Artemisinias no tratamento da Malária: Perspectivas futuras
35
0
0
Texto
Imagem



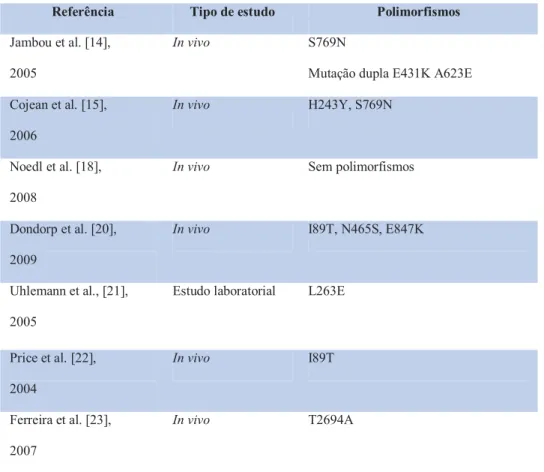
+5
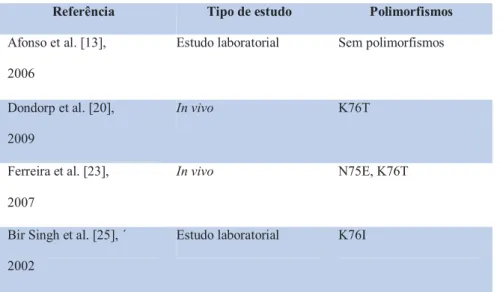
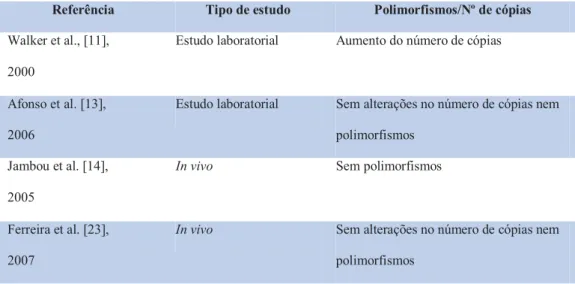

![Figura 3 – Sensibilidade in vitro à dihidroartemisinina no Bangladesh, Tailândia Ocidental, Tailândia Oriental e Cambodja [reproduzido de Noedl et al [19]; incluídos 247 doentes com malária por P](https://thumb-eu.123doks.com/thumbv2/123dok_br/19224625.964323/35.892.262.634.239.634/sensibilidade-dihidroartemisinina-bangladesh-tailândia-ocidental-tailândia-reproduzido-incluídos.webp)
Documentos relacionados