Angolatitan adamastor
, a new sauropod dinosaur
and the first record from Angola
OCTÁVIO MATEUS1,2, LOUIS L. JACOBS3, ANNE S. SCHULP4, MICHAEL J. POLCYN3, TATIANA S. TAVARES5, ANDRÉ BUTA NETO5, MARIA LUÍSA MORAIS5 and MIGUEL T. ANTUNES6
1CICEGe, Faculdade de Ciências e Tecnologia, FCT, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal 2Museu da Lourinhã, Rua João Luis de Moura, 2530-157 Lourinhã, Portugal
3Huffington Department of Earth Sciences, Southern Methodist University, Dallas, TX, 75275, USA 4Natuurhistorisch Museum Maastricht, de Bosquetplein 6-7, NL6211 KJ Maastricht, The Netherlands
5Geology Department, Universidade Agostinho Neto, Av. 4 Fevereiro, Luanda, Angola 6Academia de Ciências de Lisboa, Rua da Academia das Ciências, 19, 1242-122 Lisboa, Portugal
Manuscript received on October 19, 2009; accepted for publication on June 21, 2010
ABSTRACT
A forelimb of a new sauropod dinosaur (Angolatitan adamastorn. gen. et sp.) from the Late Turonian of Iembe (Bengo Province) represents the first dinosaur discovery in Angola, and is one of the few occurrences of sauropod dinosaurs in sub-Saharan Africa collected with good chronological controls. The marginal marine sediments yielding the specimen are reported to be late Turonian in age and, thus it represents a non-titanosaurian sauropod in sub-Saharan Africa at a time taken to be dominated by titanosaurian forms. Moreover, Angolatitan adamastoris the only basal
Somphospondyli known in the Late Cretaceous which implies in the existence of relict forms in Africa.
Key words:Angola, Cretaceous, Turonian, Dinosaurs, Sauropoda,Angolatitan.
INTRODUCTION
Although very rich in Cretaceous vertebrates, Angola has remained unexplored in recent decades. Beginning in 2005, the field work conducted by the PaleoAngola Project has significantly enhanced the palaeontological record of the Cretaceous of Angola (Jacobs et al. 2006, 2009a, b, Polcyn et al. 2009, Mateus et al. 2008, Schulp et al. 2008). The PaleoAngola Project mounted the first systematic paleontological expeditions that have been conducted since the early 1960s, when Miguel Tel-les Antunes performed an extensive and systematic sur-vey on the Angolan fossil vertebrate fauna the most rel-evant work published in 1964 (Antunes 1964). In May 2005 we (O.M. and L.J.) prospected the coastal area near Iembe which resulted in the discovery, in May, 25th (by
Proceedings of the Third Gondwanan Dinosaur Symposium Correspondence to: Octávio Mateus
E-mail: omateus@fct.unl.pt
OM), of the new taxon reported here. Follow-up exca-vations were performed in May and August 2006. To our knowledge, it was the only systematic prospecting for fossil vertebrates in this area since 1960 by M. Telles Antunes (in September) and M. Mascarenhas Neto.
GEOLOGY ANDASSOCIATEDFAUNA
The term ‘Tadi beds’ was first coined in the 1960 unpub-lished study of G. Brognon , G. Verrier, and R Thiers “Géologie du Bassin du Cuanza et du Bas-Congo” from the “Missão de Pesquisas de Petróleo Petrofina”. It was accepted and used by Antunes (1964) and Antunes and Cappetta (2002). The original study considered the ‘Tadi Beds’ to represent the base of the ‘Senonian’ (= Conia-cian), but the fish fauna led Antunes (1961, 1964: 56) to consider the Tadi Beds to be of late Turonian age. In a revision of the Cretaceous selachians of Angola, Antu-nes and Cappetta (2002: 95) reconfirm the late Turonian age of these beds, and place them in the ‘Itombe Forma-tion’ (see also Jacobs et al. 2006). The Tadi Beds are about 50 m thick (Antunes 1964: 56). G. Brognon et al. (cited in Antunes 1964: 56) provide a list of the inver-tebrate fauna, including the ammonite genera Hemitis-sotia,RomanicerasandPachydiscus, and echinoderms. The fish and shark fauna (Antunes and Cappetta 2002: 95) includesHexanchuscf. microdon,Ptychodus mor-toni,P.cf. paucisulcatus,P. whipplei,Scapanorhynchus raphiodon, S.cf. lewesii,Cretodus semiplicatus, Cre-todus sp., Cretoxyrhina mantelli, Paranomotodon an-gustidens, Squalicorax falcatus, Squalicorax sp., On-chosaurus pharaoh, andEnchoduscf. elegans.
The tetrapod fauna from the Iembe locality (An-tunes 1964, Jacobs et al. 2006; and new unpublished data) includes an eucryptodiran turtle Angolachelys mbaxi(Mateus et al. 2009), the mosasaurs Angolasau-rus bocageiAntunes 1964 (see also Polcyn et al. 2009)
and Tylosaurus iembeensis (Antunes 1964, Lingham-Soliar 1992), Plesiosauria indet., and the sauropod ma-terial described here. The discovery here reported is the first dinosaur and the first Cretaceous terrestrial animal from Angola.
SYSTEMATICS PALEONTOLOGY
Dinosauria Owen, 1842 Sauropoda Marsh, 1878 Neosauropoda Bonaparte, 1986 Titanosauriformes Salgado et al., 1997 Somphospondyli Wilson and Sereno, 1998
Angolatitan adamastorn. gen., n. sp.
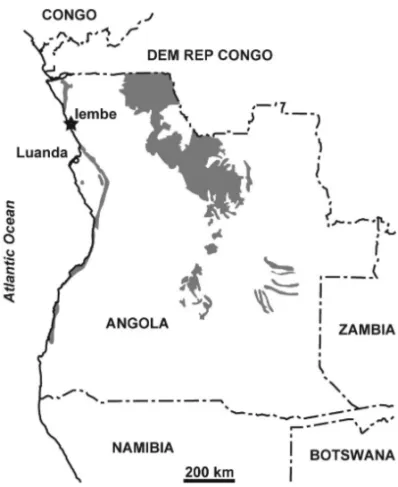
Fig. 1 – Map of Angola, showing the Tadi site (star; 08◦S/13◦E) and
the distribution of Cretaceous strata (grey) in Angola.
ETYMOLOGY
Angolatitan means Angolan giant. Adamastorwas a
mythological sea giant from the South Atlantic feared by the Portuguese sailors.
GENERIC ANDSPECIFICDIAGNOSIS
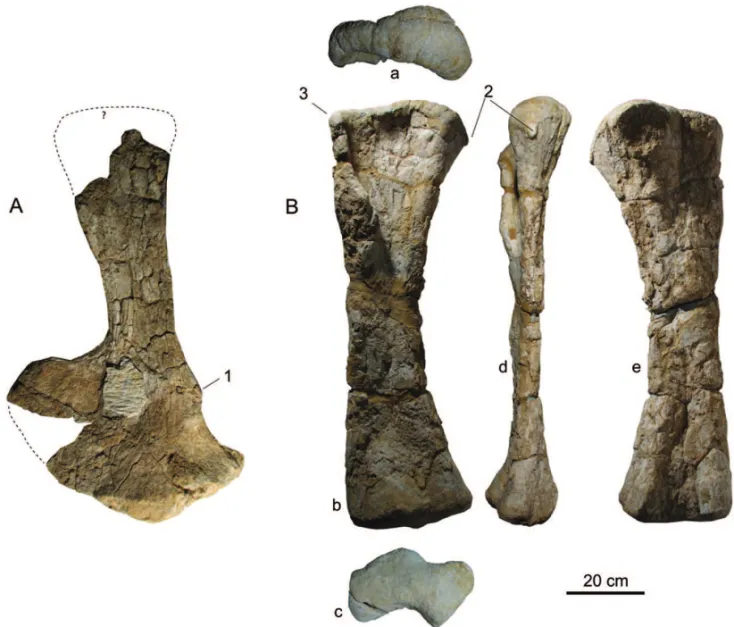
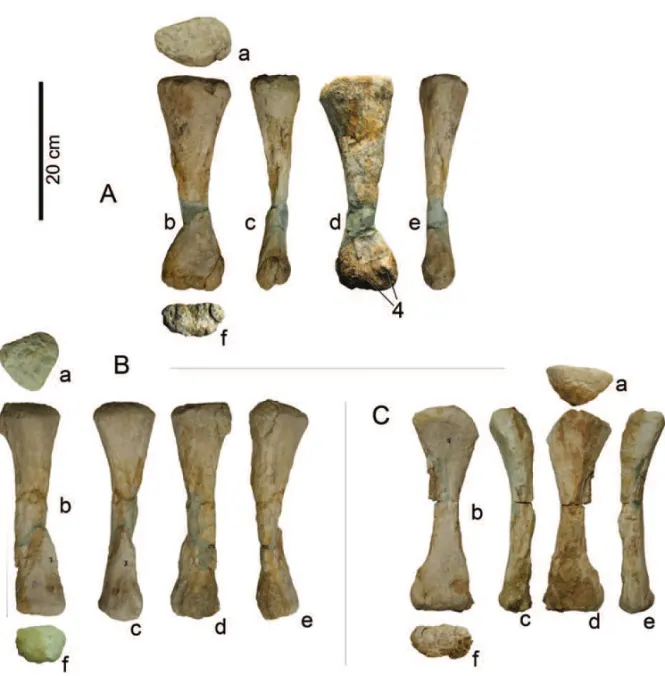
Titanosauriform sauropod with: posteroventral emin-ence in scapula in the proximal one-quarter, anterior to the acromial ridge; acute medioproximal margin of humerus; and rectangular lateral corner of proximal hu-merus; proximal outline of the ulna with anteromedial ridge (process); posterior facet of the distal epiphysis of metacarpal I with two small splint-like projections (Figs. 3 and 5).
TYPELOCALITY ANDHORIZON
Tadi, near Iembe, Tábi community, municipality of Am-briz, Bengo Province, Angola. The precise coordinates of collecting sites are on file at the participating institu-tions.
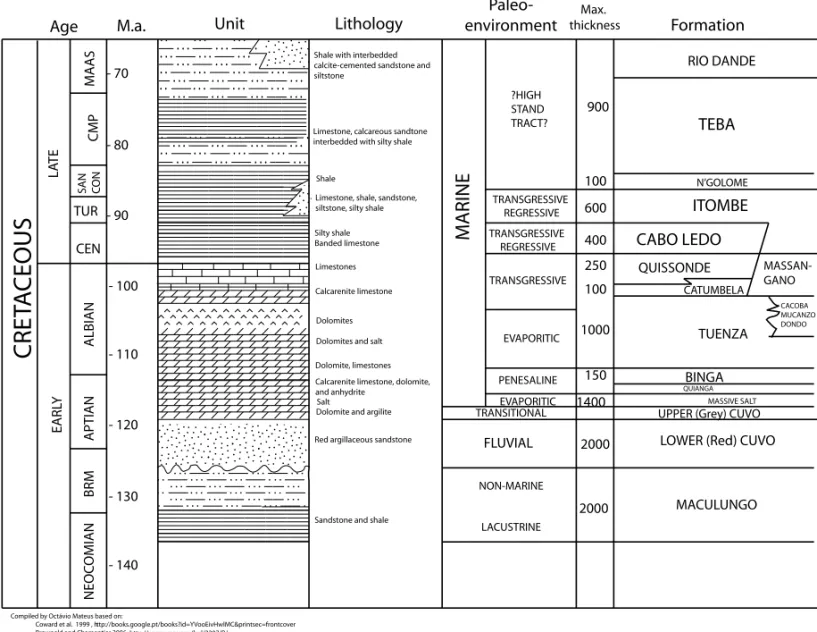
Fig. 2 – Summary of the geology of the Cretaceous of Kwanza Basin (Angola) based on Brownfield and Charpentier (2006) and Coward et al. (1999).
MATERIAL
The material includes a partial right forelimb including the scapula, humerus, ulna, radius and metacarpals I, III, and IV (Table I). All material described here is curated at Museu de Geologia da Universidade Agostinho Neto – PaleoAngola Project, Luanda, Angola (MGUAN-PA), and temporarily registered under field number MGUAN-PA-003 (replicas will be housed at Museu da Lourinhã, Portugal, and at Southern Methodist University, USA).
DESCRIPTION
The scapula is missing its posterior end and part of its proximal expansion. The preserved length is 95 cm, but it is estimated to have been 105 cm if complete. The scapula is overall straight rather than bowed medially.
However, this could be a case of taphonomical distor-tion. The proximal end is incomplete, but does not seem very expanded. The scapular blade is 16.5 cm, with the ventral supraglenoid expansion just above it, and 24 cm on the maximum dorsal expansion. The ventral supra-glenoid expansion is 65 cm in its maximum anteropos-terior width, therefore representing nearly four times the minimum width of the blade. For the description, the orientation of the scapula adopted here is with the scapular blade in a vertical dorsal position as in Romer (1956).
Fig. 3 –Angolatitan adamastorn. gen. et sp., A) right scapula in medial view; B) right humerus in proximal(a), anterior (b), distal (c), medial (d), and posterior (e) views. Autapomorphic features: 1, posteroventral eminence in scapula in the proximal one-quarter, anterior to the acromial ridge; 2, acute medioproximal margin of humerus; and 3, rectangular lateral corner of proximal humerus.
anteriorly, forming a gentle curve in its anterodorsal corner due to the absence of the dorsal hook.
The medial surface is gently concave in all its extension, except in a small (ca. 6 cm) shallow bump placed at the anterior part at the base of the scapular blade, and in the bump in the posteromedial margin of the scapula about 20 cm above the glenoid.
The contact surface between the coracoid and glenoid is perpendicular with the blade in a posterior or anterior perspective, but the contact with the cora-coid runs slightly dorsally, forming a 20◦ angle with
the horizontal (if the scapula is arranged upwards). The glenoid is flat, facing ventrally, but the medial portion, near the midline, bevelled slightly medially, forming an area facing ventromedially. A strongly bevelled
medi-ally glenoid is apomorphic of Somphospondyli accord-ing to Wilson 2002 (character 153).
The ventroposterior margin of the scapula, adja-cent to the glenoid, projects posteriorly and has a small (ca. 9 cm) vertical ridge that borders a groove some-what similar toCetiosaurus oxoniensis. A
posteroven-tral bump in the scapula in the proximal one-quarter is shared/convergent withDystrophaeusandSupersaurus, Ultrasaurus mcintoshi, Lourinhasaurus alenquerensis,
andCamarasaurus supremus, but, unlike other
sauro-pods, it is anterior to the acromial ridge. The glenoid foramen is not visible.
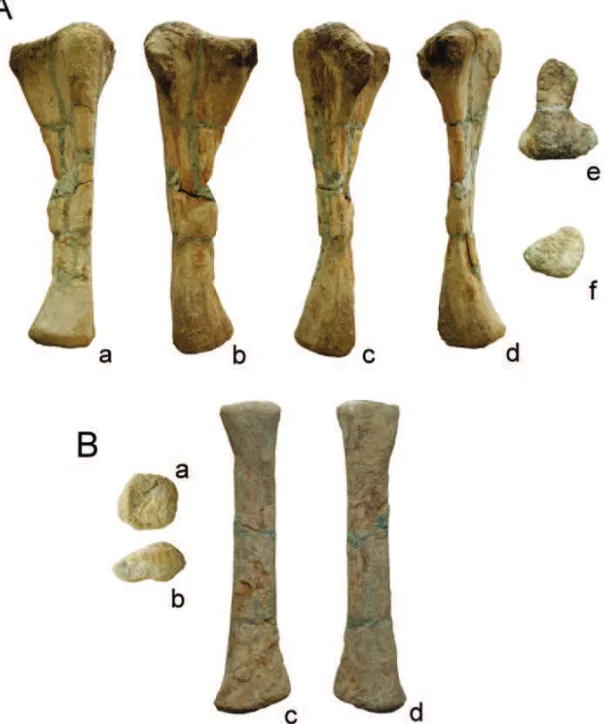
Fig. 4 –Angolatitan adamastorn. gen. et sp., A) right ulna in medial (a), anterior (b), lateral (c), posterior (d), proximal (e) and distal (f) views; and (B) right radius in proximal (a), distal (b), anterior (c), and posterior (d) views.
gracile with a straight shaft. The proximal end is L-shaped due to the well-defined deltopectoral crest. The humeral head is well developed and more projected an-teriorly than the lateral corner, which gives an asymmet-rical outline in anterior view. The lateral corner forms a rectangular corner (rather than a round corner more commonly seen in sauropods) and a straight line to the most proximal part of the humerus. The medioprox-imal corner of the humerus is very pointy due to a 4 cm triangular lateral projection, which is autapomorphic. The triangular deltopectoral crest extends along nearly the upper half of the anterior aspect of the lateral
epiph-Fig. 5 –Angolatitan adamastorn. gen. et sp., right metacarpals I (A), III (B) and IV (C), in proximal (a), anterior (b), medial (c), palmar (d), lateral (e), and distal (f) views. 4, posterior facet of the distal epiphysis of metacarpal I has two small splint-like projections (autapomorphy).
ysis, being well separated by posterior and anterior in-tercondylar grooves. The distal articular facet is slightly convex, nearly planar, without condylar expression. At the anterior face, the distal condyles are directed antero-medially without significant dorsal projection. There is a vertical groove between the lateral condyle and the lat-eral margin, that creates a notch aspect in distal view. In the posterior face, the lateral condyle is round, while the medial condyle forms an acute vertical edge. The poste-rior intercondylar groove is wide and deep.
The right ulna is complete, except for a small part of the lateral face of the diaphysis. The general aspect is gracile. It measures 69 cm in length, 23 cm at its maximum proximal expansion, and 14.5 cm in
width at its distal expansion. In proximal view, the ulna bears a tri-radiate outline, with prominent anterolateral and anteromedial processes, and a less-developed pos-terior process. The proximal outline shows an expanded anteromedial ridge (process) constricted in the base. The anteromedial ridge is nearly twice the lateral one. The olecranon process is poorly developed and does not projects significantly more proximally than the an-teromedial one. The shaft is moderately straight and less robust than in most titanosaurs. The distal epiphysis is sub-round to sub-triangular, with a slight transverse expansion.
circumference expansion expansion
Scapula 95 – 61 –
Humerus 110 43.2 36 29
Ulna 69 27* 23 14.5
Radius 63 22.5 16.5 13
Metacarpal I 30.0 – 10.5 8.5*
Metacarpal III 30.9 13* 9 8.5*
Metacarpal IV 28.5 12 9* –
*Estimated.
epiphyseal expansions. The maximum proximal width equals about 25% of the full length of the radius. The proximal end is sub-round, while the distal end is flat-tened and elongated with an anteroposterior expansion. The medial edge of the distal epiphysis is acute, but the lateral is round. The posterior face of the distal epi-physis bears a groove at the ulnar articulation procud-ing two shallow posterior condyles. The medial part projects more distally forming a bevelled angle with the main shaft.
Three metacarpals (I, III and IV) were present near the distal end of the ulna. With the exception of the bone surfaces exposed to weathering prior to dis-covery, the bones do not show signs of crushing or de-formation. The metacarpal position was identified as I, III, and IV using Janensch (1961), Apesteguía (2005) and Bonnan and Wedel (2004). The metacarpals are elongated, much more than in titanosaurs. All meta-carpals are nearly of the same size, but III is the largest and IV is the shortest preserved element. The distal end bears sutble but separated distal condyles which is a ple-siomorphic condition because the presence of divided condyles is normally associated with the presence of phalanges, lost in titanosaurs (Wilson 2002, Upchurch et al. 2004). The metacarpals bound together by the extensive diaphyseal articulation to form a tubular ar-rangement, U-shaped in proximal view. The length ratio between metacarpal III and the radius is 0.48. A ratio higher than 0.45 is used to be considered a character for Macronaria.
Metacarpal Iis robust, with proximal outline pal-modorsally flattened forming a D-shape outline, and the
medial corner more rounded than the lateral. The meta-carpal presents a crest along the whole lateral facet and with a sinosoidal area at midshaft. The distal end is transversely broad, with two well-defined condyles (but less visible than in non-macronarian sauropods) divided by distal and posterior grooves, the lateral condyle being smaller than the medial. The posterior facet of the distal epiphysis has two small splint-like projections: one just above the intercondylar groove and the other near the lateral condyle, which is autapomorphic.
The rightmetacarpal IIIis complete, with the ex-ception of a portion of the lateral distal condyle. It is subtriangular in proximal view. Ligament attachments on the palmar surface occupy almost the entire proxi-mal half of the metacarpal – which also represents the area of contact with metacarpal IV. The medial facet has an acute crest along the diaphysis that is divided in the proximal third, and rims a large facet facing anterome-dially. The distal end shows defined condyles, being the lateral one the largest.
The metacarpal IV is robust. In the proximal view it is triangular, with the anterior margin being the widest. The proximal surface is flat, but the dorsome-dial corner which is projected medorsome-dially deflects distally forming a 45◦angle with the proximal surface. The
dis-tal end is transversely broad and presents two condyles divided by a medial vertical intercondylar groove, but there is no distal intercondylar groove. This bone re-sembles the metacarpal IV of cf.Laplatasaurusfigured
There are no signs of other carpals, metacarpals or phalanges, but the lateral flattening on metacarpal IV suggests the possible presence of metacarpal V, and the distal condyles suggest the presence of phalanges.
COMPARISONS AND PHYLOGENY
The olecranon process of the ulna is more proximally projected than in Jurassic sauropods Lourinhasaurus,
Lusotitan, Apatosaurus, Camarasaurus, and Diplo-docus, but less than in most Titanosauria, when com-pared with Saltasaurus loricatus “Titanosaurus” col-berti), Tornieria robusta (Janensch 1961), Epachtho-saurus sciuttoi(Martínez et al. 2004), and Malawisau-rus dixeyi(Gomani 2005), and somehow with the same
extension as in Rapetosaurus krausei (Curry Rogers
2009). Ulna robustness (proximal width/length) is 0.33 inAngolatitan adamastor, which is lower than 0.35 in
Lusotitan atalaiensis, 0.35 in aff. TuriasaurusML368,
0.36 inMalawisaurus dixeyi(Gomani 2005: Table 7), 0.34 inRapetosaurus krausei(Curry-Rogers 2009:
Ta-ble 3), and strikingly lower than 0.41-0.43 in Epachtho-saurus sciuttoi (Martínez et al. 2004: Table 2). The
data for establishing the robustness is unreported or un-known in most titanosaurs (e.g. Limaysaurus tessonei,
Rinconsaurus cadamirus,Muyelensaurus pecheni, and Saltasaurus loricatus), for which the ulnae is clearly more robust. Wilhite (2003) provides measures for many Late Jurassic ulnae, which, in average, gives the following means for robustness: 0.40 forApatosaurus,
0.23 for Barosaurus, 0.39 forBrachiosaurus, 0.38 for Camarasaurus, and 0.33 forDiplodocus.
InAngolatitan adamastor, the metacarpals do not
show three titanosaurian characters considered by Apes-teguía (2005): bowed metacarpal I, the flat distal meta-carpal trochlea (although this is difficult to quantify), and reduced intermetacarpal contact that is 1/6 of the metacarpal length.
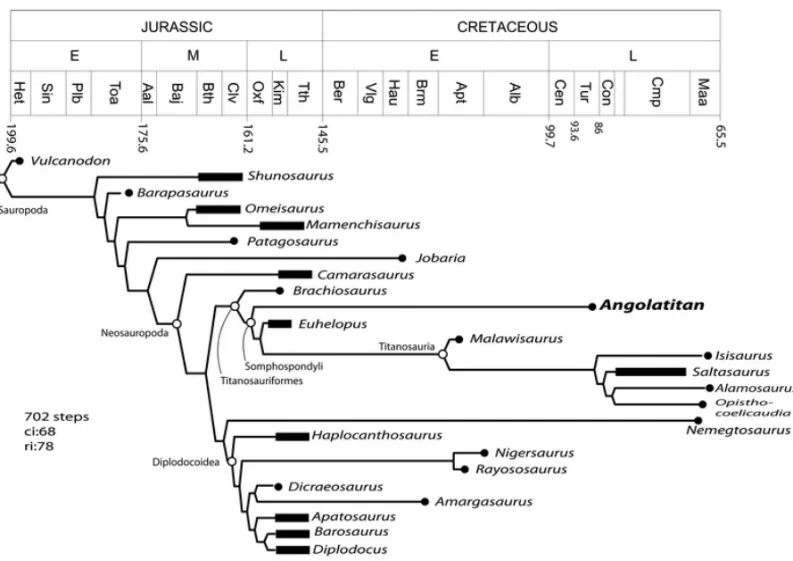
The data matrix was based in Wilson (2002), but the integration of characters in Upchurch et al. (2004) for the forelimb (Angolatitancoding using Wilson 2002, characters 150 to 200: 1 0000????10 0101110010 1???-111100 ?????????? ??????????, and Upchurch et al. 2004, characters 200 to 250: 1 100001???? ??????0101 101000?11? 101?11????). The matrix was processed
using WinClada ver 1.0 (www.cladistics.com) using a heuristic search with 100 replicates. Angolatitan ada-mastorn.gen. et sp. was placed as a basal
titanosauri-form Somphospondyli more derived than Brachiosau-rus, but less derived thanEuhelopus(see Fig. 6). The
metacarpals are slender (close toCamarasaurus
propor-tions and alike titanosaurs asOpisthocoelicaudia, when
compared in the plot provided by Bonnan and Wedel 2004). The robustness (ratio minimum diaphyseal peri-meter/length) is 0,42, which is lower than the range of 0,5-0,6 in most titanosaurs (Apesteguía 2005: Ta-ble 15.2). The size of metacarpals is equivalent (be-ing of variable length in Titanosauridae, see Apesteguía 2005), and the distal end is round, rather than flat as in Titanosauridae.
The Angolatitan adamastoris supported as Som-phospondyli by the following unambiguous synapomor-phies: humeral square proximolateral corner (charac-ter 159 in Wilson 2002), deltopectoral crest medially expanded across the cranial face of the humerus (ch. 220 in Upchurch et al. 2004), and distalmost part of the caudal surface of the humerus deeply concave be-tween the lateral and medial prominent vertical ridges (ch. 221 in Upchurch et al. 2004). However, the scapu-lar glenoid surface faces cranioventrally (ch. 203 in Upchurch et al. 2004) unlikeEuhelopusand other
de-rived Somphospondyli. The Angolan sauropod does not bear titanosaurian synapomorphies such as the ulna with prominent olecranon process (Wilson 2002: character 167) or extremely robust ulna and radius (Upchurch et al. 2004: 309).
GEOGRAPHY ANDCHRONOLOGY
Although sauropod dinosaurs have been reported from elsewhere in Africa, (e.g. Algeria, Cameroon, Egypt, Kenya, Malawi, Mali, Morocco, Niger, South Africa, Sudan, Tunisia, Zambia, Zimbabwe, and Madagascar), no others are known from the Late Turonian. The only described Late Cretaceous sauropods from Africa are the lithostrotian titanosaursAegyptosaurus baharijensis
from Egypt and Niger (Stromer 1932; early Cenoman-ian) andParalititan stromeri from Egypt (Smith et al.
Rapetosau-Fig. 6 – Phylogenetic relationships ofAngolatitan adamastorand other sauropods. See text for explanation.
rus krauseiand“Titanosaurus” madagascariensisfrom
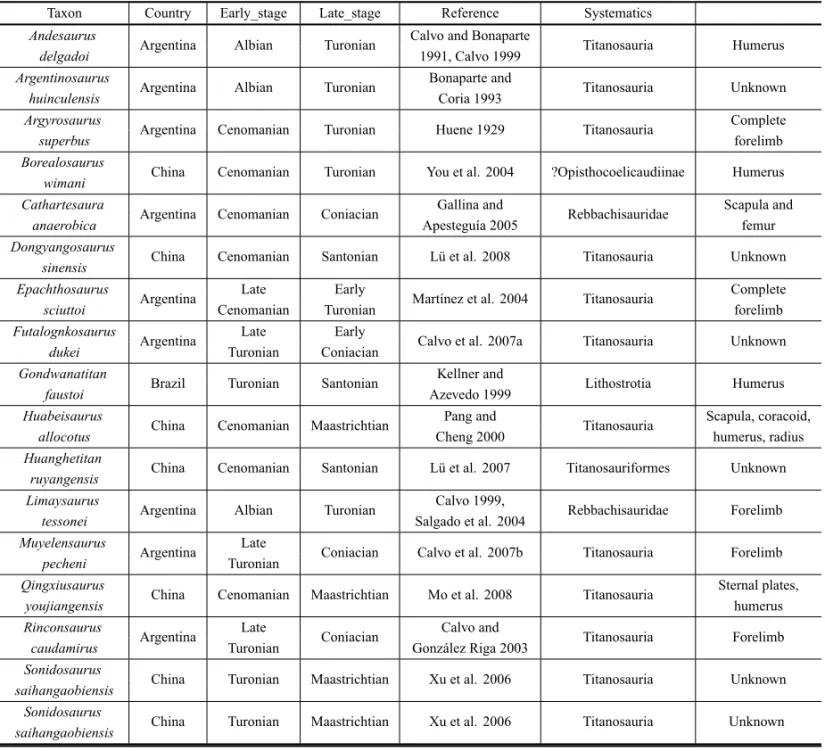
Madagascar (Campanian; Curry Rogers and Forster 2001, Weishampel et al. 2004). Globally, the circa-Turonian sauropods were found only in Argentina, Brazil, and China (Table II), and comprise mostly titanosaurs or diplodocoids. Thus, A. adamastor fills
a 15My gap in the African sauropod record, and an even wider gap in the global record of non-titanosaur sauropods.
Non-titanosaur sauropods, which were dominant during the Jurassic, were being replaced by titanosaurs and derived diplodocoids, becoming rarer in the Late Cretaceous when most identifiable African sauropods were titanosaurs. The reasons for the late Cretaceous dominance of titanosaurs over other sauropods are not fully understood, but it seems to be a general pattern in South America, in Madagascar, and probably in Africa as well.
Although some scarce remains of non-titanosaurs are known, the relevance, distribution, paleogeography,
chronology and even taxonomy of these sauropods is not well understood for the Late Cretaceous of Africa. Diplodocoids were the most successful non-titanosaur sauropods in the Cretaceous, but the presence of pos-sible brachiosaurids (Rauhut and Werner 1997) raises questions regarding the paleogeography of sauropods.
The age of the sediments yielding the specimen is Upper Turonian (∼90Ma) based on the fish and shark teeth (Antunes and Cappetta 2002) and, thus, is one of the best-dated Late Cretaceous sauropod specimens in sub-Saharan Africa. Further work in this area, in part based on ammonites collected with the specimen, will almost certainly provide refined chronology and paleo-geography of the Late Cretaceous dinosaur fauna of Africa.
During the Late Turonian, the coast of Iembe (now at 8◦S latitude) was at about 24◦S latitude due to the
TABLE II
Circa-Turonian sauropod species.
Taxon Country Early_stage Late_stage Reference Systematics
Andesaurus
Argentina Albian Turonian Calvo and Bonaparte Titanosauria Humerus
delgadoi 1991, Calvo 1999
Argentinosaurus
Argentina Albian Turonian Bonaparte and Titanosauria Unknown
huinculensis Coria 1993
Argyrosaurus Argentina Cenomanian Turonian Huene 1929 Titanosauria Complete
superbus forelimb
Borealosaurus
China Cenomanian Turonian You et al. 2004 ?Opisthocoelicaudiinae Humerus
wimani Cathartesaura
Argentina Cenomanian Coniacian Gallina and Rebbachisauridae Scapula and
anaerobica Apesteguía 2005 femur
Dongyangosaurus
China Cenomanian Santonian Lü et al. 2008 Titanosauria Unknown
sinensis Epachthosaurus
Argentina Late Early Martínez et al. 2004 Titanosauria Complete
sciuttoi Cenomanian Turonian forelimb
Futalognkosaurus
Argentina Late Early Calvo et al. 2007a Titanosauria Unknown
dukei Turonian Coniacian
Gondwanatitan
Brazil Turonian Santonian Kellner and Lithostrotia Humerus
faustoi Azevedo 1999
Huabeisaurus
China Cenomanian Maastrichtian Pang and Titanosauria Scapula, coracoid,
allocotus Cheng 2000 humerus, radius
Huanghetitan
China Cenomanian Santonian Lü et al. 2007 Titanosauriformes Unknown
ruyangensis
Limaysaurus Argentina Albian Turonian Calvo 1999, Rebbachisauridae Forelimb
tessonei Salgado et al. 2004
Muyelensaurus
Argentina Late Coniacian Calvo et al. 2007b Titanosauria Forelimb
pecheni Turonian
Qingxiusaurus
China Cenomanian Maastrichtian Mo et al. 2008 Titanosauria Sternal plates,
youjiangensis humerus
Rinconsaurus
Argentina Late Coniacian Calvo and Titanosauria Forelimb
caudamirus Turonian González Riga 2003
Sonidosaurus
China Turonian Maastrichtian Xu et al. 2006 Titanosauria Unknown
saihangaobiensis Sonidosaurus
China Turonian Maastrichtian Xu et al. 2006 Titanosauria Unknown
saihangaobiensis
15◦and 30◦mainly because of the atmospheric air
con-vection Hadley Cells, which has remained stable since the Cretaceous (Jacobs et al. 2009a). Therefore, Iembe was an arid zone during the Turonian, which is further supported by the sediments at Iembe that show no signs of terrestrial productivity. A. adamastor was probably
a sauropod adapted to arid/desert-like environment in a situation similar to the desert elephants today. Rubilar-Rogers (2006) reported a Cretaceous titanosaur sauro-pod from the Atacama Desert (Chile), which is one of the oldest deserts on Earth. However, this material came from lacustrine deposits in Atacama.
ACKNOWLEDGMENTS
who edited some pictures.
RESUMO
Um membro anterior de um novo dinossauro saurópode ( An-golatitan adamastorn. gen. et sp.) do Turoniano Superior de
Iembe (Bengo) representa a descoberta do primeiro dinossau-ro em Angola e é uma das poucas ocorrências de dinossaudinossau-ros saurópodes na África Subsaariana recolhidos com bons con-troles cronológicos. Os sedimentos marinhos marginais de onde provém o espécime estão datados do Turoniano supe-rior e, portanto, trata-se de um saurópodes não-titanossauro na África subsaariana onde predominavam titanossauros. Além disso,Angolatitan adamastoré o único Somphospondyli basal
conhecido no Cretáceo Superior, o que implica a sobrevivência de formas-relíquia na África.
Palavras-chave: Angola, Cretáceo, Turoniano, dinossauros, sauropoda,Angolatitan.
REFERENCES
ANTUNES MT. 1961. Sur la Faune de vertébrés du Cre-tacé de Iembe (Angola). CR Acad Sciences Paris 253: 513–514.
ANTUNESMT. 1964. O Neocretácico e o Cenozóico do lito-ral de Angola. Junta de Investigações do Ultramar, 255 p. ANTUNESMTANDCAPPETTAH. 2002. Sélaciens du Cré-tacé (Albien-Maastrichtien) d’Angola. Palaeontographica 12: 85–146.
APESTEGUÍAS. 2005. Evolution of the titanosaur metacar-pus. Thunder-Lizards, The Sauropodomorph Dinosaur.
Indiana University Press, p. 321–345.
BONAPARTEJFANDCORIARA. 1993. Un nuevo y gigan-tesco sauropodo titanosaurio de la Formacion Rio Limay (Albiano-Cenomaniano) de la Provincia de Neuquén, Argentina. Ameghiniana 30(3): 271–282.
BONNAN MFANDWEDEL MJ. 2004. First occurrence of
Brachiosaurus(Dinosauria: Sauropoda) from the Upper
Jurassic Morrison Formation of Oklahoma. PaleoBios 24(2): 13–21.
BROWNFIELDMEANDCHARPENTIERRR. 2006. Geology and total petroleum systems of the West-Central Coastal Province (7203), West Africa: US Geol Survey Bull 2207-B, 52 p.
Dinosaur Symposium, National Science Museum Mono-graphs #15, In: TOMIDAY, RICHTHANDVICKERS -RICHP (Eds), Tokyo: x + 296 p. P. 13–46.
CALVOJOANDBONAPARTEJF. 1991. Andesaurus delga-doigen. et sp. nov. (Saurischia-Sauropoda), Dinosaurio Titanosauridae de la Formacion Rio Limay (Albiano-Cenomaniano), Neuquén, Argentina. Ameghiniana 28(3-4): 303–310.
CALVOJOANDGONZÁLEZRIGABJ. 2003. Rinconsaurus caudamirus gen. et sp. nov., a new titanosaurid (Dino-sauria, Sauropoda) from the Late Cretaceous of Patago-nia, Argentina. Rev Geol Chile 30(2): 3–23. doi:10.4067/ S0716-02082003000200011. http://www.scielo.cl/ scielo.php?script=sci_arttext&pid=S0716-02082003000200011&lng=en&nrm=iso&tlng=en. CALVOJO, GONZÁLEZ-RIGABJANDPORFIRIJD. 2007b.
A new titanosaur sauropod from the Late Cretaceous of Neuquén, Patagonia, Argentina. Arq Mus Nac 5(4): 485– 504.
CALVOJO, PORFIRIJD, GONZÁLEZ-RIGABJANDKELL -NERAWA. 2007a. A new Cretaceous terrestrial ecosys-tem from Gondwana with the description of a new sauro-pod dinosaur. An Acad Bras Cienc 79: 529–541. COWARDMP, PURDYEG, RIESACANDSMITHDG. 1999.
The distribution of petroleum reserves in basins of the South Atlantic margins. Geol Soc, London, Spec Publ 153(1): 101–131. (Available at http://www.google.com/ books?hl=pt-PT&lr=&id=YVooEivHwlMC).
CURRY ROGERS K. 2009. The postcranial osteology of
Rapetosaurus krausei (Sauropoda: Titanosauria) from
the Late Cretaceous of Madagascar. J Vert Paleont 29: 1046–1086.
CURRYROGERSKANDFORSTERCA. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature 412: 530–534.
GALLINA PA AND APESTEGUÍA S. 2005. Cathartesaura anaerobica gen. et sp. nov., a new rebbachisaurid
(Di-nosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Rio Negro, Argentina. Rev Mus Argentino Cienc Nat 7: 153–166.
GOMANI EM. 2005. Sauropod dinosaurs from the Early Cretaceous of Malawi. Paleontologica Electronica, http:// palaeo-electronica.org/2005_1/gomani27/issue1_05.htm. HUENEF. 1929. Los Saurisquios y Ornitisquios del Cretácico
JACOBS LL, MATEUS O, POLCYN MJ, SCHULP AS, ANTUNESMT, MORAIS ML ANDTAVARES T. 2006. The occurrence and geological setting of Cretaceous di-nosaurs, mosasaurs, plesiosaurs, and turtles from Angola. J Paleo Soc Korea 22: 91–110.
JACOBS LL, MATEUS O, POLCYN MJ, SCHULP AS, SCOTESE CR, GOSWAMI A, FERGUSON KM, ROB -BINS JA, VINEYARDDPANDNETOAB. 2009a. Cre-taceous paleogeography, paleoclimatology, and amniote biogeography of the low and mid-latitude South Atlantic Ocean. B Soc Geol Fr 180(4): 333.
JACOBSLL, POLCYNMJ, MATEUSO, SCHULPASAND NETO AB. 2009b. The Cretaceous Skeleton Coast of Angola. J Vert Paleont 29(Suppl 3): 121A.
JANENSCHW. 1961. Die gliedmaszen und gliedmaszengür-tel der Sauropoden der Tendaguru-schichten. Palaeonto-graphica 3(Suppl 7): 177–235.
KELLNERAWAANDAZEVEDOSAK. 1999. A new sauro-pod dinosaur (Titanosauria) from the Late Cretaceous of Brazil. In: TOMIDAY, RICHTHANDRICHPV, Pro-ceedings of the Second Gondwanan Dinosaur Sympo-sium. Natl Sci Mus Monographs 15: 111–142.
LINGHAM-SOLIAR T. 1992. The tylosaurine mosasaurs (Reptilia, Mosasauridae) from the upper Cretaceous of Europe and Africa. Bull Inst R Sci Nat Belg Sci terre 62: 171–194.
LÜJ, AZUMAY, CHENR, ZHENGWANDJINX. 2008. A new titanosauriform sauropod from the Early Late Creta-ceous of Dongyang, Zhenjiang Province: Acta Geol Sin 82(2): 225–235.
LÜ J, XUL, ZHANGX, HUW, WUY, JIAS ANDJIK. 2007. A new gigantic sauropod dinosaur with the deep-est known body cavity from the Cretaceous of Asia. Acta Geol Sin 81(2): 167–176.
MATEUSO, JACOBSLJ, POLCYNM, SCHULPAS, NETO AB AND ANTUNES MT. 2008. Dinosaur and turtles from the Turonian of Iembe, Angola In: LIVRO DERE -SUMOS DE“TERCER CONGRESOLATINOAMERICANO DEPALEONTOLOGÍA DEVERTEBRADOS” 156 Neuquén, Argentina.
MATEUSO, JACOBSLJ, POLCYNM, SCHULPAS, VINE -YARD D, NETO AB AND ANTUNES MT. 2009. The oldest African eucryptodiran turtle:Angolachelys mbaxi, from the Cretaceous of Angola. Acta Paleo Polonica 54(4): 581–588.
MARTÍNEZ RD, GIMENEZ O, RODRIGUEZ J, LUNA M ANDLAMANNAMC. 2004. An articulated specimen of the basal titanosaurian (Dinosauria: Sauropoda)
Epach-thosaurus sciuttoifrom the early Late Cretaceous Bajo Barreal Formation of Chubut Province, Argentina. J Vert Paleont 24(1): 107–120.
MO JIN-YOU JIN-YOU, MO XIN, HUANGCHUO-LIN, ZHAOZHONG-RU, WANGWEI ANDXU. 2008. A new titanosaur (Dinosauria: Sauropoda) from the Late Cre-taceous of Guangxi, China. Vertebrat PalAsiatic 46(2): 147–156.
PANGQANDCHENGZ. 2000. A new family of sauropod dinosaur from the Upper Cretaceous of Tianzhen, Shanxi Province, China. Acta Geol Sin 74(2): 117–125. POLCYN M, JACOBS LJ, MATEUS O AND SCHULP AS.
2009. New specimens ofAngolasaurus bocageiand com-ments on the early radiations of plioplatecarpine mosa-saurs. J Vert Paleont 29(3): 165A.
RAUHUT OWM AND WERNER C. 1997. First record of Maastrichtian sauropod dinosaur from Egypt. Palaeo Afri-cana 34: 63–67.
ROMERAS. 1956. Osteology of the reptiles. Chicago, Uni-versity of Chicago, 772 p.
RUBILAR-ROGERSD. 2006. Titanosaurs from the Atacama Desert. J Vert Paleont 26(3): 117A.
SALGADO L, GARRIDO A, COCCA SE AND COCCA JR. 2004. Lower Cretaceous rebbachisaurid sauropods from Cerro Aguada Del León, Neuquén Province, northwestern Patagonia, Argentina. J Vert Paleont 24(4): 903–912. SCHULP AS, POLCYNMJ, MATEUS O, JACOBS LJAND
MORAIS ML. 2008. A new species of Prognathodon
(Squamata, Mosasauridae) from the Maastrichtian of Angola, and the affinities of the mosasaur genusLiodon.
Proceedings of the Second Mosasaur Meeting, Fort Hays Studies Special Issue 3, Fort Hays State University, Hays, Kansas, p. 1–12.
SMITH JB, LAMANNA MC, LACOVARA KJ, DODSON P, SMITH JR, POOLE JC, GIEGENGACK R AND AT -TIAY. 2001. A giant sauropod from an Upper Cretaceous Mangrove Deposit in Egypt. Science 292: 1704–1706. STROMERE. 1932. Ergebnisse der Forschungsreisen Prof.
E. Stromer in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharîjestufe (unterstes Cenoman). #11. Sauropoda. Abh. Bayer. Akad. Wissensch. Math.-naturwiss. Abtei-lung 10: 1–21.
UPCHURCHP, BARRETTPMANDDODSONP. 2004. Sauro-poda. p. 259–322 In: WEISHAMPELDB, DODSONP ANDOSMÓLSKAH (Eds), The Dinosauria, 2nd ed. WEISHAMPELDB, BARRETTPM, CORIARA, LELOEUFF
WILHITE R. 2003. Biomechanical reconstruction of the appendicular skeleton in three North American Jurassic sauropods. Ph.D. dissertation. Louisiana State Univer-sity, Baton Rouge, USA.
WILSONJ. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zool J [London] 136(2): 215–275.