UNIVERSIDADE DO ALGARVE
Benthic foraminifera as palaeo-environmental
indicators in the Northern Gulf of Cadiz
Isabel Maria de Paiva Pinto Mendes
Doutoramento em Ciências do Mar,
especialidade em Oceanografia
UNIVERSIDADE DO ALGARVE
Benthic foraminifera as palaeo-environmental
indicators in the Northern Gulf of Cadiz
Isabel Maria de Paiva Pinto Mendes
Doutoramento em Ciências do Mar,
especialidade em Oceanografia
Tese orientada por:
Prof. Doutor João Manuel Alveirinho Dias
Doutor
Joachim
Schönfeld
i
Resumo
Esta tese apresenta uma investigação sobre os foraminíferos bentónicos como indicadores paleo-ambientais na plataforma continental do Norte do Golfo de Cádis, nordeste do Oceano Atlântico. O principal objectivo foi determinar o valor dos foraminíferos bentónicos vivos como indicadores ambientais e sua aplicação em reconstruções paleoambientais durante o Holocénico.
As relações entre as faunas de foraminíferos bentónicos vivos e os parâmetros físico-químicos associados foram analisados em amostras de sedimentos superficiais, colhidos na plataforma continental entre as desembocaduras dos Rios Guadiana e Guadalquivir. As faunas de foraminíferos bentónicos vivos foram caracterizadas em detalhe e as associações das espécies mais abundantes classificadas em quatro grupos. Estes grupos permitiram caracterizar do ambiente actual e os parâmetros físico-químicos que lhe estão associados.
O estudo sedimentológico e dos foraminíferos bentónicos, enquadrado numa janela cronológica, foi realizado em três sequências sedimentares Holocénicas, colhidas em ambientes sedimentares contrastantes. A comparação entre as faunas vivas e as associações de foraminíferos bentónicos mais abundantes que ocorrem na plataforma foi também realizada. Uma abordagem integradora, usando toda a informação disponível, permitiu clarificar as alterações peloambientais, revelando cinco cenários evolutivos. Cada cenário representa as alterações ambientais ocorridas em cada ambiente estudado, durante diferentes períodos do Holocénico, influenciado por diversos factores, tais como as alterações do nível médio do mar, as descargas do rio e as propriedades do sedimento, que são reflectidas pelas diferentes associações de foraminíferos bentónicos.
Esta tese demonstra que as preferências ecológicas das espécies actuais de foraminíferos bentónicos mais abundantes, tem aplicação directa na interpretação do registo paleoambiental.
Palavras-chave: Golfo de Cádis; plataforma continental; foraminíferos bentónicos; ecologia; Holocénico
Abstract
This thesis is an investigation of benthic foraminifera as palaeo-environmental indicators on the Northern Gulf of Cadiz continental shelf, northeast Atlantic Ocean. The main objective was to determine the environmental indicator value of living benthic foraminifera and their application to Holocene palaeoenvironmental reconstruction.
Relationships between living benthic foraminiferal fauna and both physical and chemical parameters were analysed in surficial samples collected from the continental shelf between the Guadiana and Guadalquivir Rivers. Living benthic foraminiferal faunas were characterised in detail and associations of the most abundant species classified into four groups. These groups allowed the characterisation of the modern environment and associated physical and chemical parameters.
Sedimentological and benthic foraminiferal analyses, combined with a chronological framework, were performed on three Holocene sequences from contrasting sedimentary environments. A comparison between the most abundant species in living faunas and those occurring in Holocene shelf sediments was also made. An integrated approach using all the available information sheds light on paleoenvironmental changes, which were distilled into five evolutionary scenarios. Each scenario is a representation of the environmental changes that characterised the three studied environments at different times during the Holocene. They indicate the influence of various factors, including sea-level changes, river discharge and substrate properties, that are mirrored by different benthic foraminiferal assemblages.
The thesis demonstrates that the modern ecological preferences of common benthic foraminiferal species have direct applicability to interpreting the palaeoenvironmental record.
iii
Resumo alargado
Esta tese apresenta uma investigação sobre os foraminíferos bentónicos como indicadores paleo-ambientais na plataforma continental do Norte do Golfo de Cádis, nordeste do Oceano Atlântico.
Os foraminíferos bentónicos constituem um dos grupos da meio-fauna protista mais abundantes. A sua distribuição e abundância são influenciadas por parâmetros físicos, químicos e biológicos, o que os torna excelentes ferramentas para interpretações ecológicas e ambientais. A sua carapaça biomineralisada possui a capacidade de ser preservada no registo fóssil, podendo fornecer informações credíveis para compreender as alterações em ambientes marinhos, ocorridas no passado histórico e geológico. Nas últimas décadas o seu estudo tem sido aplicado nas áreas da ecologia, biogeografia, monitorização ambiental, paleoecologia, paleoceanografia e reconstruções bioestratigráficas. No entanto, interpretações exactas do registo fóssil estão dependentes do conhecimento sobre a ecologia das espécies, preferencialmente das que vivem na mesma área. Assim, é essencial obter novas informações sobre a distribuição no espaço e tempo das espécies de foraminíferos bentónicos e avaliar a sua relação com os parâmetros ambientais. Isto com o intuito de inferir o seu habitat passado, compreender as alterações ambientais ocorridas e melhorar a calibração destas espécies como indicadores (“proxies”) de parâmetros que não podem ser medidos directamente. Este conhecimento é importante não só para compreender o passado, mas também para prever futuras tendências das alterações climáticas e oceanográficas.
As plataformas continentais são zonas de transferência entre os ambientes terrestres e o oceano. As sequências sedimentares destas áreas são importantes arquivos ambientais, onde ficam registados os processos naturais, tais como variações do nível médio do mar, alterações climáticas e oceanográficas, e o impacto antrópico ocorrido durante o Holocénico. A plataforma continental do norte do Golfo de Cádis (SW da Península Ibérica) é uma área com
elevado potencial para o estudo e compreensão destes processos. Esta área recebe as descargas do Rio Guadiana, que é a principal fonte sedimentar da região. A bacia hidrográfica do Rio Guadiana é ainda altamente susceptível a mudanças climáticas, influenciada por fortes variações inter-anuais e variações em larga escala, nomeadamente a Oscilação do Atlântico Norte. As variações climáticas e o aumento da actividade antrópica nesta área alteraram significativamente o sistema durante o Holocénico, modificando o tipo e a quantidade de sedimentos exportados para a plataforma continental. Na área da plataforma a evolução pós-glacial dos sistemas deposicionais foi fortemente influenciada pela rápida elevação do nível médio do mar que levou à formação de depósitos transgressivos e de alto nível do mar (“highstand system tracts”).
Na plataforma norte do Golfo de Cádis, a maioria dos estudos sobre foraminíferos bentónicos tem-se focado nas associações totais de foraminíferos bentónicos em amostras de sedimentos superficiais, que incluem as faunas vivas e as associações subfósseis. Em oposição, existem ainda poucos estudos sobre a distribuição das associações de foraminíferos bentónicos durante o Holocénico. Além disso, a realização de estudos em ambientes de plataforma que compare os padrões de distribuição entre faunas vivas e ocorridas durante o Holocénico, essenciais para a verificação de foraminíferos bentónicos como “proxies”, são ainda escassos na Europa e inexistentes na plataforma continental do norte do Golfo de Cádis.
Para atingir o objectivo principal deste trabalho - investigar os foraminíferos bentónicos como indicadores ambientais e sua aplicação em reconstruções paleoambientais durante o Holocénico, na plataforma continental do norte do Golfo de Cádis - vários objectivos específicos foram formulados: 1) ampliar o conhecimento sobre a ecologia das faunas vivas de foraminíferos bentónicos em amostras de sedimentos superficiais, através do reconhecimento das associações de espécies, o seu habitat e sua relação com os parâmetros físico-químicos; 2) compreender as alterações paleoambientais decorridas durante o
v Holocénico com base no estudo sedimentológico e na caracterização das associações de foraminíferos bentónicos, em três ambientes distintos de deposição sedimentar na plataforma continental; 3) Comparar as associações de foraminíferos bentónicos vivos (amostras de sedimentos superficiais) com as associações fósseis (testemunhos verticais), a fim de: a) analisar os efeitos tafonómicos; b) analisar as relações entre as associações fósseis e os sedimentos em que estas ocorrem, de forma a compreender as relações entre as espécies e o seu habitat; c) reconstruir as condições ambientais ocorridas no passado, usando os foraminíferos bentónicos como “proxies”; d) compreender as alterações paleoambientais ocorridas durante o Holocénico, relacionadas com as alterações do nível médio do mar, as alterações climáticas e o impacto antrópico.
Quarenta e sete amostras de sedimentos superficiais foram analisadas para o estudo das faunas de foraminíferos bentónicos vivos, colhidas na plataforma continental entre as desembocaduras dos Rios Guadiana e Guadalquivir, usando uma draga Smyth McIntyre. Foram ainda analisadas 14 amostras, provenientes dos primeiros centímetros do topo dos testemunhos verticais, colhidos pelo método de vibração. A amostragem estendeu-se entre os 10,7 e os 115,5 m de profundidade. Em ambas as amostragens, a camada superficial foi amostrada, e guardada num frasco contendo uma solução de etanol a 95% e corante Rosa de Bengal (1 g/l). Após a lavagem das amostras, a fracção >63 μm foi analisada à lupa binocular. Sempre que possível, um número mínimo de 300 indivíduos corados foram retirados de cada amostra, colados em porta-foraminíferos, identificados e contados. Foram calculadas as abundâncias relativas das espécies, calculada a densidade populacional (número de indivíduos por 10 cm3 de sedimento), a riqueza específica, e os índices de diversidade de Shannon, equitabilidade e alfa Fisher. Foram ainda efectuadas análises de estatística multivariada, usando as espécies com abundância relativa superior a 5%, nomeadamente a Análise de Clusters (CA) e a Análise das Componentes Principais (PCA). A distribuição das espécies
mais abundantes foi ainda relacionada com dados colhidos na área de estudo, relativos à temperatura, salinidade, matéria particulada em suspensão e dados relativos à produção primária provenientes da literatura.
Os três testemunhos verticais analisados foram colhidos na plataforma continental adjacente ao Rio Guadiana. O testemunho vertical 5, com 353 cm de comprimento foi colhido na zona central do corpo lodoso, a 72 m de profundidade. O testemunho vertical 7, com 71,5 cm de comprimento, foi colhido a 36 m de profundidade, na proximidade de um depósito arenoso transgressivo. O testemunho vertical 8, com 376 cm de comprimento, foi extraído do prodelta do Guadiana a 22 m de profundidade. Ao longo dos 3 testemunhos foram efectuadas 11 datações de radiocarbono (14C) pelo método de AMS (Accelerated Mass Spectrometry).
As idades obtidas foram calibradas usando a correcção do efeito de reservatório global dos oceanos e o efeito reservatório local. No testemunho vertical 5 foram ainda realizadas duas datações pelo método da racemisação de aminoácidos (AAR). Análises sedimentológicas foram efectuadas ao longo dos 3 testemunhos estudados, nomeadamente análises granulométricas realizadas sequencialmente e análises à lupa binocular dos componentes da fracção arenosa. As associações de foraminíferos bentónicos fósseis foram analisadas e realizadas análises de clusters usando as espécies com abundância superior a 5%.
Foi efectuada a caracterização detalhada das faunas de foraminíferos bentónicos vivos, na plataforma continental entre as desembocaduras dos Rios Guadiana e Guadalquivir. Os resultados demonstraram que a densidade populacional é geralmente influenciada pelas descargas dos rios, particularmente associada a maiores concentrações de matéria particulada em suspensão e sedimentos finos. A riqueza específica revelou ser influenciada pelo tipo de sedimento. As associações de foraminíferos bentónicos obtidas a partir das espécies mais abundantes revelaram uma clara relação com os parâmetros físico-químicos do meio. Foi
vii assim possível a obtenção de quatro grupos de espécies, que caracterizam o seu ambiente actual:
O grupo 1, é representado pelas espécies Bolivina ordinaria e Hopkinsina atlantica, consideradas as mais oportunistas da associação. A sua abundância está associada às descargas dos rios, ao elevado fornecimento de matéria orgânica, a sedimentos finos e aos limites de distribuição da produtividade primária. A abundância de Hopkinsina atlantica foi também associada com a elevada produção, resultante das descargas do Rio Guadalquivir.
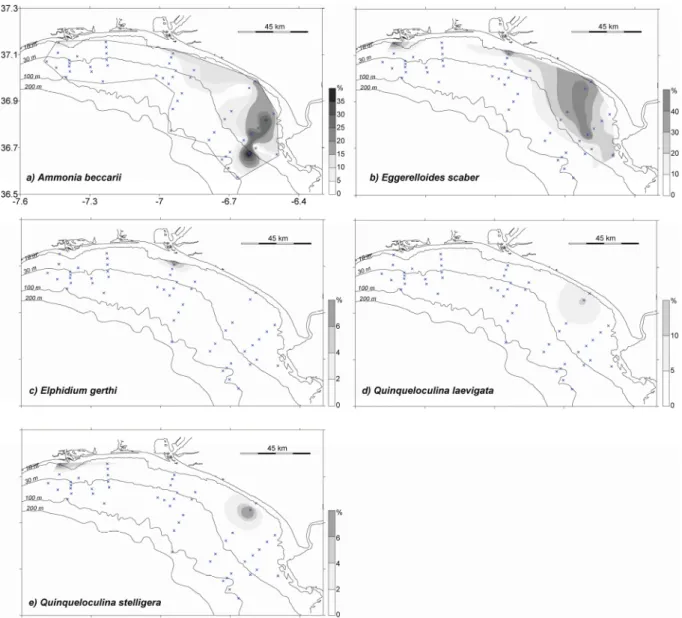
O grupo 2 é dominado por Ammonia beccarii, Eggerelloides scaber, Elphidium gerthi, Quinqueloculina laevigata e Quinqueloculina stelligera, que apresentaram maiores abundâncias em águas pouco profundas. Estas espécies foram ainda associadas a diferentes tipos de sedimento e a sua distribuição influenciada pelas descargas dos rios de diferente forma.
O grupo 3 é dominado por Bulimina aculeata, Elphidium excavatum, Bulimina elongata, Epistominella vitrea, Rectuvigerina phlegeri e Elphidium culvilliere. Estas espécies apresentaram maiores abundâncias entre os 30 e os 100 m de profundidade. Ocorrem principalmente em sedimentos lodosos, associados a reduzidos níveis de hidrodinâmica, a baixos níveis de oxigénio e altos teores de matéria orgânica.
O grupo 4 inclui as espécies que ocorrem com maior abundância a profundidades superiores. Este grupo foi dividido no Sub-grupo 4A (Bolivina catanensis, Cassidulina laevigata, Stainforthia sp., Nouria polymorphinoides, Bolivina italica, Bolivina striatula, e Nouria sp.) que apresentou maiores abundâncias em amostras seleccionadas e o Sub-grupo 4B (Brizalina dilatata, Brizalina spathulata, Bulimina marginata, Nonionella iridea, Nonionella stella e Nonionella turgida) em que as espécies apresentaram uma distribuição irregular em áreas distintas da plataforma. A maior abundância de espécies do género
Nonionella foi também relacionada com valores inferiores de temperatura e salinidade. A abundância da espécie N. iridea foi ainda associada a filamentos de “upwelling”.
As alterações paleoambientais ocorridas durante o Holocénico foram investigadas em três ambientes marinhos distintos da plataforma continental adjacente ao Rio Guadiana: o corpo lodoso localizado na plataforma média (core 5), que registou os últimos ca. 11.500 Cal anos BP; o depósito transgressivo localizado na plataforma média superior (core 7), registando os últimos ca. 10.400 Cal anos BP; e o prodelta do Guadiana (core 8) localizado na plataforma interna que registou os últimos ca. 5.000 Cal anos BP. Estes ambientes diferem em termos de profundidade, regime sedimentar e associações de foraminíferos bentónicos. No entanto, foi observado nos três ambientes que a fracção arenosa era predominantemente de origem fluvial. As análises sedimentológicas e dos foraminíferos bentónicos foram efectuadas em cada registo sedimentar e integradas num quadro cronológico. Foram identificados quatro estágios evolutivos (descritos posteriormente), que fornecem informações sobre os padrões de deposição sedimentar ocorridos durante o Holocénico, em resposta às alterações do nível médio do mar, às descargas do Rio Guadiana, às alterações climáticas e ao impacto antrópico.
As análises efectuadas no testemunho vertical 7 colhido junto ao depósito arenoso transgressivo, indicam que este é um ambiente complexo e requer a realização de mais estudos, no sentido de melhor compreender a sua formação e evolução.
A comparação entre as faunas de foraminíferos bentónicos vivos e associações de mortos/fósseis, na plataforma continental adjacente ao Rio Guadiana, revelaram que, em média, 95% das faunas vivas foram preservadas no registo fóssil. A comparação entre as associações de foraminíferos vivos e as totais, revelaram maior abundância de indivíduos vivos das espécies B. ordinaria e E. excavatum em Fevereiro, indicando um período reprodutivo coincidente com o período de amostragem. No entanto, dependendo do ambiente da plataforma analisado, para além dos períodos reprodutivos, outros factores como a
ix profundidade, o tipo de sedimento e a quantidade e qualidade de alimento disponível, revelaram ter um papel importante na abundância das espécies R. phlegeri, B. dilatata, N. iridea, N. stella e N. turgida. No corpo lodoso situado na plataforma média, a abundância destas espécies foi principalmente influenciada por fenómenos de “upwelling” local. No entanto, para validar estas conclusões, mais estudos usando amostras sem perturbação dos sedimentos superficiais para a comparação entre as associações de organismos vivos e totais são requeridos.
Os processos que ocorrem após a morte do organismo (post-mortem), tais como a destruição e dissolução das carapaças, parecem ser responsáveis pelas diferenças encontradas entre as associações de organismos vivos e mortos/fósseis. Estes processos parecem justificar a maior abundância das espécies B. ordinaria, R. phlegeri, B. dilatata, N. iridea e N. stella nas associações de foraminíferos vivos comparativamente às associações de mortos ou fósseis. Além disso, estas espécies apresentaram menores abundâncias nos testemunhos verticais do que nas amostras de sedimentos superficiais. No entanto, estas espécies foram também as mais abundantes em pelo menos um dos três ambientes estudados durante o Holocénico, indicando que estas espécies podem ser usadas como indicadores paleoambientais. As variações das espécies de foraminíferos bentónicos mais abundantes podem ser consideradas indicadores úteis das alterações ambientais ocorridas durante o Holocénico em ambientes de plataforma.
A abundância das espécies E. scaber, E. gerthi and Q. stelligera é controlada pela profundidade, preferindo águas pouco profundas, e pelas propriedades do substrato. A espécie A. beccarii ocorreu em todos os ambientes com abundância reduzida. A espécie E. scaber pode ser usada como indicadora do aumento da matéria orgânica na plataforma, devido à sua associação com sedimentos finos, baixos níveis hidrodinâmicos e tolerância a elevadas
concentração de metais pesados. As maiores abundâncias de E. gerthi and Q. stelligera parecem estar associadas a ambientes de águas pouco profundas e sedimentos grosseiros.
As abundâncias das espécies Bulimina aculeata, B. elongata, C. laevigata e B. spathulata são controladas pela profundidade da coluna de água, por baixos níveis de hidrodinâmica e por sedimentos finos. A abundância de Bulimina aculeata foi ainda controlada pela ocorrência de sedimentos finos e reduzidos níveis de oxigénio, em águas pouco profundas. As espécies C. laevigata e B. spathulata apresentaram o seu nicho ecológico em zonas mais profundas da plataforma. Durante o Holocénico a sua distribuição foi possivelmente controlada pela subida do nível médio do mar e aumento do teor de silte e argila nos sedimentos.
A abundância das espécies A. mamilla e P. mediterranensis, associada a sedimentos grosseiros, pode ser considerada um indicador do nível médio do mar mais baixo, isto é, localizado numa posição inferior à actual. A combinação entre a subida do nível médio do mar e a competição entre espécies parecem controlar a abundância da S. fusiformis, a qual desapareceu após o nível médio do mar ter atingido a sua posição actual. A abundância de Q. rugosa foi também controlada pela subida do nível médio do mar, e pode ser indicativa de um ambiente arenoso restrito a zonas pouco profundas.
Uma abordagem integradora, usando toda a informação disponível, permitiu clarificar as alterações peloambientais ocorridas na plataforma continental adjacente ao Rio Guadiana durante o Holocénico, apresentando cinco cenários evolutivos. Esta integração revelou um cenário evolutivo adicional, que se seguiu à estabilização do nível médio do mar (entre ca. 5.000 e 4.500 Cal anos BP), detalhando assim os estágios evolutivos definidos anteriormente.
O primeiro cenário evolutivo, entre ca. 11.500 e ca. 10.000 Cal anos BP, corresponde a um período de subida do nível médio do mar, resultando na deposição de sedimentos finos e na redução dos níveis de hidrodinâmica em áreas mais profundas da plataforma. O aumento
xi na abundância das espécies B. ordinaria e R. phlegeri indica ainda um aumento de matéria orgânica de origem terrestre. As abundâncias relativamente elevadas de N. iridea e N. stella indicam também produtividade elevada durante este período.
Entre ca. 10.000 e ca. 5.000 Cal anos BP, as descargas do rio e o fornecimento de sedimentos terrígenos para a plataforma diminuíram, facto relacionado com a subida do nível médio do mar para uma posição próxima da actual e consequente aumento da distância à foz do rio.
Após a estabilização do nível do mar que terá ocorrido entre ca. 5.000 e 4.500 Cal anos BP, a maior abundância de E. vitrea, B. dilatata e B. marginata indica o estabelecimento de novos nichos ecológicos em áreas de maior profundidade e menor influência das descargas do rio. A maior abundância de B. dilatata, C. laevigata e R. phlegeri indica ainda um curto período de forte “upwelling”.
As abundâncias relativamente constante das espécies de foraminíferos entre os ca. 5.000 - 4.500 e os ca. 1.000 Cal anos BP, indicam condições ambientais relativamente estáveis, que se seguiram após o estabelecimento dos nichos ecológicos a maiores profundidades, de acordo com a estabilização do nível médio do mar, os reduzidos níveis de hidrodinâmica e o elevado teor de matéria orgânica de origem marinha e terrestre.
Os últimos ca. 1.000 anos reflectem as condições actuais de deposição, que se estabeleceram após a formação do prodelta na plataforma interna e a formação do corpo lodoso na plataforma média. Ocorrem eventos de “upwelling” sazonal na plataforma média com consequente aumento da produção primária. Este período reflecte ainda o impacto antrópico na bacia hidrográfica do Guadiana através da intensificação do uso do solo, da actividade mineira, da construção de barragens e da desflorestação, e as alterações climáticas, como o inicio da Pequena Idade do gelo e a relação com a fase negativa da Oscilação do Atlântico Norte.
Esta tese demonstra portanto que as preferências ecológicas das espécies actuais de foraminíferos bentónicos mais abundantes, têm aplicação directa na interpretação do registo paleoambiental na plataforma continental adjacente ao Rio Guadiana.
xiii
Para o Paulo, a Camila e
xv
Acknowledgments
This is my thesis, its achievement was only possible with the help and collaboration of many people. Simple words cannot transmit all the help and assistance they gave me throughout the years. Without them, this thesis would certainly be different.
My deep gratitude to my internal supervisors, Prof. Alveirinho Dias and Prof. Óscar Ferreira for their invaluable support during the past years. They have given me the opportunity to grow as a person and researcher.
A very special thanks to my external supervisor, Dr. Joachim Schönfeld, for all his support, incentive, and professional and scientific advice. His teaching helped me to better understand the world of benthic foraminifera.
I want to express my gratitude to Ramon González and Paco Lobo for their support and endurance over the last few years. I feel lucky for having known you both.
I also want to thank all the CIACOMAR team members involved in the collection of the surficial samples during the SIRIA project, and in the collection and sampling of the sediment cores from CRIDA project campaigns: Ramon, Carla, Amélia, Paco, Sandrinha, Alexandre, Julio Cunha, Margarida, Patrícia, Filipa and Ana Matias. I hope I didn´t forget anyone!
My gratitude to our Instituto Tecnológico e Nuclear (ITN) partners in the CRIDA project, for their collaboration in the CRIDA campaigns and valuable data discussions. Special thanks, amongst others, Dra. Fátima Araújo, Catarina Corredeira and Didier Burdloff.
I would like to thank to everyone who helped me with taxonomic issues, especially to my supervisor Dr. Schönfeld and also Dr. Perfecto Villannueva Guimerans (University of Cadiz).
I wish to thank Ramon Gonzalez and Patrícia Silva for the counting and identification of sand grains for the sedimentological study and also to Sr. Cunha and Margarida for grain size analyses.
Furthermore, I am very grateful to the people who gave me advice, assistance and discussion in all manner of areas. In particular, to Eng. Monge Soares (ITN) for the helpful discussion about age calibration and local reservoir effect, and to Dr. Francisco Fatela (Faculdade de Ciências da Universidade de Lisboa) for providing facilities for scanning electron microscope photography.
I want to thank all my colleagues from CIACOMAR/CIMA who, during the past few years, gave me their constant help, encouragement and friendship. In particular, to Ana Matias, Francisca, Rita, Margarida, Sr. Cunha, André, Ricardo, Carlos, Sarita and Simon.
Last, but not least, a very special thanks to Paulo and Camila, who share their life with me and fill me with happiness. To my parents, Fernando and Fernanda, for everything they did for me all over these years; to my sister Sandra for all her help and support; and to my grandmother Laurinda. You make me feel much loved. This acknowledgment extends also to my other family and friends, who have, in different ways, always supported me and encouraged to go further.
INSTITUTIONAL AND FINANCIAL SUPPORT:
The work was funded by:
• Fundação para a Ciência e a Tecnologia (FCT), PhD grant reference SFRH/BD/18342/2004.
• Instituto do Mar, grant reference IMAR/Plano Geral de Monitorização/I.
• Project CIRCO (Climate changes from isotopic records during the Holocene in South-western Iberia) from the FCT (PTDC/CLI/66393/2006), research grant.
xvii This work benefited from the following scientific projects:
• SIRIA (Situação de Referência da Região Costeira Algarvia Influenciável pela Barragem do Alqueva) from Ministério da Defesa Nacional e Fundação das Universidades Portuguesas.
• CRIDA (Consequences of river discharge modifications on coastal zone and continental shelf) from the FCT (POCTI/P/MAR/15289/99).
• IMCA (Impact of climatic and anthropic variations on the Northern continental shelf, Gulf of Cadiz) from the FCT (POCI/CLI/60192/2004).
xix
Contents
Resumo i Abstract ii Resumo alargado iii Acknowlegments xv Contents xix List of figures xxiii List of tables xxviii
1. Introduction 1
1.1. Motivation for this study 2 1.2. Aims and objectives 4 1.3. Structure of the thesis 5
2. Study area
7
2.1. Location 8
2.2. Climatic characteristics 8 2.3. Oceanographic setting 10
2.3.1. Shelf current patterns 10 2.3.2. Tide and wave regime 12 2.4. Shelf bathymetry 12
2.5. Fluvial and sediment supply 13
2.6. Modern surficial sediment distribution 14 2.7. Holocene evolution 15
2.7.1. Continental shelf 15 2.7.2. Estuary infilling 16 2.8. Anthropogenic impact 17
3. Distribution of living benthic foraminifera 19
3.1. Introduction 203.2. Material and methods 22
3.2.1. Analyses of benthic foraminifera 22
3.2.2. Temperature, salinity and suspended particulate matter near sea floor 26 3.3. Results 27
3.3.1. Benthic foraminiferal community 27 3.3.1.1. Multivariate analyses 31
3.3.2. Individual distribution of benthic foraminifera 37 3.3.3. Hydrological parameters 42
3.3.3.1. Water temperature and salinity 42
3.3.3.2. Suspended particulate matter concentration 43 3.3.4. Primary productivity 45
3.4. Discussion 47 3.5. Conclusions 57
4. Holocene paleoenvironmental changes on the continental shelf off the
Guadiana River
61
4.1. Introduction 62
4.2. Material and methods 65 4.2.1. Sediment cores 65
4.2.2. AMS radiocarbon dating 66 4.2.2.1. Age model 67
4.2.3. Grain size 67
4.2.4. Components of sand fraction 68 4.2.5. Benthic foraminifera 69
4.3. Results 70
4.3.1. Age models 70
4.3.2. Sediment characteristics 72 4.3.3. Benthic foraminifera 76
4.3.3.1. Core 5: Middle shelf mud body 76
4.3.3.2. Core 7: Upper middle shelf transgressive bulge 79 4.3.3.3. Core 8: Inner shelf prodeltaic wedge 83
4.3.3.4. Comparison between environments 86 4.4. Discussion 89
4.4.1 Evolutionary stages 89
4.4.1.1. From ca. 11500 to ca. 10000 Cal yr BP 90 4.4.1.2. From ca. 10000 to ca. 5000 Cal yr BP 91 4.4.1.3. From ca. 5000 to ca. 1500-1000 Cal yr BP 95
xxi 4.5. Conclusions 103
5. Comparison between recent (living) and Holocene distribution of benthic
foraminifera 109
5.1. Introduction 110
5.2. Material and methods 112
5.2.1. Surficial benthic foraminiferal distribution 112
5.2.2. Benthic foraminiferal distribution in sediment cores 112 5.2.3. Comparison between the distribution of benthic foraminifera in surficial samples and sediment cores 113
5.3. Results 114
5.3.1. Living species that showed different abundance in the vicinity of core locations 115
5.3.1.1. Species with higher abundance in the vicinity of river outflow 115 5.3.1.2. Species with higher abundance between 30 and 100 m water depth 118 5.3.1.3. Species with higher abundance in deep areas 120
5.3.2. Living species without variation in the vicinity of core locations 125 5.3.2.1. Species with high abundance in shallow waters 125
5.3.2.2. Species with high abundance below 30 m water depth 127 5.3.3. Species abundant only in sediment cores 129
5.4. Discussion 132
5.4.1. Species richness 132
5.4.2. Living versus total assemblages in the top of the cores 133 5.4.3. Living versus dead/fossil assemblages 136
5.4.4. Dead/fossil versus living assemblages without variation in the vicinity of core locations 144
5.4.5. Dead/fossil assemblages with low representation in the living fauna 147 5.4.6. Shelf evolutionary scenarios 148
5.5. Conclusions 151
6. General conclusions 155
7. Taxonomy
161
xxiii
List of figures
2.1 – Location map of the study area in the Gulf of Cadiz. ... 8 2.2 – Modern surficial sediment distribution of the northern Gulf of Cadiz continental
shelf, between the Guadiana and Guadalquivir Rivers (adapted from Gonzalez et al., 2004)... 15 3.1 – Location of the samples studied, including the collection method and
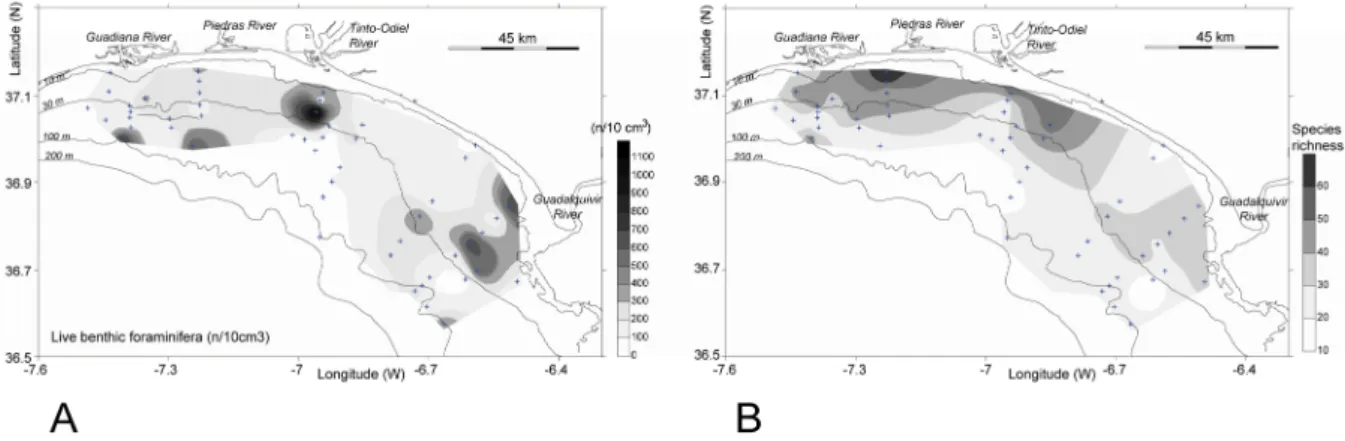
characteristics of surface sediments on the continental shelf of the study area (adapted from Gonzalez et al., 2004). ... 24 3.2 – Distribution of: A - living benthic foraminifera, in number of specimens (n) per
10 cm3,and B - species richness in the study area. ... 28 3.3 – Variation of the number of living benthic foraminifera per 10 cm3 and species
richness with water depth... 29 3.4 – Variation in diversity indices with water depth: a) Shannon index; b) Equitability
index; c) Alpha-Fisher index. Average values are represented by a dashed line... 29 3.5 – Variation of species richness with the number of specimens counted in each
analysed sample and the respective logarithmic trend lines: A – dataset including all the analysed samples; B – dataset including only the samples used in statistical analyses. ... 30 3.6 – Variation of species richness (E100) with water depth. Average and standard
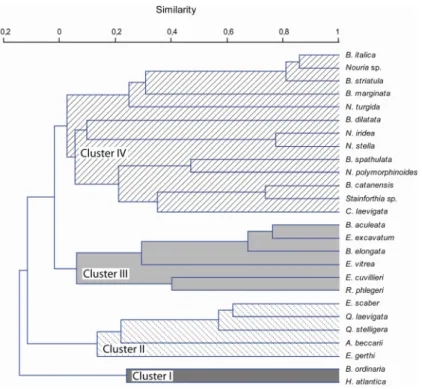
deviation obtained for 10 m water-depth intervals: A – dataset including all the analysed samples; B – dataset including only the samples used for statistical analyses. ... 31 3.7 – Dendrogram resulting from R-mode cluster analyses (correlation method joined
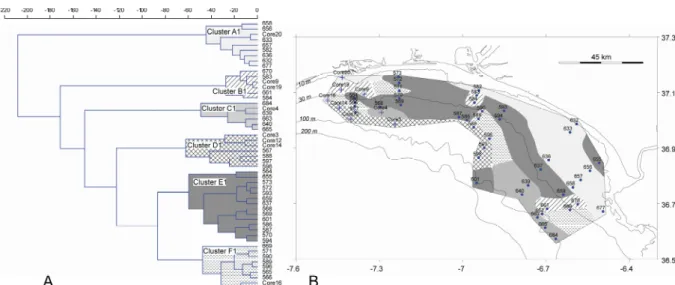
by UPGMA) based on the 26 species with total abundance >5%... 32 3.8 – Spatial distribution of the four biofacies recognised by R-mode cluster analyses,
3.9 – Q-mode cluster analysis (Bray-Curtis measure joined by UPGMA) of the samples based on a total abundance >5%. A- Dendrogram with four clusters recognised. B- Spatial distribution of the four clusters across the continental shelf... 34 3.10 – Principal Components Analysis result (components 1 and 2) based on species
with total abundance >5%... 35 3.11 – Dendrogram resulting from R-mode cluster analyses (Ward’s method) based on
16 species that occurred in at least half of the analysed samples (26). ... 36 3.12 – Q-mode cluster analysis (Ward’s method) of the samples based on 16 species
that occurred in at least half of the analysed samples (26). A- Dendrogram with six clusters recognised. B- Distribution of the six clusters across the continental shelf. ... 37 3.13 – Distribution of species most abundant in areas under the direct influence of
rivers outflow. a) Bolivina ordinaria; b) Hopkinsina atlantica. ... 38 3.14 – Distribution of species most abundant in shallow waters. a) Ammonia beccarii;
b) Eggerelloides scaber; c) Elphidium gerthi; d) Quinqueloculina laevigata; e) Quinqueloculina stelligera... 39 3.15 – Distribution of species most abundant between 30 and 100 m water depth. a)
Bulimina aculeata; b) Elphidium excavatum; c) Bulimina elongata; d) Epistominella vitrea; e) Rectuvigerina phlegeri; f) Elphidium cuvillieri. ... 40 3.16 – Species with isolated high abundances in a few samples. a) Bolivina catanensis;
b) Cassidulina laevigata; c) Stainforthia sp.; d) Nouria polymorphinoides; e) Bolivina italica; f) Bolivina striatula; g) Nouria sp... 41 3.17 – Species with a patchy distribution. a) Brizalina dilatata; b) Brizalina spathulata;
c) Nonionella iridea; d) Nonionella stella; e) Nonionella turgida; f) Bulimina marginata. ... 42 3.18 – Temperature distribution near sea floor, measured during the period of surficial
xxv 3.19 – Salinity distribution near the sea floor, measured during the same period of
surficial sediment collection... 44 3.20 – Suspended particulate matter concentrations (mg/l) distributed near the sea floor,
calculated for water samples collected during the same period of surficial sediment collection... 44 3.21 – Primary productivity in the study area, obtained from the advection nitrogen flux
to the photic zone (Ruiz and Navarro, 2006) and chlorophyll concentrations in surface waters (Navarro and Ruiz, 2006)... 45 3.22 – Comparison of areas of high primary productivity in the study area with the
surficial distribution of individual benthic foraminiferal species: a) A. beccarii, b) E. scaber, c), B. ordinaria, d), R. phlegeri, e), H. atlantica, f) N. iridea... 46 4.1 – Location of the studied cores (5, 7 and 8) and distribution of surface sediments on
the continental shelf off the Guadiana River (adapted from Gonzalez et al., 2004)... 68 4.2 – Age model for cores 5, 7 and 8, based on calibrated ages with error bars, and
sedimentation rates between age control points... 72 4.3 – Grain size (A) and mean grain size (B) variations with age (Cal yr BP) for the
three environments analysed on the Guadiana continental shelf: a) middle shelf mud body; b) upper middle shelf transgressive bulge; c) inner shelf prodeltaic wedge. ... 74 4.4 – Variation of the components of sand fraction with age (Cal yr BP) for the three
environments analysed on the Guadiana continental shelf: a) middle shelf mud body; b) upper middle shelf transgressive bulge; c) inner shelf prodeltaic wedge. ... 75 4.5 – Variation with age in core 5 of: number of benthic foraminifera per gram of dry
sediment, species richess, Shannon index diversity, relative abundances of benthic foraminiferal species, with abundance >5% in at least one sample and proposed faunal assemblages zonation. ... 78 4.6 – Dendrogram classification resulting from cluster analyses based on 17 species
produced by R-mode analysis (correlation method joined by UPGMA); b) Samples associated produced by Q-mode analysis (Ward’s method)... 79 4.7 – Variation with age (Cal yr BP) in core 7 of: number of benthic foraminifera per
gram of dry sediment, species richess, Shannon index diversity, relative abundances of benthic foraminiferal species, with abundance >5% in at least one sample and proposed faunal assemblages zonation. ... 81 4.8 – Dendrogram classification resulting from cluster analyses based on 11 species
with relative abundance >5% in at least one sample: a) Foraminiferal assemblages produced by R-mode analysis (correlation method joined by UPGMA); b) Samples associated produced by Q-mode analysis (Ward’s method)... 83 4.9 – Variation with age (Cal yr BP) in core 8 of: number of benthic foraminifera per
gram of dry sediment, species richess, Shannon index diversity, relative abundances of benthic foraminiferal species, with abundance >5% in at least one sample and proposed faunal assemblages zonation. ... 85 4.10 – Dendrogram classification resulting from cluster analyses based on 12 species
with relative abundance >5% in at least one sample: a) Foraminiferal assemblages produced by R-mode analysis (correlation method joined by UPGMA); b) Samples associated produced by Q-mode analysis (Ward’s method)... 86 4.11 – Variation with age (Cal yr BP) of: a) benthic foraminifera per gram of dry
sediment; b) species richness, c) Shannon Index. A, B and C represent the three environments analysed on the Guadiana continental shelf as: A- middle shelf mud body; B- upper middle shelf transgressive bulge and C- inner shelf prodeltaic wedge. ... 87 4.12 – Variation with age (Cal yr BP) of relative abundances of benthic foraminiferal
species, with abundance >5% in at least one sample, for the three environments analysed on the Guadiana continental shelf: a) middle shelf mud body; b) upper middle shelf transgressive bulge, c) inner shelf prodeltaic wedge. The results obtained by R-mode (Cluster I and II) and by Q-mode (Cluster A and B) cluster analyses, when consistent, are also represented. The dash lines indicate the results obtained by zonation in each environment... 88
xxvii 4.13 – Sea-level curves, proposed by different authors with respective location.
Variation of depth below present sea level with: a) radiocarbon ages (Ky BP); b) calibrated age (Cal Ky BP)... 95 5.1 – Location of the surface samples and sediment cores on the continental shelf off
the Guadiana River. The grey area limited by the dashed line indicates nitrogen flux to the photic zone (adapted from Ruiz and Navarro, 2006)... 114 5.2 – Relative abundance of Bolivina ordinaria: a) surficial distribution; b) record in
sediment cores. ... 116 5.3 – Relative abundance of Hopkinsina atlantica: a) surficial distribution; b) record in
sediment cores. ... 117 5.4 – Relative abundance of Elphidium excavatum: a) surficial distribution; b) record in
sediment cores. ... 118 5.5 – Relative abundance of Epistominella vitrea: a) surficial distribution; b) record in
sediment cores. ... 119 5.6 – Relative abundance of Rectuvigerina phlegeri: a) surficial distribution; b) record
in sediment cores... 120 5.7 – Relative abundance of Brizalina dilatata: a) surficial distribution; b) record in
sediment cores. ... 121 5.8 – Relative abundance of Bulimina marginata: a) surficial distribution; b) record in
sediment cores. ... 122 5.9 – Relative abundance of Nonionella iridea: a) surficial distribution; b) record in
sediment cores. ... 123 5.10 – Relative abundance of Nonionella stella: a) surficial distribution; b) record in
sediment cores. ... 124 5.11 – Relative abundance of Nonionella turgida: a) surficial distribution; b) record in
5.12 – Relative abundance variation with age (Cal yr BP) along cores 5, 7 and 8 from species with high abundance in shallow waters. ... 127 5.13 – Relative abundance variation with age (Cal yr BP) along cores 5, 7 and 8 from
species with high abundance below 30 m water depth. ... 129 5.14 – Relative abundance variation with age (Cal yr BP) along cores 5, 7 and 8 from
species more abundant only in sediment cores. ... 130 5.15 – Schematic representation of the five shelf evolutionary scenarios (A to E)
identified off the Guadiana River, during the Holocene. ... 150
List of tables
4.1 – AMS Radiocarbon data from cores 5, 7 and 8. AAR ages data from core 5 (Asp aspartic acid, Glu glutamic acid)... 71
Chapter 1
1.1. Motivation for this study
Foraminifera constitute the most abundant group of shelled microorganisms in modern marine environments. They are generally small (<1 mm) although some can exceed few centimetres and most have a test which may be preserved in the fossil record (Murray, 2006). There are 45 living planktonic species recognised (Hemleben et al., 1989). The overwhelming majority of modern Foraminifera are benthic. They have greater diversity and a much longer geological record than planktonic foraminifera. The oldest fossil species are of Cambrian age, whereas the planktonic foraminifera originated in the mid Jurassic (e.g. Sen Gupta, 1999a).
The first biological investigations of foraminifers began in the 19th century, however, only in the second half of the 20th century the ecology of Foraminifera became a major area of
study. Increasing interest in this field was driven by the realization that the distribution of Foraminifera in the present-day environment can provide reliable clues for understanding marine environmental changes in the historical past and geological record (e.g. Sen Gupta, 1999a). Benthic foraminifera are highly dependent on environmental conditions, and even subtle changes are mirrored by shifts in species composition and abundance (e.g. Murray, 2001).
Over the last decades, benthic foraminifera have found wide application in ecology, biogeographic, environmental monitoring, palaeoecological and palaeoceangraphic reconstructions, biostratigraphy and correlation. However, accurate interpretations of the fossil record depend largely on our knowledge of the ecology of living benthic foraminiferal species, preferably from the same area (e.g. Alve, 2003; Murray, 2006). Thus it is essencial to gather new information on the spatial and temporal distribution of living benthic foraminiferal species and their relationship with environmental parameters in order to infer past habitats, understand environmental changes and improve proxy calibration. This knowledge
1. Introduction
3 is important not only to understand the past, but likewise to predict future trends in climatic and oceanographic change.
Continental shelves are areas of transfer of terrestrial material to the ocean. Sedimentary sequences from these areas are important environmental archives, registering a number of natural processes such as sea-level, oceanographic and climatic changes, as well as recent anthropogenic impacts during the Holocene (e.g. Lesueur et al., 1996; Bauch et al., 2001; Evans et al., 2002; Oldfield et al., 2003). The northern Gulf of Cadiz continental shelf (SW Iberian Peninsula) is an area with considerable potential for studying and understanding these processes. The area receives discharge from Guadiana and Guadalquivir Rivers, the main regional sediment sources. Guadiana River basin itself is highly susceptible to climatic changes, influenced by strong seasonal variations and larger-scale annual and decadal signals, such as the North Atlantic Oscillation (Dias et al., 2004; Trigo et al., 2004). Climatic variations and the increase in human activity in the Guadiana catchment area have significantly altered the system (Fletcher et al., 2007), changing both the type and quantity of sediment exported to the continental shelf (Gonzalez et al., 2001). The postglacial evolution of depositional systems in this area was strongly influenced by rapid sea-level rise, leading to the formation of transgressive and highstand system tracts (Lobo et al., 2001, 2004).
The sediments on the continental shelf off the Guadiana River possess a relatively clear signature and origin (e.g. Gonzalez et al., 2004; 2007; Lobo et al., 2004). This facilitates the establishment of a relationship between environmental factors, sedimentary influx, and variations in benthic foraminiferal assemblages.
In the northern Gulf of Cadiz, most of the literature concerning benthic foraminifera has focused on total benthic foraminiferal assemblages, including living fauna and subfossil assemblages of empty tests (e.g. Galhano, 1963; Ubaldo and Otero, 1958; Levy et al., 1993; 1995; Villanueva, 2000; 2001; Villanueva and Canudo, 1998; 1999; 2008; Villanueva and
Cervera, 1998; 1999a; 1999b; Villanueva et al., 1999a; 1999b; González-Regalado et al., 2001; Mendes et al., 2004). In contrast, only a few attempts were made in this area to reconstruct the distribution of benthic foraminifera during the Holocene (e.g. Mendes et al., 2006; 2010). Furthermore, the comparison between living and Holocene distribution patterns, essential for verifying benthic foraminiferal proxies, is generally scarce in European shelf environments and has not yet been done on the Gulf of Cadiz continental shelf.
1.2. Aims and objectives
The main objective of this thesis is to determine the environmental indicator value of benthic foraminifera living on the northern Gulf of Cadiz continental shelf and apply this to Holocene palaeoenvironmental reconstruction. In order to achieve this overarching goal, several specific objectives were formulated, namely:
1. Increasing ecological information about living benthic foraminiferal faunas in surficial samples, through establishing of their species associations and their relationship to physical and chemical parameters in their habitats.
2. Understanding the Holocene paleoenvironmental changes based on sedimentological and benthic foraminiferal characterisation in three different sedimentary depositional environments on the continental shelf.
3. Compare living benthic foraminiferal assemblages with fossil assemblages in sediment cores, in order to:
a) evaluate taphonomic effects;
b) analyse associations between fossil assemblages and the sediments in which they occur to better understand species-environment relationships;
c) reconstruct past environmental conditions using benthic foraminifera as a proxy;
1. Introduction
5 d) understand the Holocene paleoenvironmental changes associated with sea-level
changes, climatic variations and anthropogenic impact.
1.3. Structure of the thesis
This thesis is organised into six main chapters, followed by a taxonomic compilation. Each of the chapters introducing new data includes its own introduction, material and methods, results, discussion and conclusions.
- Chapter 1 is a general introduction, where this sub-chapter is included, describes the
motivation to study benthic foraminifera on the northern Gulf of Cadiz continental shelf and the main objectives of this study.
- Chapter 2 provides general information concerning the study area.
- Chapter 3 presents an investigation of the distribution and ecology of living benthic
foraminiferal faunas, in the study area as a mean of accurately determining their value as proxies (addressing objective 1).
- Chapter 4 presents an integrated approach that links sedimentology, fossil
foraminiferal assemblages and radiocarbon dating to reveal Holocene paleoenvironmental changes in three distinct sedimentary environments on the continental shelf off the Guadiana River (objectives 2 and 3c).
- Chapter 5 integrates assemblages of the most abundant species from recent (Chapter
3) and Holocene (Chapter 4) shelf sediments to evaluate taphonomic effects, reconstruct palaeoenvironmental changes and propose an integrated model for the evolution of the Guadiana shelf area during the Holocene (addressing objective 3);
- Chapter 7 presents the taxonomy of foraminiferal taxa recorded on the northern
Gulf of Cadiz continental shelf, illustrated with scanning electron microscope and direct light microscope photographs.
- Chapter 8 provides a list of the bibliographic references cited in this thesis.
- Chapter 9 presents the appendices (A, B, C and D), which include tables with the
relative abundance of the total assemblages (taxa with more than one specimen), for all analysed samples.
Chapter 2
2.1. Location
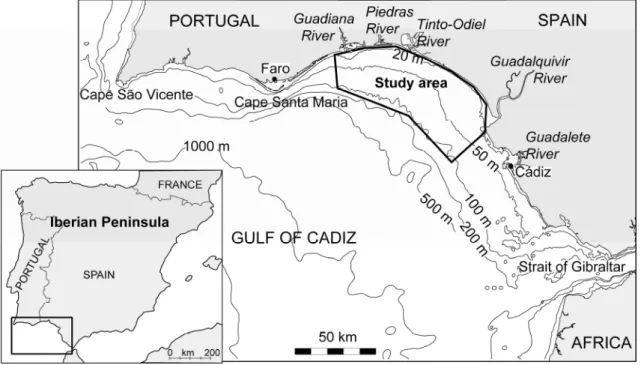
The Gulf of Cadiz is a wide basin located west of the Strait of Gibraltar, in the northeast Atlantic Ocean, between the Iberian Peninsula and northwest African coasts. The study area is located in the middle part of the northern Gulf of Cadiz continental shelf (south-western Iberian Peninsula) between the mouths of the Guadiana and Guadalquivir Rivers. In this area, the Guadiana River is a natural border between Portugal and Spain (Fig. 2.1).
The Gulf of Cadiz is a basin of relatively warm and oligotrophic waters that interrupts the fringe of cold and nutrient-rich upwelled water, during the upwelling season, dividing the North Atlantic Coastal province into the European and African sectors (e.g. Garcia-Lafuente and Ruiz, 2007).
Figure 2.1 – Location map of the study area in the Gulf of Cadiz.
2.2. Climatic characteristics
The study area is characterised by a Mediterranean climate, with hot, dry summers with at least two months of drought after the summer solstice, and mild winters during which the majority of the rain falls. The regional climate of the Guadiana River basin is classify as
2. Study area
9 semi-arid (Morales, 1995). As a consequence, the Guadiana run-off volume is subjected to high seasonal and inter-annual variations (e.g. Loureiro et al., 1986). Episodic floods play a major role in the supply of sediment from the river basin to the continental shelf (Morales, 1997; Portela, 2006).
These situations can be further aggravated by the influence of the North Atlantic Oscillation (NAO), which is the leading pattern of weather and climate variability over the Northern Hemisphere (e.g. Hurrell, 1995; Luterbacher et al., 2001). The NAO refers to a redistribution of atmospheric mass between the Arctic and the subtropical Atlantic, and swings from one phase to another producing large change in surface air temperature, winds, storminess and precipitation over the Atlantic, as well as the adjacent continents. The NAO also affects the ocean through changes in heat content, gyre circulations, mixed layer depth, salinity, high latitude deep water formation and sea ice cover (e.g. Hurrell and Deser, 2010).
A positive phase of the NAO reflects a below-normal pressure in the northern North Atlantic and an above-normal pressure over the central North Atlantic, leading to strong westerly winds associated with warm and moist air masses across the northern Europe during winter. On the western Iberian margin, this phase enhances upwelling conditions (e.g. Abrantes et al., 2005; Lebreiro et al., 2006; Bartels-Jónsdóttir et al., 2009). During a negative phase of the NAO, the atmospheric system changes, resulting in dry conditions at northern latitudes, and increases precipitation at the southern latitudes of Europe (Hurrell, 1995). Consequently on the Iberian Peninsula this phase results in more rainfall, and subsequent flooding in the river basin during winter mouths, also confirmed for the Guadiana River basin (Trigo et al., 2002; 2004; Dias et al., 2004).
The reconstruction of the climate history in the Guadiana valley based on pollen spectra indicates a warm, dry early Holocene phase between ca. 11790 and ca. 9000 Cal yr BP; a generally warm, humid phase, with prevalence of oceanic conditions from ca. 9000 to
ca. 5000 Cal yr BP; and a return to warm, dry conditions after ca. 5000 Cal yr BP (Fletcher et al., 2007).
2.3. Oceanographic setting
2.3.1. Shelf current patterns
The North Atlantic Central Water (NACW) is a representative water mass of the upper 1000 m of the water column. Criado-Aldeanueva et al. (2006) based on data from spring 2001, identified two surface coastal variations of the NACW, not considered as water mass: the Atlantic Water (SAW) and the Warm Shelf Waters (SW). SAW has it origin in the shallow NACW modified by air-sea interactions, and it was found between the surface and a depth of approximately 100 m over the entire region, except for the continental shelf. The SW has been detected mainly over the continental shelf, comes from the noticeably SAW influence by continental processes, including heating and fresh water inputs from land. To the northern part of the Gulf of Cadiz, the lower part of the NACW layer is entrained by Mediterranean Water towards the open ocean, while the upper layer forms part of the anticyclonic surface circulation of the Gulf of Cadiz (Criado-Aldeanueva et al., 2006).
In the Gulf of Cadiz the general surface circulation is anticyclonic with short-term, meteorologically induced variations (Criado-Aldeanueva et al., 2006). Recently studies showed a permanent anticyclonic circulation in the open sea during spring-summer and a more or less complex circulation on the continental shelf. Two cells of cyclonic circulation were located over the northern Gulf of Cadiz: the western shelf linked with open-sea processes and the eastern shelf linked to coastal processes (Garcia-Lafuente et al., 2006 and Garcia-Lafuente and Ruiz, 2007).
The northern continental shelf of Gulf of Cadiz is divided by the Cape Santa Maria (off Faro) in two shelves (Fig. 2.1) of different shape that hold different oceanographic
2. Study area
11 processes. The western shelf is narrow, under the influence of the almost-permanent upwelling off Cape São Vicente, related with wind-stress curl (e.g. Fiúza, 1983; Vargas et al., 2003; Criado-Aldeanueva et al., 2006; Garcia-Lafuente and Ruiz, 2007). To the east of Cape Santa Maria, the eastern shelf is wider, receiving sediment and energy inputs from land as well as tidally-driven processes, making this shelf more productive (e.g. Navarro and Ruiz, 2006; Garcia-Lafuente and Ruiz, 2007; Prieto et al., 2009).
The dynamics in this area is also strongly dependent on wind regime. Westerly winds enhance the upwelling off Cape São Vicente, creating a second intense core of upwelling off Cape Santa Maria and generalize the upwelling along the southern Portuguese coast (e.g. Vargas et al., 2003; Garcia-Lafuente and Ruiz, 2007). The upwelling off Cape Santa Maria is a more likely coastal process with a short time response to changes in the wind regime: it is enhanced under westerlies, and it weakens, and even fades out, under easterlies. Off Cape Santa Maria the upwelled water can form a filament extended southward, which reaches its maximum signature at about 40-50 m depth, becoming almost indistinguishable at around 200 m depth (Criado-Aldeanueva et al., 2006). These upwelling events could induce new production resulting from vertical advection of nitrogen, two orders of magnitude greater than the maintained by vertical diffusion (Ruiz and Navarro, 2006). Easterly winds favour a warm coastal countercurrent that is observed in the eastern shelf and may invade the western shelf connecting both shelves in an east to west direction, often turning poleward around Cape São Vicente (e.g. Relvas and Barton, 2002; 2005; Lobo et al., 2004; Garcia-Lafuente et al., 2006; Garcia-Lafuente and Ruiz, 2007; Relvas et al., 2007).
The coastal zone between Guadiana and Guadalquivir River mouths showed the highest chlorophyll concentrations values of northern Gulf of Cadiz, based on remote sensing data from the Sea-viewing Wide Field-of-view Sensor recorded between 1998 and 2000 (Navarro and Ruiz, 2006). In the open sea, the surface chlorophyll maxima occur in winter
and in coastal areas in spring followed by a second bloom either in summer or fall. In coastal areas these blooms are mainly due to the presence of several processes that favour the nutrient entrance, such as upwelling events, rainfall and river discharges. Local winds also influence the biological production, westerlies cause an increase in chlorophyll concentrations and easterlies a decrease in phytoplankton biomass (Navarro and Ruiz, 2006; Prieto et al., 2009).
2.3.2. Tide and wave regime
The coast between the Guadiana and Guadalquivir Rivers is considered mesotidal, with mean tidal range of 2 m. Off the Guadiana River mean spring tide is of 2.82 m (e.g. Morales, 1997) and in the north of the Cadiz Bay can reaches a maximum tidal range of 3.7 m (e.g. Benavente et al., 2000).
The offshore wave climate is dominated by west-southwest waves (71% of the occurrences; Costa et al., 2001). Southeastern waves that consist of short-period waves generated by regional winds are also frequent (23% of the occurrences; Costa et al., 2001). Wave energy is moderate with an average annual significant offshore wave height of 1.0 m and average peak period of 8.2 s (Costa et al., 2001). Storm events in the region were defined as events with significant offshore wave height greater than 3 m (Pessanha and Pires, 1981). Between 1986 and 1993, storm conditions corresponded to 1% of the offshore wave climate (Costa, 1994).
2.4. Shelf bathymetry
On the northern Gulf of Cadiz shelf, the maximum width occur on the middle part, and decreases both towards Cape São Vicente and the Strait of Gibraltar. The shelf has a minimum width of 5 km off Faro (Portugal), increases to 20-25 km off the Guadiana River and attains a width of more than 30 km close to the Guadalquivir River mouth. Accordingly,
2. Study area
13 the mean slope of the Portuguese shelf is 0.5°, decreasing to slopes of less than 0.3° on the Spanish shelf. The mean Guadiana shelf slope is 0.32° (Roque, 1998; Lobo et al., 2001).
The shelf break lies at varying water depths in concordance with width and sea floor gradients changes. Between the mouths of the Guadiana and Guadalquivir Rivers it is located at 120 to 150 m water depth (Baldy, 1977).
2.5. Fluvial and sediment supply
The study area receives fluvial supply from the Guadiana, Piedras, Tinto-Odiel and Guadalquivir Rivers (Fig. 2.1). The annual mean water discharge of Guadalquivir River (160 m3s-1) is about twice the Guadiana River (80 m3s-1) discharge. The contributions of the
Tinto-Odiel system are small, with mean discharges of 20 m3s-1 (van Geen et al., 1997).
The Guadalquivir River is the main fluvial input of the Gulf of Cadiz northern margin, however its associated mud sedimentary wedge is deflected south-eastward of the study area, mainly dominated by the North Atlantic Central Water current (e.g. Gutiérrez-Mas et al., 1996; Nelson et al., 1999; Lobo et al., 2004, as North Atlantic Surface Water). The Guadiana River is, therefore, the main sediment source to the study area. The estimated sediment supply from the river basin to the shelf between 1946 and 1990 was 57.90 x 104 m3yr-1 for the average suspended load and 43.96 x 104 m3yr-1 for bedload (Morales, 1995; 1997). For the winter of 2000/2001, an exceptionally rainy year compared to the previous 10 years, the estimated sand onto the inner shelf was about 7.5-9.5 x 105 m3 (Gonzalez et al., 2007). These
numbers are about twice the average ones estimated by Morales (1995; 1997). However after the closure of the Alqueva dam in February 2002, the sediment supply decreased drastically. Estimates excluding occasional discharges related with flood events, indicated around 10 x 103 m3yr-1 of sand is today exported (as bedload) to the inner shore (Garel et al., submitted).
The second regional sediment source is the littoral drift. Prevailing onshore wave conditions along the coastline produce an eastward net annual littoral drift estimated to be around 180 x 103 m3yr-1 of mostly sandy sediment, carrying sediments from the southern Portuguese coast towards the eastern portion of the Gulf of Cadiz (Gonzalez et al., 2001).
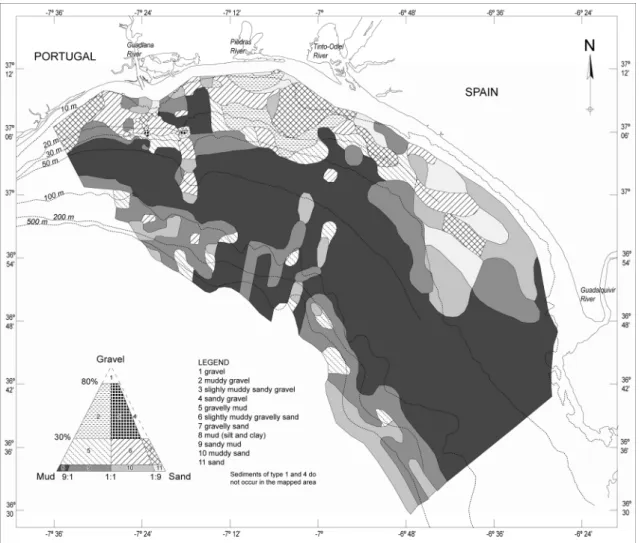
2.6. Modern surficial sediment distribution
Sandy deposits dominate the inner shelf down to a depth of approximately 25 m, particularly in nearshore zones (Fig. 2.2) (Moita, 1985; Fernández-Salas et al., 1999; Nelson et al., 1999; Gonzalez et al., 2004; 2007). This deposits are interrupted by a prodeltaic wedge off the Guadiana Estuary, consisting of sandy muds and muds forming an oblong mud patch with an area of about 60 km2 (Fernández-Salas et al., 1999; Gonzalez et al., 2004; 2007) and by a prodelta off Guadalquivir mouth, consisting of muds (Nelson et al., 1999; Gonzalez et al., 2004; 2007).
The transition between the inner and middle shelf (or outer infralittoral for other authors) located between 25 and 30 m depth consists of sands and sandy mud. Additionally, this area features a series of rocky outcrops of reduced extent, and probably of Holocene age (e.g. Fernández-Salas et al., 1999; Gonzalez et al., 2004), containing varying amounts of terrigenous gravel.
The middle shelf is characterised by an extensive mud belt, consisting of very fine-grained material. Crossing from north to south through the surface of the outer infralittoral ridge and the middle shelf mud deposits lie a series of transgressive deposits composed of muddy gravelly sands and muddy sands (Fig. 2.2) (Gonzalez et al., 2004; 2007).
On the outer shelf below 100 m depth, sediments are generally dominated by sands and silty clay. They are locally interrupted by large patches of sand and gravelly sand in the vicinity of the shelf edge (Gonzalez et al., 2004; 2007).
2. Study area
15
Figure 2.2 – Modern surficial sediment distribution of the northern Gulf of Cadiz continental shelf, between the Guadiana and Guadalquivir Rivers (adapted from Gonzalez et al., 2004).
2.7. Holocene evolution
2.7.1. Continental shelf
The postglacial evolution of depositional systems on the northern Gulf of Cadiz shelf was strongly influenced by rapid sea-level changes, leading to the formation of transgressive and highstand system tracts (Nelson et al., 1999; Lobo et al., 2001; 2002; 2004). Seismic profiles show four transgressive parasequences exposed from the outer to middle shelf (Lobo et al., 2001). Highstand deposits occur on the inner shelf as a prodeltaic wedge and on the middle shelf as an extensive mud belt (Nelson et al., 1999; Gonzalez et al., 2004; 2007; Lobo et al., 2004). The thickest part of the Holocene layer off the Guadiana river mouth is offset to the southeast of the river mouth has an elongate taper to the southeast. All the mud belt
margins except the southeast are interrupted by non depositional areas. At its thickest point is localised in the central shelf, the apparent Guadiana-derived Holocene layer is about 15 m thick and 25 km wide (Nelson et al., 1999).
The sediment distribution patterns in the northern Gulf of Cadiz shelf indicate that, since the Late Quaternary, depositional dynamics express the interaction of several factors dominated by the south-eastward transport of sediments under the influence of North Atlantic Central Water, across the Cadiz shelf toward the Strait of Gibraltar (Nelson et al., 1999). Sediment dispersal during the Holocene highstand period is also controlled by an intermittent counter current system, which seems to be intimately linked with the wind regime (Lobo et al., 2004).
2.7.2. Estuary infilling
Several studies deal with estuarine valley infilling during the Holocene in the northern Gulf of Cadiz. On the Spanish coast these studies have been focused in the Tinto-Odiel, Guadalquivir and Guadalete estuaries (e.g. Dabrio et al., 2000; Lario et al., 2002) and on the Portuguese coast in the Guadiana and Gilão-Almargem estuaries (Boski et al., 2002; 2008; Lobo et al., 2003). More recently Zazo et al. (2008) elaborated an integrated approach from previous papers, including information from both Spanish and Portuguese coastal sectors.
The sequence of events that led to the formation of the Guadiana River estuary in its present form indicated that at the beginning of the Holocene, around 9800 Cal yr BP, the Guadiana estuary started to be rapidly filled with clay sediments. Between 7500-7000 Cal yr BP the central part of the estuary began to accommodate coarser sediments partly derived from the adjacent shelf, whilst clay continued to deposit in the marginal lagoons. Since that time, sea-level rise decelerated, leading to the enclosure of lagoons behind sand spits, and to the deposition of predominantly sandy sediment inside the estuary. This occurred until sea
2. Study area
17 level stabilisation at a level close to the present one which took place around 5000 Cal yr BP (Boski et al., 2002).
2.8. Anthropogenic impact
Palynological evidences in the lower Guadiana valley indicate an important increase in human influence on the landscape after ca. 4000 Cal yr BP (Fletcher et al., 2007). An archaeological approach to regional environmental pollution indicates that in the beginning of Third millennium BC, a sudden territorial occupation in the Iberian Pyrite Belt structured on the cooper metallurgy. This occupation was of such magnitude that provoked an unprecedented impact on the environment, increased deforestation, increase on the rate of erosion processes and contamination with heavy metals, at a regional scale, the waters of the Gulf of Cadiz, specifically the area corresponding to the mouth of Tinto-Odiel River (Nocete et al., 2005).
In the southern Portugal, between ca. 3000 and ca. 300 yr BP, and equating to late pre-Roman, pre-Roman, Moorish and early Portuguese stages of settlement, exploitation of the region for timber and other agricultural products caused widespread erosion, sediment movement, deposition of extensive valley fills and siltation of estuaries (Chester and James, 1999). A more recent survey in southern Portugal in the region of Mértola (a village near the upper Guadiana estuary, approximately 70 km from the mouth), by Boone and Worman (2007), revealed that after cessation of the Roman administration of Iberia in the 5th century A.D. (ca.
1500 Cal yr BP) and over the next 500 years, particularly after the Muslim invasion of A.D. 711 (ca. 1200 Cal yr BP), settlement density increased six fold relative to the number of inhabitants during the Roman period. These authors also verified that during the mid-12th century, the majority of rural villages were abandoned. The region remained largely depopulated until the mid-late 1400s, and settlement density in the region was never again as
high as it was during the later Medieval Islamic period (800 to 900 Cal yr BP). Geoarchaeological evidence of widespread erosion and soil loss suggests that overuse of land may have been a factor in the abandonment (Boone and Worman, 2007).
The anthropogenic influences on the Guadiana River basin have increased significantly during the last 150 years through the construction of dams, mining, and deforestation with consequent increase of soil erosion, which affected the amount of sediment supply to the shelf (Gonzalez et al., 2001; Dias et al., 2004; Gonzalez and Dias, 2006).
Chapter 3
Distribution of living benthic
foraminifera
3.1. Introduction
All applications of benthic foraminifera as environmental indicators are based upon an understanding of their ecology. Any analysis of distribution patterns and abundances of organisms needs to take into account the interaction between individuals, different species, and the chemical and physical environment (e.g. Murray, 1991a; 2006). Only studies of living (stained) fauna can be used for ecological interpretations, to ensure that benthic foraminifera are closely linked to their surrounding environment. Even subtle changes in environmental parameters are mirrored by faunal changes (e.g. Murray, 2001). Because benthic foraminifera are highly dependent on environmental conditions, they have been used extensively for ecological and palaeoecological studies. In the future, foraminiferal ecology will continue to play an important role in the interpretation of the geological record, not only to understand the past but to help predict future trends as well (e.g. Murray, 2006).
Continental shelves are areas of high primary productivity and relatively high environmental variability (Sen Gupta, 1999b). These environments receive nutrients and organic carbon compounds from fluvial supply, as well as from decomposition of organic material in surface sediment and by benthic primary production from seaweed and benthic diatoms. Water movement through wave and tidal circulation distributes nutrients through the water column and promotes enhanced primary production (Murray, 2006). These sources also supply dissolved organic matter, which feeds bacteria and thus supplies the food chain (Loubere and Fariduddin, 1999). Except for species that feed on live food or have endosymbiontes, labile organic matter and the bacteria feeding on it are primary food sources for benthic foraminiferal assemblages (e.g. Murray, 2006).
Dissolved oxygen and organic carbon flux are considered as the main factors controlling depth distribution of benthic foraminifera in sediments (van der Zwaan et al., 1999). In deeper waters, where some environmental factors have little or no spatial or