Arq. NeuroPsiquiatr. vol.73 número4
Texto
Imagem

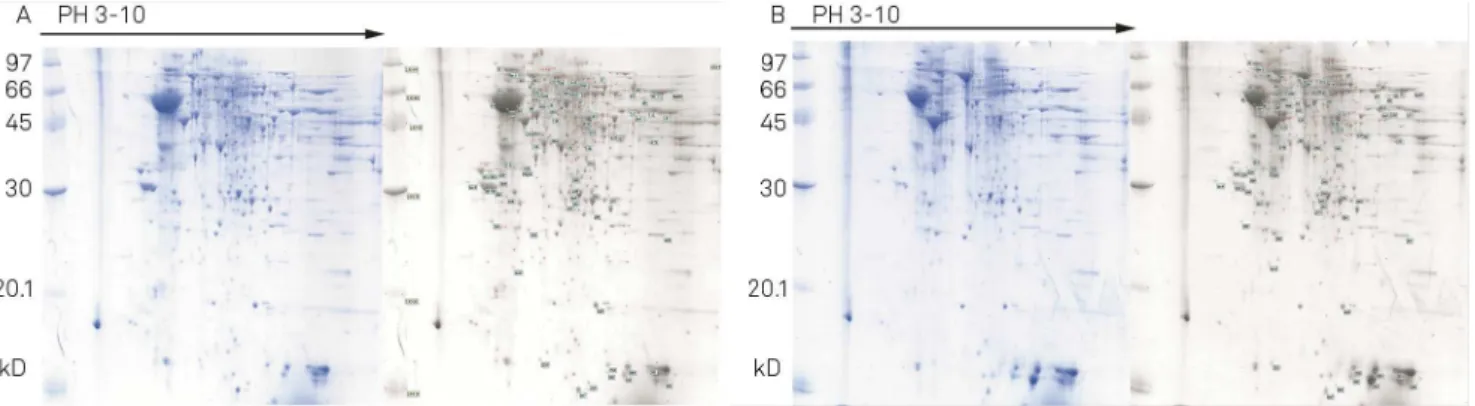
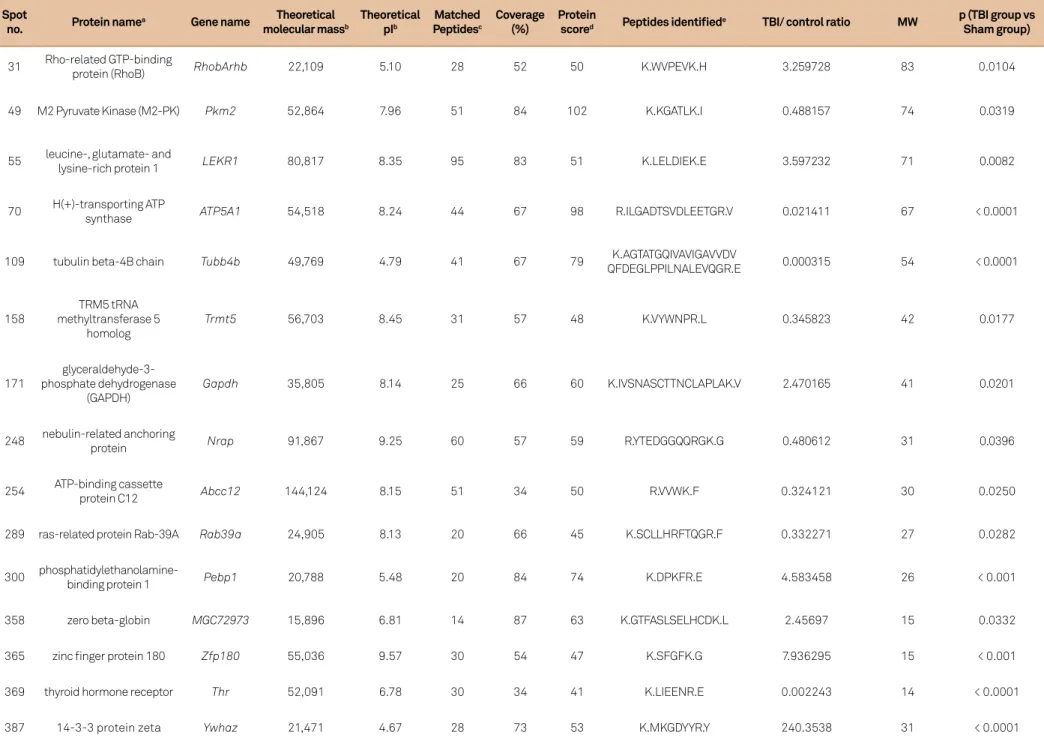
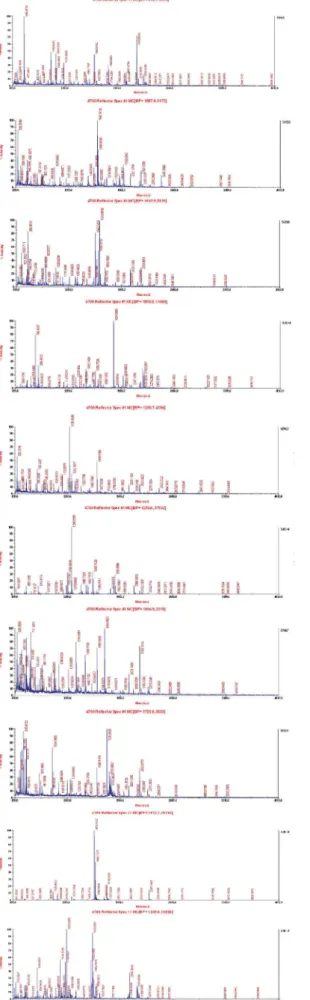
Documentos relacionados
Os resultados da avaliação realizada pelos alunos, através do questionário, sinalizaram para uma boa receptividade relativa a metodologia ativa. As questões propostas, apresentadas
ment applied systemically is effective in pre- venting brain damage in animal models of focal ischemia (14,15), spinal cord ischemia (13), traumatic brain injury (10), and neuro-
We hypothesized that (i) as a group, participants with complicated MTBIs would report greater symptoms and have worse neurocognitive outcomes than those with uncomplicated MTBI,
In Audiology, auditory processing and English language proiciency, the upper semicircular canal dehiscence syndrome and auditory training after traumatic brain injury are
El artículo: “ Measurement of intracranial pressure and short-term outcome of patients with traumatic brain injury: a propensity-matched analysis ” por Biselli-Ferreira et al., (2)
Tal circunstância, além de ser vista como penosa para os próprios AO’s (responsáveis pelo transporte e distribuição deste tipo de produtos pelas várias unidades do
Evidence for cellular damage in normal-appearing white matter correlates with injury severity in patients following traumatic brain injury: A magnetic resonance spectroscopy
Objective documentation of traumatic brain injury subsequent to mild head trauma: multimodal brain imaging with MEG, SPECT, and MRI. Head Trauma
