1 Recebido para publicação em 25.9.2012 e aprovado em 26.11.2012.
2 Gorgan University of Agricultural Sciences and Natural Resources (GUASNR), Gorgan, Iran, <[email protected]>.
FACTORS AFFECTING
Cyperus difformis
SEED GERMINATION AND
SEEDLING EMERGENCE
1Fatores que Afetam a Germinação de Sementes e a Emergência de Plântulas de
Cyperus
difformis
DERAKHSHAN, A.2 and GHEREKHLOO, J.2
ABSTRACT - Specific knowledge about the dormancy, germination, and emergence patterns of weed species aids the development of integrated management strategies. Laboratory studies were conducted to determine the effect of several environmental factors on seed germination and seedling emergence of Cyperus difformis. Germination of freshly harvested seeds was inhibited by darkness; however, when seeds were subsequently transferred to complete light they germinated readily. Our results showed that 2 wk of cold stratification overcome the light requirement for germination. Seeds of C.difformis were able to germinate over a broad range of temperatures (25/15, 30/20, 35/25, and 40/30 oC day/night). The response of
germination rate to temperature was described as a non-linear function. Based on model outputs, the base, the optimum and the ceiling temperatures were estimated as 14.81, 37.72 and 45 oC, respectively. A temperature of 120 oC for a 5 min was required to inhibit 50% of
maximum germination. The osmotic potential and salinity required for 50% inhibition of maximum germination were -0.47 MPa and 135.57 mM, respectively. High percentage of seed germination (89%) was observed at pH=6 and decreased to 12% at alkaline medium (pH 9) pH. Seeds sown on the soil surface gave the greatest percentage of seedling emergence, and no seedlings emerged from seeds buried in soil at depths of 1 cm.
Keywords: osmotic potential, salinity stress, pH, soil depth, temperature.
RESUMO - O conhecimento específico sobre os padrões de dormência, germinação e emergência de espécies de plantas daninhas auxilia o desenvolvimento de estratégias integradas de gestão. Foram realizados estudos de laboratório para determinar o efeito de vários fatores ambientais na germinação
de sementes e emergência de plântulas de Cyperus difformis. A germinação das sementes
recém-colhidas foi inibida pela escuridão; no entanto, quando as sementes foram transferidas posteriormente para a luz total, germinaram prontamente. Nossos resultados mostraram que duas semanas de
estratificação fria superam o requisito de luz para a germinação. As sementes de C. difformis
conseguiram germinar numa ampla gama de temperaturas (25/15, 30/20, 35/25, e 40/30 oC dia/
noite). A resposta da taxa de germinação à temperatura foi descrito como uma função não-linear. Com base em resultados do modelo, as temperaturas de base, ótima e e teto foram estimadas em 14,81,
37,72 e 45 oC, respectivamente. Foi preciso uma temperatura de 120 oC por mais 5 minutos para
inibir 50% da germinação máxima. O potencial osmótico e a salinidade necessários para uma inibição
de 50% da máxima germinação foram -0,47MPa e 135,57mM, respectivamente. Foi observada uma
alta percentagem de germinação das sementes (89%) com pH=6, a qual diminuiu para 12% em meio alcalino (pH 9) de pH. As sementes semeadas na superfície do solo resultaram na maior porcentagem de emergência de plântulas, e nenhuma plântula emergiu a partir de sementes enterradas no solo a
uma profundidade de 1cm.
INTRODUCTION
Cyperus difformis is a species of sedge known by several common names, including variable flatsedge and smallflower umbrella-sedge. This plant is native to southern Europe, most of Africa, Asia, and Australia, and it is naturalized in other areas of the world (Holm et al., 1991). It is a prolific seed producer, which normally grows in flooded or very moist soils. This plant is primarily a weed of paddy or flooded rice, and it is a weed of rice in 46 countries (Rao et al., 2007). It is frequently found in small pools, along rivers, canals, and streams, in open wet places, and in grassy swamps. It grows best in rich, fertile soils, but it can also grow in poorer sandy or clay soils of unused lands or in fallow rice fields (Holm et al., 1991; Chauhan & Johnson, 2009a). The ability of the species to complete a vegetative and reproductive cycle within a month or so makes it especially competitive in a crop that requires at least 90 or more days to reach maturity (Holm et al., 1991).
Seed germination is a key event in determining the success of a weed in an agro-ecosystem and it may be regulated by several factors, including temperature, light, soil salinity, moisture, and pH. Dormancy status influences the germination of seeds in light and dark. Non-dormant seeds of some species germinate more readily under light than under dark conditions (Baskin & Baskin, 1998). Temperature plays a major role in determining the periodicity of seed germination and the distribution of species (Guan et al., 2009). Germination rate usually increases linearly with temperature, at least within a well-defined range, and declines sharply at higher temperatures (Alvarado & Bradford, 2002). Knowledge of the effect of constant and alternating temperatures on C. difformisgermination can help us understand its emergence pattern and its management in different cropping systems. Light is an important ecological determinate for germination; the absence of light acts as a soil-depth indicator that prevents many seeds from germinating (Crisraudo et al., 2007). Similarly, an ability to germinate under conditions of moisture stress or high salinity can enable a weed to take advantage of conditions that limit the growth of other
species. In preparation for cropping, farmers in many areas burn the cut fallow vegetation or crop residues before the crop is ‘‘dibble sown.’’ The heat generated by burning may also affect weed seed germination.
Little information exists about the effect
of environmental factors on C. difformis
germination in Iran. Some information on the
germination ecology of C. difformis is available
in the Philippines (Chauhan & Johnson, 2009a). In previous studies, germination of C. difformis was stimulated by light and warm fluctuating temperatures. Germination decreased by 50% of the maximum rate at an osmotic potential of -0.12 MPa and only 23 mM NaCl was required to inhibit 50% of maximum germination (Chauhan & Johnson, 2009a).
Although the information on C. difformis can
be useful, observations suggest that the
germination requirements of C. difformis in
Iran are different from those in the Philippines. A better understanding of germination of
C. difformis seeds would be useful in developing effective control measures. The objectives of this study were to determine the effect of light, temperature, pH, osmotic
stress, and salt stress on C. difformis seed
germination and emergence.
MATERIALS AND METHODS
Experiments were conducted in 2011 at the Seed Technology Laboratory, Department of Agronomy, Gorgan University of Agricultural Science and Natural Resources, Iran (latitude
36o51 ‘N, longitude 54o16 ‘E). Seeds of
C. difformis were collected from several rice fields in Golestan, a province of Iran in September 2011. Seeds collected from many randomly selected plants were bulked, cleaned manually, placed in a plastic container, and
stored at 5 oC.
General germination test protocols
Germination of Cyperus difformis was
water used for replenishing moisture was kept in an incubator to ensure a water temperature consistent with the incubator. Germination was determined after 14d, at which time seeds with emerged radicles (>2mm) were considered to have germinated.
Effect of fluctuating temperature and light on germination
Germination of freshly harvested seeds was determined in growth chambers under fluctuating day/night temperatures (40/30,
35/25, 30/20, and 25/15 oC) in both light/
dark and continuous dark conditions. The photoperiod was set at 12 h to coincide with the high-temperature phase. For germination in complete darkness, dishes were wrapped in two layers of aluminum foil.
Effect of cold stratification on germination
Seeds were placed between two pieces of Whatman No. 1 filter paper moistened with distilled water in a 9-cm-diam Petri dish then wrapped in two layers of aluminum foil. Samples were subsequently stored in a
refrigerator (5 oC) for 2 wk. Germination was
determined after the cold stratification period under fluctuating day/night temperature 35/
25 oC in both light/dark and continuous dark
conditions.
Effect of constant temperature
The seeds of C. difformis were incubated
under fluctuating day/night conditions (12/ 12 h light/dark) at 7 constant temperatures
(from 15 oC to 45 oC with 5 oC intervals).
Germinated seeds were counted at different time intervals based on observed germination rate under different temperatures, with shorter and longer periods for higher and lower temperatures, respectively. Time estimates taken for cumulative germination to reach 50% of its maximum at each replicate were interpolated from the germination progress curve versus time. The times for 10% and 90% germination were also determined by interpolation and are designated as D10 and D90.
Effect of high temperature ‘‘pretreatment’’ on germination
A study was conducted to determine the effect of high temperatures, as seeds
might experience during the burning of fallow vegetation, on seed germination of
C. difformis. Fifty seeds were placed in an oven in an open container at 60, 80, 100, 120,
140, 160, 180, and 200 oC temperatures for
5 min. The treated seeds were subsequently
incubated at 35/25 oC day/night temperature
in a 12-h photoperiod for 14 d. In the control treatment, seeds were kept at room
temperature (25 oC) before being incubated.
Effect of salt and osmotic stress on germination
To examine the effect of salinity on
C. difformis germination, seeds were placed in dishes containing 5 ml of solution of 0, 25, 50, 100, 150 and 200 mM NaCl. In a separate study, the effects of moisture stress on seed germination were assessed by incubating seeds in solutions with osmotic potentials of 0, -0.1, -0.2, -0.4, -0.6, -0.8, and -1 MPa, prepared by dissolving polyethylene glycol 8000 in distilled water, as described by Michel
(1983).The seeds were incubated at 35/25 oC
day/night temperature in a 12-h photoperiod for 14 d.
Effect of pH of buffered solutions on germination
Seeds were placed on filter papers soaked with 5 mL of buffered solutions with pH ranging from 4 to 9. Buffer solutions were prepared using potassium hydrogen phthalate in combination with either 0.1 M HCl or 0.1 M NaOH to obtain solution pH levels of 3, 4, 5, and 6. A 25 mM sodium tetraborate decahydrate solution was used in combination with control either of 0.1 M HCl or 0.1 M NaOH to obtain solution pH levels of 7, 8, and 9 (Shaw et al.,
1991). The seeds were incubated at 35/25 oC
day/night temperature in a 12-h photoperiod for 14 d.
Effect of seed burial depth on seedling emergence
through a 3 mm sieve before commencing the experiment. Soil in the pots was moistened initially using a mist sprayer and then sub-irrigated to maintain adequate soil moisture. Seedling emergence was defined as the coleoptile being visible at the soil surface. The experiment was terminated when no further emergence was recorded for a 3 wk period.
Statistical analyses
A randomized complete block design with four replications was used in all experiments. Each replicate was arranged on a different shelf or bench in the germination chamber and considered as a block. Protected LSD test at the 5% probability level was used to evaluate differences between the effect of temperature and light. Regression analysis was used to determine the effect of high temperature pretreatment, salt and osmotic stress, pH, burial depth, and to quantify the cardinal temperatures.
The segmented model (Equation 1) was used to quantify the response of germination rate to temperature and to estimate cardinal temperatures (Ritchie & NeSmith, 1991).
b b
T
T
T
T
T
f
0)
(
ifb
b
T
T
T
(eq. 1)
0 01
)
(
T
T
T
T
T
f
cif
T
0
T
T
c0
)
(
T
f
ifT
T
b orT
T
cwhere, T is the temperature, and Tb, To
and Tc are the base, optimum and ceiling
temperatures, respectively. High temperature pretreatment, salt and osmotic stress data showed a sigmoid trend and a three-parameter logistic model was fitted to data (Equation 2).
rate maxG
x
x
G
G
50exp
1
(eq. 2)where G is the percentage germination at
temperature and concentration x of NaCl or
osmotic potential, Gmax is the maximum
germination (%), x50 is the temperature, NaCl
concentration or osmotic potential required for 50% inhibition of maximum germination and
Grate indicates the slope of the curve in x50. pH
data were subjected to a binomial model. An exponential function was fitted to the seedling emergence (%) data (Equation 3).
E = Emaxexp(– Eratex) (eq. 3)
where E represents emergence (%), Emax is the
maximum emergence and Erate indicates the
slope. Statistical Analysis System (SAS, 1989) and SigmaPlot 8.0 were used for analyzing data.
RESULTS AND DISCUSSION
Effect of fluctuating temperature and light on germination
Germination was inhibited by darkness; however, when seeds were subsequently transferred to complete light, they germinated readily. Seed germination response to light varies considerably from species to species. Seeds of some species require light to germinate (Chauhan & Johnson, 2009a) while others germinate equally in light and dark (Teuton et al., 2004). In the study of Chauhan & Johnson (2009a), three sedge species
(including C.difformis, C.iria and Fimbristylis
miliacea) had an absolute light requirement for germination. The results of the present study
confirmed that seeds of C.difformis are
positively photoblastic. The response to light is thought to be controlled by phytochrome, a light-absorbing pigment within most plants. In photoblastic seeds, light exposure may convert inactive-phytochrome ‘‘red’’ to the active-phytochrome ‘‘far-red.’’ Germination of
C.difformis seeds in light (optimum condition) suggests that its germination and subsequent emergence in the field will be favored by the presence of seeds at, or near, the soil surface.
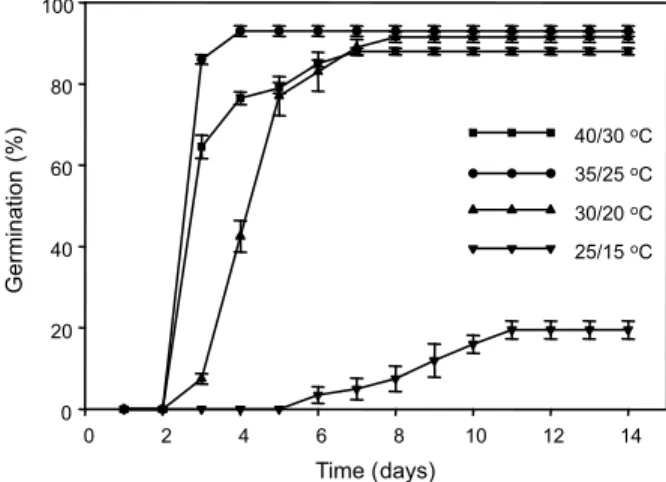
Seed germination of C.difformis was
influenced (P≤0.01) by fluctuating day/night
conditions (Figure 1). C. difformis seeds
had greater germination at 30/20 oC (91.5%),
35/25 oC (93%) and 40/30 oC (88%) alternating
day/night temperatures than at 25/15 oC.
Germination rate differed among the tested temperatures, with germination being greater
35/25 oC and 40/30 oC) than at the coldest
temperature (25/15 oC). The time to onset
of germination and rate of increase were influenced by alternating temperature regimes. Time to onset of germination decreased with increasing temperature. For example, the 35/25 alternating temperature regime conditions caused the fastest onset of germination and highest rate of germination compared to the other three alternating
temperature regimes; therefore, 35/25 oC was
selected for the subsequent experiments. These data suggest that germination might be limited to a small proportion of seeds in cooler regions of the tropics or during cool seasons. This temperature enhancement of seed germination has been reported in other weed
species, including, synedrella (Synedrella
nodiflora) (Chauhan & Johnson, 2009b), rice
flatsedge (C. iria) and lesser fimbristylis
(F. miliacea) (Chauhan & Johnson, 2009a). High germination percentages immediately after seed harvest indicating a low level of primary dormancy in this species.
Effect of cold stratification on germination
While cold stratification is an absolute requirement for germination for some species; in others, it may only improve the percentage and rate of germination and decrease the time to the onset of germination. After 2 wk of cold stratification, the germination percentage of
C. difformis seeds was similar (more than 90%) in both light/dark and continuous dark conditions under fluctuating day/night
temperature 35/25 oC. The light requirement
for C. difformis germination was completely overcome by cold stratification. The ability of cold stratification to overcome the light requirement for germination was previously
reported for common groundsel (Senecio
vulgaris) (Popay & Roberts, 1970), short’s
goldenrod (Solidago shortii) (Walck et al., 1997)
and black nightshade (Solanum ptycanthum)
(Zhou et al., 2005).
Effect of constant temperature on germination
In the fluctuating day/night condition (12/ 12 h light/dark), the highest germination percentiles have been obtained between 25 to
40 oC and germination reached zero at 15 and
45 oC (Figure 2). Seeds have the capacity to
germinate over a defined range, characteristic for each species; hence, there are clear minimum and maximum temperatures for germination and, between them, a broad range over which germination of all seeds can be attained (Bewley & Black, 1994).
The influence of temperature on the germination rate was described by a segmented model. The model fitted to relative germination rate vs. mean temperature for 3 percentiles (D10, D50 and D90) is shown in
Figure 1 - Effect of fluctuating day/night temperatures (25/15, 30/20, 35/25 and 40/30 oC) on seed germination of
C. difformis (12/12 h light/dark).
Time (days)
0 2 4 6 8 10 12 14
Germination
(%)
0 20 40 60 80 100
40/30oC
35/25oC 30/20oC
25/15oC
Figure 2 - Effect of constant temperatures (20, 25, 30, 35 and 40 oC) on seed germination of C. difformis in the fluctuating day/night conditions (12/12 h light/dark).
Time (h)
0 100 200 300
Germination
(%)
0 20 40 60 80 100
20oC
25oC
30oC
35oC
Figure 3. According to the segmented model for percentiles of D10, D50 and D90, the base
temperature ranged from 14.46 oC to 15.39 oC.
The optimum temperature for all percentiles
varied between 36.97 oC and 38.23 oC.
Model outputs, the base and the optimum
temperatures were estimated as 14.81 oC and
37.72 oC, respectively. Ceiling temperature for
all percentiles was estimated as 45 oC. In a
previous study (Chozin & Nakagawa, 1988),
greater germination of C. iria was obtained at
a constant temperature of 35 oC (69%) than at
15 oC (17%). Similar types of results were
observed for different plants (Vidal et al., 2007; Rizzardi et al., 2009; Ghaderi et al., 2010).
Germination rate is more temperature sensitive than final germination percentage in Setaria lutescens and Amaranthus retroflexus
(Bewley & Black, 1994) similar to our finding for C.difformis. Germination rate is most rapid at optimum temperature and declined with decrease and/or increase temperature. A decline in germination rate with decreasing temperature is partly associated with decline in the imbibition rate observed with a reduction in temperature (Bewley & Black, 1994). Germination rate is affected by the depth of dormancy, imbibition rate and the rate of catabolic and anabolic pathways all of which are directly or indirectly temperature dependent while the maximum seed germination is more affected by the rate of
rehydration rather than the speed of the physiological pathways affecting cell expansion (Bewley & Black, 1994).
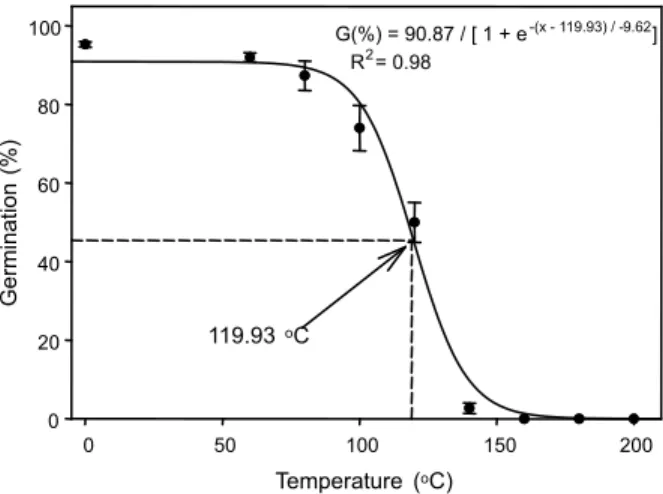
Effect of high temperature ‘‘pretreatment’’ on germination
A sigmoid response was observed in the
germination of C. difformis to the pretreatment
of high temperatures (Figure 4). Germination was greater than 74% up to a temperature of
100 oC but decreased progressively with further
increases in temperature, resulting in no
germination after ≥ 160 oC. Estimates from the
fitted model indicate that a temperature of
120 oC was required for a 5 min interval to
inhibit 50% of maximum germination. Many farmers in tropical countries practice vegetation burning to clear their land of debris before crop planting (Roder et al., 1997). Fire-induced high temperatures can destroy the seeds and can also affect some allelochemicals known to inhibit seed germination (Roder et al., 1997). Burning vegetation can increase
surface temperature to 550 oC (Cook, 1939);
however, temperature can decrease at a rate
of 100 oC cm-1 in the first 5 cm below the soil
surface (Sanchez, 1976). Assuming these values, although seeds on the soil surface might be destroyed by high temperature, seeds
Figure 3 - Predicted (lines) vs. observed (symbols) germination rate ofC. difformis at different constant temperatures for different germination percentiles (D10, D50 and D90) using the segmented model.
Figure 4 - Effect of 5 min exposures to a range of oven temperatures on seed germination of C. difformis after 14 d of incubation in light/dark at 35/25 oC fluctuating day/night temperature. The line represents a three-parameter sigmoid model fitted to the data. The temperature required for 50% inhibition of maximum germination was estimated as 119.93 oC.
Temperature (oC)
10 20 30 40 50
Germination
rate (1/h)
0.000 0.005 0.010 0.015 0.020 0.025 0.030
D10 D50 D90 D10 D50 D90
Temperature (oC)
0 50 100 150 200
Germination
(%)
0 20 40 60 80
100 G(%) = 90.87 / [ 1 + e-(x - 119.93) / -9.62]
R2= 0.98
buried below 4 cm could escape from the effect of heat. These seeds can remain dormant while buried; however, they can germinate when brought onto the soil surface by inter-cultivation or tillage operations and exposed to light.
Effect of Salt and osmotic stress on germination
A three-parameter sigmoid model was fitted to the germination percentage obtained at different concentrations of NaCl (Figure 5). Germination was greater than 85% up to the concentration of 100 mM NaCl; 29% germination occurred at 150 mM NaCl, although germination was completely inhibited at 200 mM NaCl. The concentration required for 50% inhibition of maximum germination was 135.57 mM NaCl. Salinity is a major abiotic constraint for plants and can negatively affect important physiological processes. In addition to physiological processes in the plants, sodium can alter soil structure and fertility by replacing calcium and magnesium in the anion exchange process, and that leads to nutrient and water stress (DiTommaso, 2004).
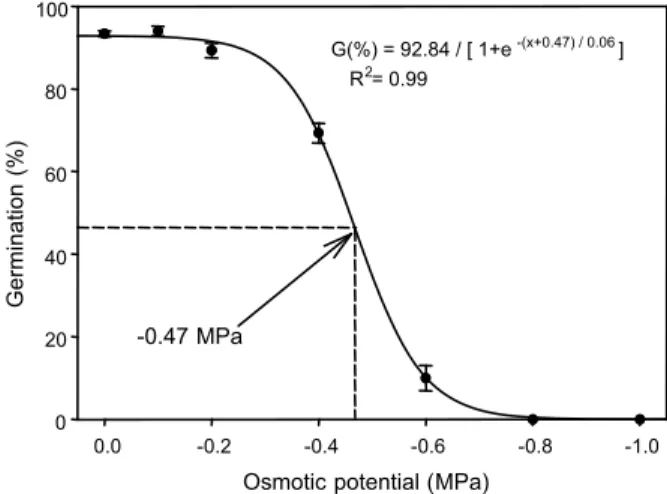
A three-parameter sigmoid model was fitted to the germination values obtained at different osmotic potentials (Figure 6).
Germination decreased from 94 to 10% as osmotic potential decreased from 0 to -0.6 MPa, and it was completely inhibited at osmotic potential of -0.8 MPa. The osmotic potential required for 50% inhibition of the maximum germination was -0.47 MPa. As water potential decreased beyond -0.4 MPa, germination decreased sharply, suggesting
that C. difformis favors a moist environment
for germination. This is supported by the occurrence of this weed in lowland fields where soil moisture is high, whereas in upland fields germination would most likely occur in the rainy season.
Effect of pH of buffered solutions on germination
A quadratic model was fitted to the germination values obtained in response to pH of
buffered solution (Figure 7). Seed germination
has occurred in a range of pH between 4 and 9, but the maximum germination (89%) was observed at pH=6. Germination decreased to 80 and 12% at pH 4 and 9, respectively. Seed of many species germinate to high percentages over a wide range of pH values (Jain & Singh, 1989; Souza Filho et al., 2001), but those of others germinate to high percentages only at specific pH values (Wilson Jr., 1979; Chachalis & Reddy, 2000).These results suggest that seeds
Figure 5 - Effect of NaCl concentration on seed germination of C. difformis after 14 d of incubation in light/dark at 35/25 oC fluctuating day/night temperature. The line represents a three-parameter sigmoid model fitted to the data. The salt concentration required for 50% inhibition of maximum germination was estimated as 135.57 mM.
Figure 6 - Effect of osmotic potential on seed germination of C. difformis after 14 d of incubation in light/dark at 35/25 oC fluctuating day/night temperature. The line represents a three-parameter sigmoid model fitted to the data. The osmotic potential required for 50% inhibition of the maximum germination was estimated as -0.47 MPa.
NaCl concentration (mM)
0 50 100 150 200 250
Germination
(%)
0 20 40 60 80 100
G(%) = 96.38 / [ 1+e-(x-135.57) / -17.01] R2= 0.99
135.57mM
Osmotic potential (MPa)
-1.0 -0.8
-0.6 -0.4
-0.2 0.0
Germination
(%)
0 20 40 60 80 100
G(%) = 92.84 / [ 1+e-(x+0.47) / 0.06] R2= 0.99
of C.difformis cannot survive in very alkaline conditions and that a pH below 7 was favorable
for germination of C. difformis.
Effect of seed burial depth on seedling emergence
Seed burial depth greatly affected seedling
emergence of C. difformis (Figure 8). The
greatest level of emergence (67%) was observed for seeds placed on the soil surface. Emergence decreased sharply with increasing burial depth, and no seedlings emerged at burial depths of 1 cm or greater. These results corroborate those of the studies on temperature and light. It has been reported that very little light is transmitted by any type of soil at a depth greater than 4 mm (Benvenuti, 1995). Because of the absolute requirement for light, limited light penetration may be a major reason for no emergence from buried seeds. In addition, the magnitude of temperature fluctuation in soils increases near the soil surface and declines in the soil profile (Rubin & Benjamin, 1984). Hypoxia and low rates of gaseous diffusion at deeper depths could be other reasons for the lack of emergence (Benvenuti, 2003). Besides,
small-seeded species such as C.difformis may not
have enough reserves to support seedling emergence from depth.
In conclusion, the highest germination
levels in C. difformis at 30/20 oC, 35/25 oC
and 40/30 oC alternating temperatures
indicating that higher range of the tested temperatures favour germination of this species. Germination of freshly harvested seeds was inhibited by darkness. Our results show that 2 wk of cold stratification overcome the light requirement for germination. Segmented model outputs, the base, the optimum and the ceiling temperatures
were estimated as 14.81, 37.72 and 45 oC,
respectively. Greater emergence from seeds placed on the soil surface suggests that no-till farming practices would favour emergence of
C. difformis in the field. As this species could not emerge from a depth of 1 cm, shallow tillage operations that bury seeds below this depth could be an option to limit the
germination of C. difformis. This may have
practical significance in weed management systems as tillage options may be devised to limit the portion of the soil seed bank exposed to the surface layers and thus the ability to germinate. It should be noted that germination responses to some factors (e.g., osmotic potential and salinity) were different between our study and the study conducted with seeds of the Philippines population (Chauhan & Johnson, 2009a). This suggests that
C. difformis could be polymorphic in its response to some factors, and further studies are required to elucidate this. If differences between populations exist, this could warrant different management practices in different countries.
Figure 7 - Effect of pH of buffered solution on seed germination of C. difformis after 14 d of incubation in light/dark at 35/ 25 oC fluctuating day/night temperature. The line represents a quadratic model fitted to the data.
Figure 8 - Effect of seed burial at various soil depths on seedling emergence of C. difformis after 21 d of incubation in light/ dark at 35/25 oC fluctuating day/night temperature. The line represents an exponential curve fitted to the data.
pH
4 5 6 7 8 9
Germination
(%)
0 20 40 60 80 100
Y= -5.58 x2+ 58.21 x - 63.11
R2= 0.96
Burial depth (cm)
0 1 2 3 4 5
Seedling
emergence
(%)
0 20 40 60 80 100
LITERATURE CITED
ALVARADO, V.; BRADFORD, K. J. A hydrothermal time model explains the cardinal temperatures for seed germination. Plant Cell. Environ., v. 25, n. 8, p. 1061-1069, 2002.
BASKIN, C. C.; BASKIN, J. M. Seeds: ecology, biogeography and evolution of dormancy and germination. New York: Academic Press, 1998. p. 666.
BENVENUTI, S. Soil light penetration and dormancy of jimsonweed (Datura stramonium) seeds. Weed Sci., v. 43, n. 3, p. 389-393, 1995.
BENVENUTI, S. Soil texture involvement in germination and emergence of buried weed seeds. Agron. J., v. 95, n. 1, p. 191-198, 2003.
BEWLEY, J. D.; BLACK, M. Seeds: physiology of development and germination. 2.ed. New York: Plenum Press, 1994. p. 445.
CHACHALIS, D.; REDDY, K. N. Factors affecting Campsis radicans seed germination and seedling emergence. Weed Sci., v. 48, n. 2, p. 212-216, 2000.
CHAUHAN, B. S.; JOHNSON, D. E. Ecological studies on Cyperus difformis, Cyperus iria and Fimbristylis miliacea: three troublesome annual sedge weeds of rice. Ann. Appl. Biol., v. 155, n. 1, p. 103-112, 2009a.
CHAUHAN, B. S.; JOHNSON, D. E. Seed germination and seedling emergence of synedrella (synedrella nodiflora) in a tropical environment. Weed Sci., v. 57, n. 1, p. 36-42, 2009b.
CHOZIN, M. A.; NAKAGAWA, K. Autecological studies on Cyperus iria L. and C.microiria Steud., annual Cyperaceous weeds. Weed Res., v. 33, n. 1, p. 23-30, 1988.
COOK, L. A contribution to our information on grass burning. Southern. Afr. J. Sci., v. 36, n. 1, p. 270-282, 1939.
CRISRAUDO, A. et al. Effects of after harvest period and environmental factors on seed dormancy of Amaranthus species. Weed Res., v. 47, n. 4, p. 327-334, 2007.
DITOMMASO, A. Germination behavior of common ragweed (Ambrosia artemisiifolia) populations across a range of salinities. Weed Sci., v. 52, n. 6, p. 1002-1009, 2004.
GHADERI, F. A. et al. Influence of environmental factors on seed germination and seedling emergence of yellow sweet clover (melilotus officinalis). Planta Daninha, v. 28, n. 3, p. 463-469, 2010.
GUAN, B. et al. Germination responses of Medicago ruthenica seeds to salinity, alkalinity and temperature. J. Arid Environ., v. 73, n. 1, p. 135-138, 2009.
HOLM, L. G. et al. The world’s worst weeds: distribution and biology. Honolulu: University of Hawaii Press, 1991. p. 609.
JAIN, R.; SINGH, M. Factors affecting goatweed (Scoparia dulcis) seed germination. Weed Sci., v. 37, n. 6, p. 766-770, 1989.
MICHEL, B. E. Evaluation of water potential of solution of polyethylene glycol 8000 both in absence and presence of other solutes. Plant Physiol., v. 72, n. 1, p. 66-70, 1983.
POPAY, A. I.; ROBERTS, E. H. Factors involved in the dormancy and germination of Capsella bursa-pastoris (L.) Medik. and Senecio vulgaris (L.). J. Ecol., v. 58, n. 1, p. 103-121, 1970.
RAO, A. N. et al. Weed management in direct-seeded rice. Adv. Agron., v. 93, n. 1, p. 153-255, 2007.
RITCHIE, J. T.; NESMITH, D. S. Temperature and crop development. In: HANKS, J.; RITCHIE, J. T., (Ed.). Modeling plant and soil systems. Madison: ASA, CSSSA, SSSA, 1991. p. 452.
RIZZARDI, M. A. et al. Effect of cardinal temperature and water potential on morning glory (Ipomoea triloba) seed germination. Planta Daninha, v. 27, n. 1, p. 13-21, 2009.
RODER, W. et al. Weeds in slash-and-burn rice fields in northern Laos. Weed Res., v. 37, n. 2, p. 111-119, 1997.
RUBIN, B.; BENJAMIN, A. Solar heating of the soil: involvement of environmental factors in the weed control process. Weed Sci., v. 32, n. 1, p. 138-142, 1984.
SANCHEZ, P. A. Soil management in shifting cultivation areas. In: SANCHEZ, P. A. (Ed.). Properties and
management of soils in the tropics. New York: John Wiley & Sons, 1976. p.347-412.
SAS Institute. User’s guide. Version 6. 4.ed. Cary: 1989. p. 943.
SHAW, D. R. et al. Redvine (Brunnichia ovata) germination and emergence. Weed Sci., v. 39, n. 1, p. 33-36, 1991.
SOUZA FILHO, A. P. S. et al. Seed germination in weeds from cultivated pasture areas: Mimosa pudica and Ipomoea asarifolia. Planta Daninha, v. 19, n. 1, p. 23-31, 2001.
TEUTON T. C. et al. Factors affecting seed germination of tropical signalgrass (Urochloa subquadripara). Weed Sci., v. 52, n. 3, p. 376-381, 2004.
VIDAL, R. A. et al. Impact of temperature, light and seed depth on emergence and germination of Conyza bonariensis and Conyza Canadensis resistant to glyphosate.
Planta Daninha, v. 25, n. 2, p. 309-315, 2007.
WALCK, J. L. et al. A comparative study of the seed germination biology of a narrow endemic and two
WILSON JR., R. G. Germination and seedling development of Canada thistle (Cirsium arvense). Weed Sci., v. 27, n. 2, p. 146-151, 1979.