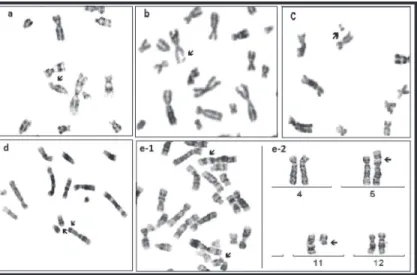
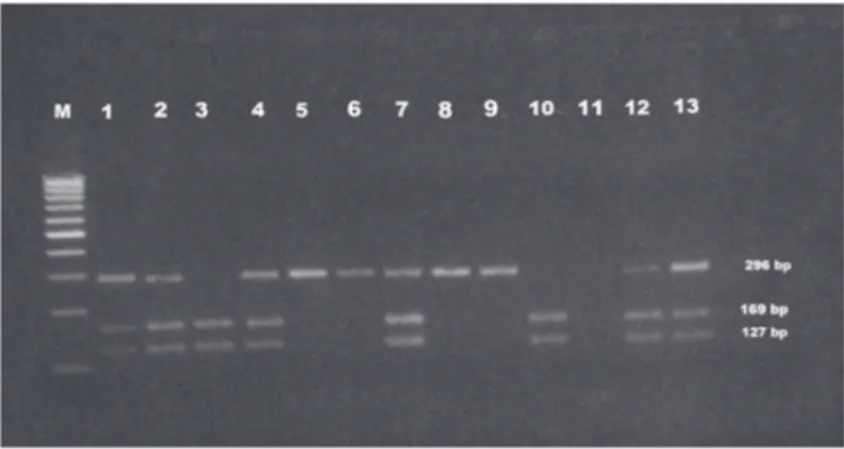
Role of p53 codon 72 polymorphism in chromosomal aberrations and mitotic index in patients with chronic hepatitis B
Texto
Imagem
Documentos relacionados
Declaro estar ciente e ter compreendido a proposta de pesquisa abaixo descrita, consentindo desta forma em participar como voluntário do projeto “Avaliação do torque nos movimentos
The prevalence of cholelithiasis was higher in patients with SS homozygous and S β heterozygous thalassemia when compared to patients with sickle cell disease.. In 50% of
Objective - To compare Doppler parameters of portal vein in patients with chronic hepatitis C with a healthy control group and to correlate these parameters with ibrosis
A matriz curricular do curso FIC em “Práticas Pedagógicas e Saberes Indígenas na Escola”, integrado à Ação “Saberes Indígenas na Escola” (SECADI/MEC),
In the present study, the activation status of B cells from peripheral blood of healthy controls (N = 20) and patients with acute hepatitis B (AHB, N = 15) or chronic hepatitis
In our study, we only evaluated patients hospitalized in the ICU, but even when compared with similar populations of patients reported in literature, we observed a
