Neutralização direta de ERO
A taurina está presente abundantemente em células fagocitárias (KIM e CHA, 2008) e age como um scavenger de HClO gerado endogenamente, que constitui uma molécula muito tóxica ao organismo. A taurina cloramina é um aldeído estável e não tóxico formada por meio da reação entre a taurina e HClO e age como uma molécula sinalizadora do decréscimo do processo de alguns mediadores como óxido nítrico
(NOS), fator de necrose tumoral α (TNF-α), prostaglandina 2 (PGE2) e enzima ciclooxigenase 2 (COX-2) envolvidos em reações inflamatórias (QUINN, 1998).
2.5 EXERCÍCIO FÍSICO E ESTRESSE OXIDATIVO
Os efeitos benéficos da atividade física já são bem conhecidos. Em indivíduos saudáveis a prática regular de exercício promove um aumento na aptidão física e previne várias doenças como cardiopatias, obesidade e diabetes. O aumento do VO2max em atletas, a melhora a resistência à insulina em pacientes diabéticos e a diminuição da morbidade ocasionada pelo infarto agudo do miocárdio são alguns efeitos favoráveis do treinamento físico verificado em alguns estudos (FAISAL et al., 2009; PRAET et al., 2007; RISTOW et al., 2009).
O estresse oxidativo tem sido associado a diminuição da performance, fadiga, dano muscular e overtraining (CARMELI et al., 2000). Por essa razão, alguns pesquisadores sugerem que reduzir o estresse oxidativo pode melhorar a tolerância ao exercício físico bem como a performance (ALESSIO et al., 1999).
Embora os benefícios do aumento no VO2 máx sejam bem estabelecidos, um paradoxo bioquímico é verificado. O aumento no consumo de O2 é essencial para aptidão cardiovascular e performance, porém o aumento no consumo durante ou após o exercício pode ser prejudicial, quando é excedida a capacidade normal do individuo (PINHO, 2005).
A geração de ERO durante o exercício depende do tipo (aeróbia ou anaeróbia), intensidade, duração, exigência de energia, os níveis de consumo de oxigênio e tensões mecânicas impostas sobre os tecidos (FISCHER-WELLMAN, 2009). O exercício físico intenso provoca um aumento de 10 a 20 vezes no consumo total de oxigênio do organismo e um aumento de 100 a 200 vezes na captação de oxigênio pelo tecido muscular, favorecendo o aumento na produção de ERO (SAXTON et al., 1994). Entretanto, estudos têm demonstrado que o treinamento de endurance aumenta as defesas antioxidantes, assim como a capacidade oxidativa do músculo (PINHO et al., 2006; RÁDAK et al., 1999; TERBLANCHE, 2000).
35
BERG, 2002). Esses processos geram um acúmulo de ERO em algumas situações que podem ocasionar prejuízos relevantes na performance.
2.5.1 Formação de ERO durante o metabolismo do oxigênio na cadeia respiratória
A fosforilação oxidativa resulta na produção de ATP na mitocôndria em situações normais. A oxidação do substrato ocorre no ciclo de krebs e na cadeia transportadora de elétrons com o oxigênio como receptor de elétrons. Na cadeia respiratória, 95-99% do oxigênio consumido é reduzido em água por redução tetravalente (reação 1) catalizada pela CoQ (FINAUD et al., 2006).
Entretanto, 1-5% de O2 do fluxo da cadeia tranportadora de elétrons são desviados para a formação de superóxido (FINAUD et al., 2006). No complexo I, a principal rota de fuga para o oxigênio é reagir com o ferro e o enxofre encontrados no local e no complexo III. O produto formado pelas duas reações é o superóxido, sendo que o complexo III libera-o por ambos os lados do interior da membrana mitocondrial. Não está claro se este superóxido atravessa a membrana externa mitocondrial ou é dismutado por Cu / Zn-SOD localizada na mitocôndria localizada no espaço inter-membrana. Durante o exercício, a geração de ERO que ocorre durante a atividade contrátil está diretamente relacionada com o elevado consumo de oxigênio que ocorre com o aumento da atividade mitocondrial (POWERS e JACKSON, 2007).
2.5.2 Formação de ERO durante isquemia-reperfusão
A alta concentração de Ca2+ em situações isquêmicas tem sido associada com outra via de produção de ERO. O acréscimo das concentrações de Ca2+ pode ativar a enzima fosfolipase A2 a qual libera o ácido araquidônio a partir dos fosfolipídeos. Zuo et al. (2004) concluiu que o metabolismo do ácido araquidônico, em associação com a atividade da lipoxigenase, é a maior fonte de O2- no músculo esquelético.
Exercícios intensos podem aumentar a produção de ERO devido à hipóxia e reoxigenação temporárias que ocorrem no músculo exercitado em função de contrações e relaxamentos estabelecidos ciclicamente. Durante a contração, a compressão vascular estabelece um quadro de isquemia, gerando uma hipóxia. Entretanto no relaxamento, acontece a reperfusão, e, consequentemente, a reoxigenação (HALLIWELL E GUTTERIDGE, 2007).
Sob condições aeróbias, o oxigênio suficiente assegura que o ATP seja reposto primeiramente via fosforilação oxidativa mitocondrial e que a hipoxantina/xantina sejam convertidas para ácido úrico pela xantina desidrogenase do que pela xantina oxidase. Além disso, o músculo esquelético tem baixa atividade da XO. Todavia a XO pode ser um importante caminho quando o músculo apresentar um déficit de adenina dinucletídeo. Essa situação teoricamente pode acontecer em situação isquêmica, exercício isométrico, sprint, déficit de O2, exercícios com limitação vascular de fluxo sanguíneo (BEJMA e JI, 1999, CHEVION et al., 2003).
37
2.5.3 Formação de ERO durante processo inflamatório
Danos musculares provocados por exercícios levam a migração de várias células do sistema de defesa como leucócitos, neutrófilos, monócitos e macrófagos que são capazes de produzir ERO (MOOREN e VÖLKER, 2004). A ativação de leucócitos após o dano muscular, induzido pelo exercício físico, pode estimular a produção de RL para atacar os antígenos invasores. Em particular os neutrófilos são as maiores fontes de produção de O2- pela reação NADPH-oxidase (Reação 9). Na presença de H2O2 e íon clorido, os neutrófilos geram o potente HClO via reação catalizada pela enzima mieloperoxidase. A netrofilia induzida pelo exercício ocorre como resultado da desmarginação de neutrófilos de tecidos endoteliais (mediados por catecolaminas) e medula óssea (mediado pelo cortisol) (TREVOR E SANDY, 2001). A produção demasiada de ERO responsáveis pelo ataque ao agente invasor no processo inflamatório é chamada de burst oxidativo e é responsável pela primeira linha de defesa contra o desenvolvimento de patogenias (DRÖGE, 2002).
Reação 9:
2 O2 + NADPH 2 O2- + NADP+ + H+
Uma concentração relevante de ERO pode modular o sinal sensível-redox para aumentar as funções imunológicas dos linfócitos (DRÖGE, 2002). Embora isso seja uma reação de proteção do organismo, pode ser causa de inflamações agudas, devido à produção de mediadores pro-inflamatórios adicional como interleucinas 1,6 e 8, TNF-
α e PGE, levando a produção de processo inflamatório adicional e aumentando a produção de ERO (MASTALOUDIS et al., 2001).
2.6 RELAÇÃO ENTRE EXERCÍCIO EXCÊNTRICO E ESTRESSE OXIDATIVO
de ERO (FINAUD et al., 2006). O ferro liberado neste processo da hemoglobina e ferritina pode ser fator preponderante na formação adicional de ERO, pois o ciclo redox do ferro promove a reação de Fenton e Haber-Weiss (Conjunto de Reações 4) as quais resultam na produção de radical hidroxila. Esse radical livre é altamente danoso, é capaz de retirar um átomo de hidrogênio dos ácidos graxos poliinsaturados (PUFA) da membrana celular e iniciar a peroxidação lipídica (WELCH et al.,2002).
Estudos em animais (DUAN et al., 1990; WARREN et al., 1996) têm mostrado que após a prática de exercícios excêntricos acontece um acúmulo de Ca+2 mitocondrial, produzindo uma queda no potencial da membrana mitocondrial para a produção de ATP e, consequentemente, diminuição das contrações musculares. Isto pode ocorrer pois o exercício excêntrico pode desequilibrar a homeostase do cálcio devido à excessiva atividade de contração e lesão muscular e isquemia tecidual. Esse conjunto de processos pode aumentar a geração de ERO pela via xantina-oxidase (KÖNIG e BERG, 2002).
2.7 LESÕES NO EXERCÍCIO EXCÊNTRICO
A lesão muscular é causada por exercícios físicos extenuantes em músculos não habituados a receberem certos estímulos, principalmente quando envolvem contrações musculares excêntricas quando os músculos são alongados enquanto exercem força (CLARKSON e SAYERS, 1999). No exercício excêntrico, as lesões ocorrem porque as ações de alongamento provocam uma extensão além do normal de alguns sarcômeros, causando danos a essas estruturas (CLEBIS e NATALI, 2001). Um exemplo de exercício excêntrico é a corrida em declive, em que na descida a contração do músculo do quadríceps sofre o processo de alongamento a fim de controlar o grau de flexão do joelho contra a força da gravidade (PROSKE e MORGAN, 2001).
39
al. (2006) afirmam que as lesões ocorridas pela ação excêntrica podem causar diminuição do alongamento do músculo, dor muscular tardia e inflamação do músculo.
Em um de seus estudos com treinamento excêntrico em humanos, Sorichter et al. (2006) evidenciou que a máxima força muscular associada à ação excêntrica causa a rápida dissociação e/ou degradação e remoção imediata de troponina do tipo I. Dessa forma, são possíveis alterações no complexo do filamento da troponina imediatamente após a lesão muscular, induzida por exercício físico.
Além disso, mudanças na função muscular a nível celular foram observadas após o exercício excêntrico. Ocorre uma diminuição na quantidade de proteínas transportadora de glicose e lactato/ H+ e sua ação torna-se prejudicada (CLARKSON e SAYERS, 1999). Estudos mostram a elevação de creatina quinase e lactato desidrogenase por vários dias, após treinamento excêntrico (DONNELY et al., 1992; ESTON et al., 1996; NOSAKA e CLARKSON, 1994), confirmando assim a ação deletéria do exercício excêntrico nas fibras musculares.
Os exercícios excêntricos induzem a dor muscular tardia (DMT) (GLEESON et al., 1997). Funciona como um mecanismo de proteção que age para diminuir a atividade muscular e prevenir lesões mais graves (ZAINUDDIN et al., 2006). Caracterizada pela sensibilidade local ou generalizada nos músculos estressados, manifesta-se depois de oito horas do término do exercício e progride de intensidade nas primeiras 24 horas, alcançando seu pico entre 48 e 72 horas (TRICOLI, 2001). Gleeson et al. (1997) afirma que a DMT inicia entre seis e dez horas após o exercício excêntrico e o pico acontece entre 24 e 48 horas depois do exercício.
3 MÉTODOS
Todos os procedimentos foram aprovados pelo Comitê de Ética no Uso de Animais da Universidade do Extremo Sul de Santa Catarina (UNESC) – Brasil e realizados conforme a Lei nº 11.794/08 (DOU 196, Seção 1, Outubro/2008).
3.1 CARACTERIZAÇÃO DA PESQUISA
Esta pesquisa caracteriza-se por ser do tipo experimental com delineamento experimental (THOMAS e NELSON, 2002), pois se trata de comparar a resposta da suplementação do antioxidante em grupo experimental e controle.
3.2 AMOSTRA
41
Tabela 3: Composição básica do produto (ração Nuvilab® CR-1) / kg do produto
Valor Nutricional da dieta padrão % por kg do produto
Umidade (máx.) 12,5
Proteína Bruta (min.) 22 Extrato Étereo (min.) 4,5 Matéria mineral (máx.) 10
Matéria fibrosa (máx.) 8
Cálcio (máx.) 1,4
Fósforo 0,8
3.3 TRATAMENTO EXPERIMENTAL
Os animais foram divididos em quatro grupos (n=6): controle (C); exercício excêntrico (EE); exercício excêntrico + taurina (EET) e exercício excêntrico + salina (EES).
3.4 PROTOCOLO DE ADAPTAÇÃO AO EXERCÍCIO
Todos os animais foram ambientados em esteira ergométrica sem estimulação elétrica durante uma semana, antes da sessão de exercício excêntrico (10 minutos de corrida), com velocidade constante (10 m.min-1) e sem inclinação.
3.5 PROTOCOLO DE EXERCÍCIO EXCÊNTRICO DE LONGA DURAÇÃO
sessão de treinamento de adaptação. A intensidade do exercício realizado pode ser considerada moderada (+ 60% do VO2 máx.) de acordo com Brooks e White (1978).
3.6 SUPLEMENTAÇÃO
A taurina sintética (marca Sigma®) foi administrada neste estudo de acordo com o experimento de Myasaki et al. (2004). A taurina foi manipulada por gavagem (300mg/kg de peso corporal diluído em 1ml de água deionizada) em dose única diária por um período de 14 dias consecutivos antes da sessão de exercício e 2, 12, 24 e 48 horas depois dela. Foi utilizada 1ml de salina (NaCl 0,9%) como veículo.
3.7 EUTANASIA
43
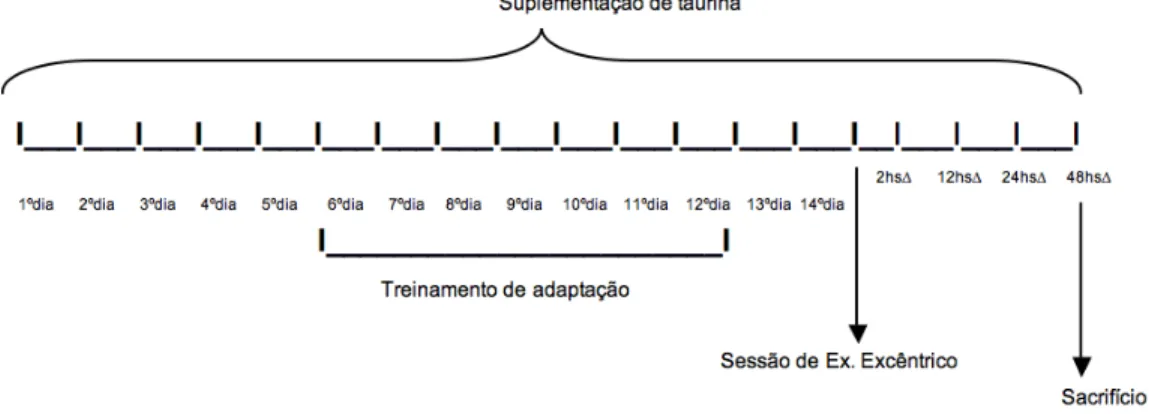
3.8 DESENHO DO ESTUDO
Figura 1 – Desenho do estudo “Efeitos da suplementação de taurina sobre os parâmetros de estresse oxidativo induzidos pelo exercício excêntrico”.
3.9 ENSAIOS BIOQUÍMICOS
3.9.1 Produção de Espécies Reativas de Oxigênio
3.9.1.1 Ânion Superóxido
O superóxido é determinado pela oxidação da adrenalina em tampão contendo SMP, succinato (inibidor da cadeia de transferência de elétrons) e catalase. Um controle negativo é determinado com a presença de SOD (PODEROSO et al., 1996).
tampão lavagem à amostra. Centrifugou-se por mais 10 minutos e o pellet foi ressuspendido. Esta última fase foi realizada 2 vezes antes de dosar.
3.9.2 Marcadores de Danos Oxidativos
3.9.2.1 Espécies Reativas ao Ácido Tiobarbitúrico (TBARS)
Este método foi utilizado para a avaliação do estado de oxidação dos ácidos graxos em sistemas biológicos. O dano em lipídeos da membrana é determinado pela formação de subprodutos da lipoperoxidação (malondialdeído-MDA), que são substâncias reativas ao aquecimento do ácido tiobarbitúrico (TBA), formadas durante a peroxidação em sistemas de membranas e microssomos. O MDA reage com o TBA gerando um produto róseo lido em espectrofotômetro (DRAPER e HADLEE, 1990).
Esta análise foi realizada conforme descrito por Draper e Hadlee (1990). As amostras foram homogeinizadas em tampão TBA, lisadas em TCA 5% e centrifugadas por 10 min. O sobrenadante foi fervido por 30 min em TBA 67%. Após o resfriamento em temperatura ambiente, as substâncias reativas ao ácido tiobarbitúrico foram quantificadas a 532 nm.
3.9.2.2 Carbonilação de proteínas
Este método foi utilizado para dosar a oxidação de proteínas. Baseia-se no princípio de que vários RL atacam resíduos de proteínas como aminoácidos (principalmente histidina, arginina, lisina e prolina) resultando em produtos com o grupo carbonil, o qual pode ser medido pela reação com dinitrofenihidrazina (LEVINE et al., 1990).
45
HCl (1:3) adicionado de 10 mM de DNPT. Após o período de incubação, adicionou-se 100 L de TCA 20% e as amostras foram centrifugadas a 14000g por três minutos. O pellet foi lavado com 500 L de etanol-etilacetato. As amostras foram incubadas por mais 30 minutos a 60°C, centrifugadas novamente a 14000g por 3 minutos. O conteúdo de formação de carbonil foi determinado espectrofotometricamente em 370nm, usando um coeficiente 22.0000 molar-1.
3.9.2.3 Sulfidrilas
O ácido ditionitrobenzóico (DTNB) é reduzido por tióis gerando um derivado amarelo (TNB) e lido espectrofotometricamente a 412 nm. Este método determina os tióis totais da amostra, sendo um parâmetro de medida de dano oxidativo às proteínas (AKSENOV e MARKESBERY, 2001).
As amostras do quadríceps foram homogeneizadas em 1 ml de tampão PBS (7.4 pH) com 1 mM de EDTA. Logo após, foi adicionado a amostra DTNB e incubada por 30 minutos à temperatura ambiente no escuro. Por fim, as amostras foram lidas espectrofotometricamente.
3.9.3 Atividade de enzimas antioxidantes
3.9.3.1 Superóxido dismutase (SOD)
O produto resultante da reação catalisada pela SOD é o H2O2 que deve ser retirado do meio o mais rápido possível. Uma unidade de enzima é definida pela quantidade transformada em 1µmol de substrato por minuto. A atividade enzimática foi determinada pela inibição da auto-oxidação da adrenalina medida espectrofotometricamente (480nm), segundo Bannister e Calabrese (1987).
leitura, adicionou-se 10 µl de catalase e 970 µl de tampão glicina à quantidade de amostra (10, 20 ou 30 µl) e logo depois zerou-se o espectro. Foi adicionado 17 µl de adrenalina e leu-se novamente em 180 segundos.
3.9.3.2 Catalase (CAT)
Esta análise mediu a atividade da enzima produzida pelas células e organelas da fração retirada do quadríceps em resposta a quantidade de peróxido de hidrogênio determinada pela queda na absorbância (240nm), conforme previamente descrito por Aebi (1984).
O procedimento iniciou com a homogeinização das amostras em tampão fosfato. Foram centrifugadas em 3000g por 10 minutos. Foram pipetadas 50µl de amostra e 1ml de tampão fosfato em cubeta de quartzo. Logo depois foi zerado o espectro. Foi utilizado o tampão fosfato com peróxido de hidrogênio para fazer a leitura. Foi adicionado 1ml de peróxido de hidrogênio à 50µl de cada amostra e feita a leitura nos tempos de 0 segundos, 30 segundo e 60 segundos.
3.9.4 Determinação da Proteína
A quantidade de proteínas de todos os ensaios foram mensurados usando a técnica de LOWRY et al. (1951). O princípio do método baseia-se em uma mistura contendo molibdato, tungstato e ácido fosfórico, reagente Folin-Ciocalteau, que sofre uma redução quando reage com proteínas, na presença do catalisador cobre (II) e produz um composto com absorção máxima em 750 nm.
47
3.10 CÁLCULO E TAMANHO DA AMOSTRA
Baseando-se em Silva et al. (2009) em que uma relação média (± Erro padrão médio - EPM) de TBARS de 0,14 ± 0,07nmol/mg proteína antes do protocolo de exercício excêntrico e de 0,24 ± 0,08nmol/mg proteína após a sessão de exercício, no presente estudo o tamanho do grupo amostral necessário para detectar uma diferença significativa, considerando um nível de significância de 5% e poder de 80%, foi de 6 animais por grupo totalizando 24 animais em todo experimento.
3.11 TRATAMENTO ESTATÍSTICO
4 RESULTADOS
4.1 PRODUÇÃO DE ESPÉCIES REATIVAS DE OXIGÊNIO
4.1.1 Ânion Superóxido
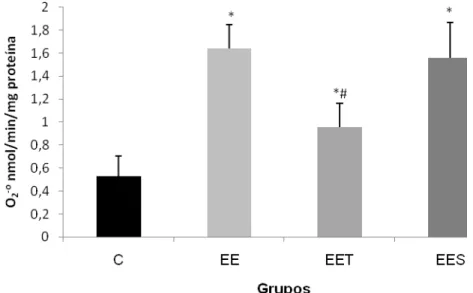
Houve um aumento significativo na produção de superóxido após a sessão de exercício excêntrico em todos os grupos em relação ao grupo controle - C (0,52 0,17 nmol/min/mg proteína). O grupo EET apresentou uma redução na produção do ânion Superóxido (0,9 0,2 nmol/min/mg proteína) quando comparado com o grupo EE (1,62 0,2 nmol/min/mg proteína) e o grupo EES (1,55 0,3 nmol/min/mg proteína) -Figura 2.
49
4.2 MARCADORES DE DANOS OXIDATIVOS
4.2.1 Espécies Reativas ao Ácido Tiobarbitúrico (TBARS)
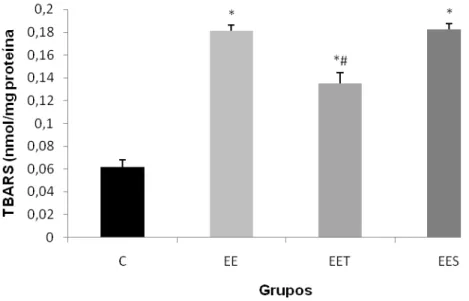
Na figura 3, a quantidade de TBARS aumentou em todos os grupos em comparação ao C (0,06 0,006 nmol/mg proteína). No entano, EET apresentou quantidade de TBARS menor (0,13 0,001nmol/mg proteína) em relação a ambos os grupos (EES - 0,18 0,03 nmol/mg proteína e EE - 0,18 0,05 nmol/mg proteína).
4.2.2 Conteúdo total de tióis
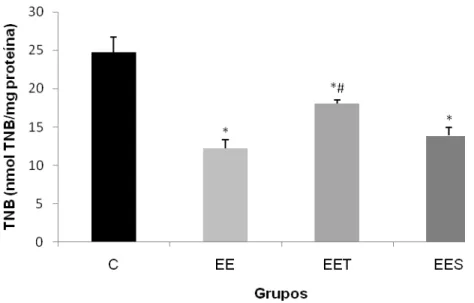
Ocorreu uma diminuição no conteúdo total de tióis em todos os grupos após a sessão de exercício excêntrico em relação ao grupo C (Figura 4). Entretanto, o grupo EET apresentou um maior conteúdo total de tióis (18,8 0,3 nmol TNB/mg proteína) em comparação com os demais grupos (EES -13,8 1 nmol TNB/mg proteína; EE - 12,2 1 nmol TNB/mg proteína).
Figura 4: Conteúdo total de tióis no músculo do quadríceps de ratos, após uma sessão de exercício excêntrico: grupo controle (C), grupo exercício excêntrico (EE), grupo exercício excêntrico com suplementação de taurina (EET) e grupo exercício excêntrico com placebo (EES). Os valores estão representados em média+EPM e os resultados expressos em nmol TNB/mg proteína. Diferenças significativas em relação ao grupo controle (*) e em relação ao EES (#) com nível de significância p<0,05.
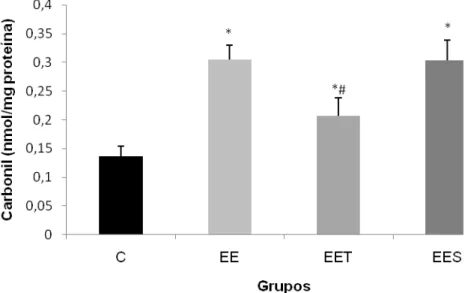
4.2.3 Carbonilação de proteínas
51
0,2 nmol/mg proteína) quando comparado com ambos os grupos (EES- 0,30 0,03 nmol/mg proteína; e EE - 0,30 0,025 nmol/mg proteína).
Figura 5: Níveis de carbonilação de proteínas no músculo do quadríceps de ratos, após uma sessão de exercício excêntrico: grupo controle (C), grupo exercício excêntrico (EE), grupo exercício excêntrico com suplementação de taurina (EET) e grupo exercício excêntrico com placebo (EES). Os valores estão representados em média + EPM e os resultados expressos em nmol /mg proteína. Diferenças significativas em relação ao grupo controle (*) e em relação ao EES (#) com nível de significância p<0,05.
4.3 ATIVIDADES DE ENZIMAS ANTIOXIDANTES
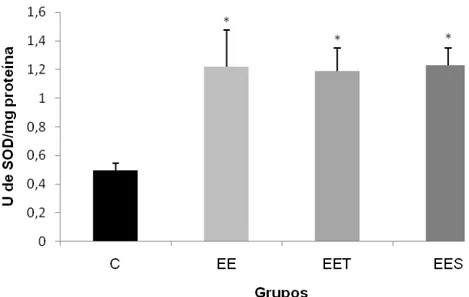
4.3.1 Superóxido dismutase (SOD)
Figura 6: Atividade da enzima Superóxido Dismutase (SOD) no músculo do quadríceps de ratos, após uma sessão de exercício excêntrico: grupo controle (C), grupo exercício excêntrico (EE), grupo exercício excêntrico com suplementação de taurina (EET) e grupo exercício excêntrico com placebo (EES). Os valores estão representados em média + EPM e os resultados expressos em U de SOD /mg proteína. Diferenças significativas em relação ao grupo controle (*) e em relação ao EES (#) com nível de significância p<0,05.
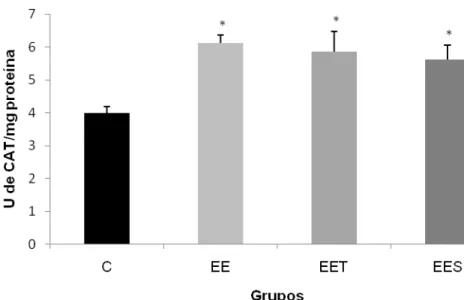
4.3.2 Catalase (CAT)
53
5 DISCUSSÃO
Neste estudo foi possível observar que uma sessão de exercício excêntrico ocasionou um aumento na produção de ERO (representado pelo ânion superóxido), no dano oxidativo de lipídeos e proteínas e na atividade das enzimas antioxidantes (SOD e CAT). Corroborando com outros autores (BLOOMER et al., 2004; CHILDS et al., 2001; SILVA et al., 2008), é possível confirmar que este tipo de exercício provoca um desequilíbrio no sistema oxidante/antioxidante, causando o estresse oxidativo.
Com o objetivo de atenuar os efeitos deletérios ocasionados pelo exercício excêntrico, utilizou-se a suplementação de taurina como recurso antioxidante. A quantidade suplementada de taurina neste trabalho foi adaptada de acordo com Miyazaki et al. (2004) que encontrou efeitos positivos na performance de ratos, com doses entre 100 e 500mg de taurina /kg de peso corporal.
Diversos estudos mostram que o exercício excêntrico provoca alterações nos marcadores de estresse oxidativo em um período entre 24 e 72 horas pós-exercício (GOLDFARB et al., 2005; LEE et al., 2002, SILVA et al., 2008). Neste estudo optou-se por avaliar os parâmetros de estresse oxidativo 48 horas depois de uma sessão de exercício excêntrico.
A geração de ânion superóxido, durante o exercício físico, ocorre por diversas vias, sendo que algumas delas são: o aumento no fluxo de elétrons na cadeia transportadora, ativação da xantina-oxidase e NADPH-oxidase durante e depois das contrações excêntricas (HALLIWEL e CROSS, 1994; ZUO et al., 2004; BUTTERFIELD et al., 2006).
55
Este efeito antioxidante da taurina pode ser atribuído à capacidade que ela tem de se ligar ao HClO e gerar um produto menos tóxico, a taurina cloramina (TauCl) que possui a capacidade de neutralizar o ânion superóxido e o óxido nítrico (KIM e CHA, 2009). A TauCl pode ainda inibir as ações do fator nuclear kappa B (NFĸB), um fator de transcrição que participa da resposta inflamatória (GURUJEYALASHMI et al., 2000) e pode bloquear a produção de quimiocinas nos macrófagos alveolares que estão envolvidos no recrutamento de neutrófilos (LIU e QUINN, 2002; ZEYBEK et al., 2006) diminuindo o processo inflamatório. Outra possibilidade, segundo Hansen et al. (2006) é que a taurina pode atuar como antioxidante em locais com alta produção de ERO, incluindo a mitocôndria, corroborando com a hipótese que a taurina pode estar envolvida no metabolismo oxidativo.
Os danos em lipídeos das membranas celulares (lipoperoxidação) induzidos por ERO podem ocasionar prejuízos irreversíveis para a célula como a apoptose (FINAUD et al., 2006). A lipoperoxidação pode agravar o dano da membrana por aumento da permeabilidade (PASANTES-MORALES e CRUZ, 1985). Neste estudo, a suplementação de taurina diminuiu os níveis de TBARS (marcador indireto de lipoperoxidação) após o exercício excêntrico em comparação com os demais grupos. Alguns autores mostraram resultados similiares quanto ao efeito protetor da taurina sobre a lipoperoxidação (KIM et al., 1996; ZEYBECK et al., 2006). A taurina demonstrou reduzir o dano induzido pelo exercício no músculo, bloqueando o aumento de TBARS no músculo extensor longo de ratos no estudo de Dawson et al. (2002). Bakker e Berg (2002) reportam que ela altera indiretamente o funcionamento de canais de Cl- no músculo esquelético pela interação com os fosfolipídeos próximos ao canal. O efeito protetor que a taurina exerce sobre este tipo de lipídeo pode ser atribuído a essa capacidade de interação com os fosfolipídeos da membrana e a melhora nas propriedades de estabilização das paredes celulares, limitando o ataque de ERO (GOODMAN et al., 2009; ZHANG et al., 2004).