Identification, classification and expression pattern analysis of sugarcane cysteine proteinases
Texto
Imagem


Documentos relacionados
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Concomitantemente ao desenvolvimento do método analítico, as concentrações dos elementos menores e maiores em amostras dos bivalves coletados ao longo da Baía de Todos
Entretanto, o modelo de Reiners e Mohanty para as estrelas binárias, assim como para estrelas simples, não é capaz de descrever o comportamento evolutivo de estrelas do tipo espectral
A nosso parecer, se a Fazenda Pública demonstrar que a jurisprudência do plenário do Supremo Tribunal Federal não reflete o entendimento prevalente, mas um
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
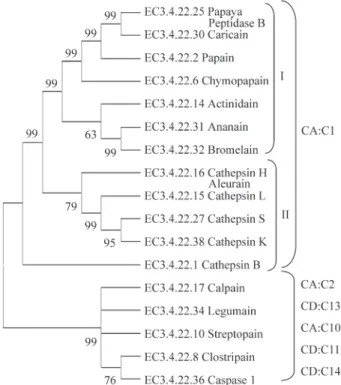
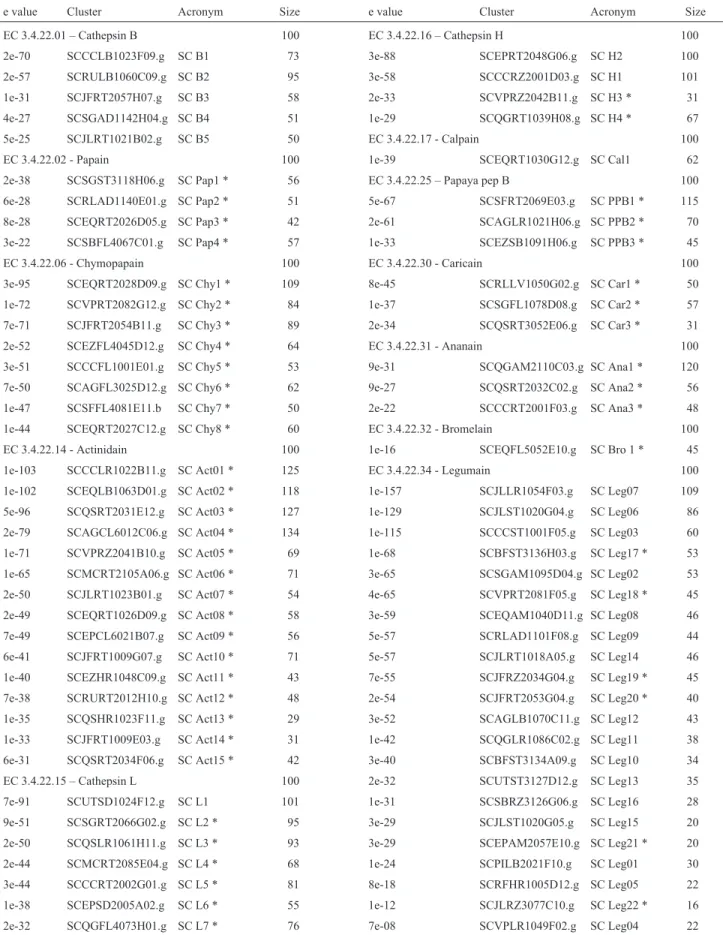
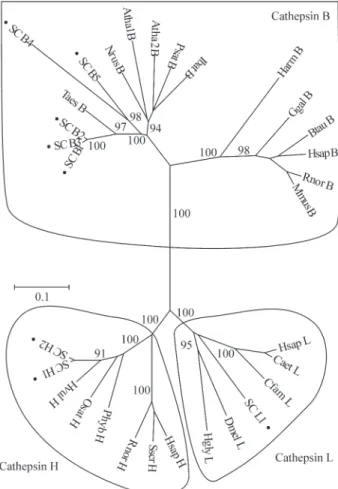
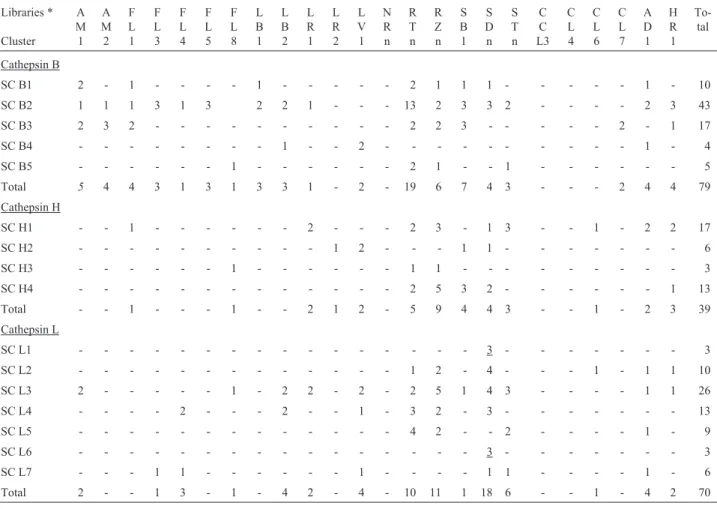
A set of genes related to secondary metabolism was extracted from the sugarcane expressed sequence tag (SUCEST) database and was used to investigate both the gene expression pattern
Subclass III GRPs have never before been isolated from monocotyledonous plants, but we identified ten clus- ters in the SUCEST data bank, six of them representing pu-
A classificação de MSRM é atribuída, segundo o Decreto-Lei n 128/2013, de 5 de setembro, quando preencham uma das seguintes condições: possam necessitar de vigilância
