Braz. j. oceanogr. vol.64 número2
Texto
Imagem
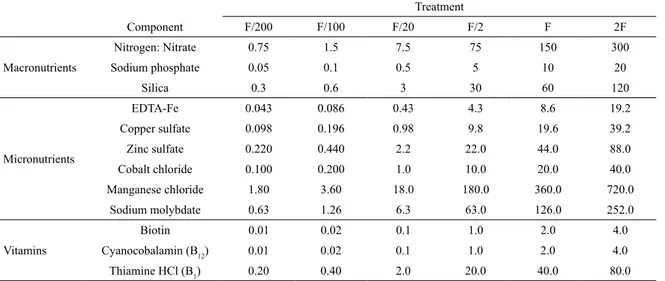
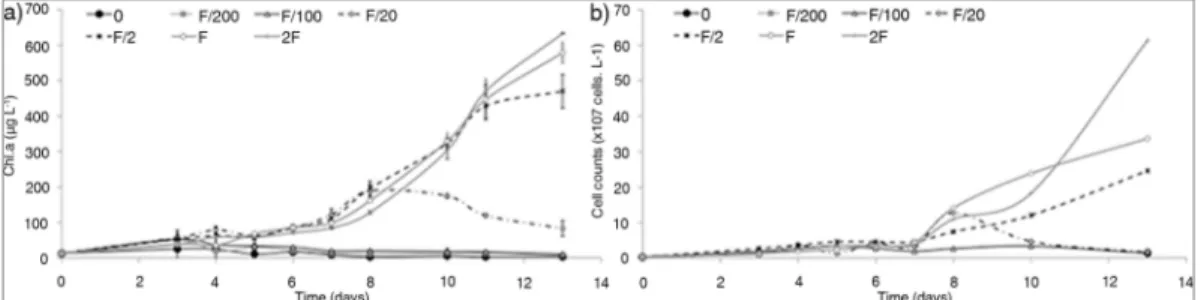
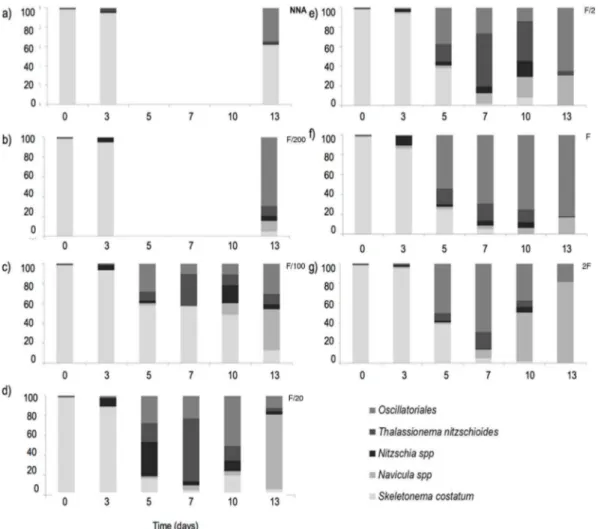
Documentos relacionados
Extinction with social support is blocked by the protein synthesis inhibitors anisomycin and rapamycin and by the inhibitor of gene expression 5,6-dichloro-1- β-
i) A condutividade da matriz vítrea diminui com o aumento do tempo de tratamento térmico (Fig.. 241 pequena quantidade de cristais existentes na amostra já provoca um efeito
It is for these reasons that the present study seeks to report on the occurrence of Litopenaeus vannamei in the estuarine lagoon complex of Cananeia, Iguape
The objective of this study was to determine the Universal Soil Loss Equation (USLE) rainfall erosivity factor (R) by using the standard procedure based on pluviographic records
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
Nos últimos anos, a Segurança da Informação deixou de ser um tema da TI e alcançou o alto escalão das empresas, passando a exigir todo um trabalho jurídico para harmonizar
