PROGRAMA DE PÓS-GRADUAÇÃO STRICTO SENSU
DOUTORADO EM CIÊNCIAS GENÔMICAS E
BIOTECNOLOGIA
Autor: Ludovico Migliolo
Orientador: Prof. Dr. Octavio Luiz Franco
Co-orientadora: Profª. Drª. Beatriz Simas Magalhães
CARACTERIZAÇÃO MOLECULAR E FUNCIONAL DE
PEPTÍDEOS SINTÉTICOS MULTIFUNCIONAIS DERIVADOS
DE PEPTÍDEOS ANTICONGELANTES DE
Pleuronectes
americanus
LUDOVICO MIGLIOLO
CARACTERIZAÇÃO MOLECULAR E FUNCIONAL DE PEPTÍDEOS SINTÉTICOS MULTIFUNCIONAIS DERIVADOS DE PEPTÍDEOS ANTICONGELANTES DE
Pleuronectes americanus W.
Tese apresentada ao Programa de Pós-Graduação
“Stricto Sensu” em Ciências Genômicas e Biotecnologia da Universidade Católica de Brasília, como requisito para obtenção de Título de Doutor em Ciências Genômicas e Biotecnologia.
Orientador: Prof. Dr. Octávio Luiz Franco
Co-orientadora: Profª. Drª. Beatriz Simas Magalhães
7,5cm
Ficha elaborada pela Biblioteca Pós-Graduação da UCB 10/09/2013. M634c Miglioli, Ludovico.
Caracterização molecular e funcional de peptídeos sintéticos multifuncionais derivados de peptídeos anticongelantes de pleuronecteres americanus W. / Ludovico Miglioli – 2012.
170 f.il.; 30 cm
Tese (doutorado) – Universidade Católica de Brasília, 2012. Orientação: Prof. Dr. Octàvio Luíz Franco
Coorientação: Profa. Dra. Beatriz Simas Magalhães
1. Peptídeos. 2. Infecção. 3. Biotecnologia. I. Franco, Octávio Luiz, orient. II. Magalhães, Beatriz Simas, coorient. III. Título.
Tese de autoria de Ludovico Migliolo, intitulada “CARACTERIZAÇÃO MOLECULAR E
A todas as pessoas que participaram de maneira direta e indireta dessa conquista, mas
principalmente as que foram e são bastante especiais; e que considero muito importante para que
esse trabalho tenha sido o meu maior sonho realizado.
Ao Professor Doutor Octávio Luiz Franco que de maneira direta fez parte da construção
do cientista que sou agora, - por todos os ensinamentos e tempo - dedicados ao meu aprender.
Dedico também às pessoas mais importantes da minha vida que sempre me apoiaram
indiretamente; ao meu eterno amor Renata Bezerra Duarte Migliolo (minha esposa) e a minha
família comandada pelas matriarcas guerreiras Elizabete Luzia Fioravanti, Lúcia Bezerra Varela
AGRADECIMENTO
Eu agradeço sinceramente a todas as pessoas do Centro de Análises Proteômicas e
Bioquímicas - CAPB. Principalmente ao Professor Doutor Octávio Luiz Franco e a Professora
Doutora Beatriz Simas Magalhães que acreditaram nesse trabalho de alguma forma.
Gostaria de agradecer de modo muito especial à pessoa que mais amo nesse mundo
Renata Bezerra Duarte Migliolo a qual me deu muito apoio e principalmente me escutou demais
nessa jornada, que para ela sempre foi uma jornada muito maluca.
Gostaria de agradecer a uma pessoa que mesmo distante, aproximadamente 2572 Km, me
apoiou e apoia em todas as decisões que tomei e tomo, Elizabete Luzia Fioravante (minha mãe),
te amo demais. Também gostaria de agradecer minha pequena irmã, Sophia Migliolo Dias e a
grande irmã, Liss Fioravante Lavra que também amo muito. Mais um agradecimento especial
para a minha sogra que também amo bastante por ter criado tão bem sua filha e por ter aprendido
a fazer ótimas comidas.
Gostaria de agradecer aos meus amigos (as) (irmãos) Anderson Jacome de França,
Rodrigo Aquino de Oliveira, Adeliana Silva de Oliveira e Osmar Silva Nascimento, grandes
A todos os pós-doc e doutorandos que estão e/ou passaram pelo CAPB e EMBRAPA
Aulus, Kelly, Michele (grande parceira), William (grande parceiro), Jeeser (grande parceiro),
Daniel, Suzana, Fabiano, Rocha, Juliane, Bety, Thaís, Symon, Bernardo (grande parceiro),
Rocha, Natália, Aline, Stela, Vivian, Loiane, Clarissa, Leandro, Leonardo Lima Pepino de
Macedo (grande amigo), Carol, Carol (Japão), Professor Doutor Edilson (grande amigo) e
Antonie Honore (good guy).
Aos mestrandos e praticamente mestres Júnior, Mirna, Isabel, Rosa, Renato, Tiago e
Flávia pelas boas horas de risadas e muito entusiasmo nas nossas jornadas de pesquisa.
Aos novos alunos de iniciação cientifica Marlon, Allan, Tatiane, Elaine, Ana Paula,
Hortência e Renata que passaram ou ainda residem no laboratório. A todos os funcionários da
UCB que de algum modo proporcionaram um ambiente adequado para os experimentos, em
especial a nossa técnica “nona” Kênia Chaves. Caso tenha esquecido alguém agradeço a todos
que fazem parte do Centro de Análise Proteômicas.
Agradeço aos amigos australianos professor David Craik, Roben Craik, Michele
Colgrave, Swamy (Indian friend) e Uru (Indian friend) pelos bons momentos culturais,
Agradeço também, aos ilustríssimos professores (as) e pesquisadores (as) Simoni Campos
Dias, Marcelo Bemquerer, Taia Maria, Sonia Maria de Freitas, Diego Nolasco, Cristine Chaves
Barreto, Betânia Ferraz Quirino, Luciano Paulino e Paula Andréia Silva que participaram e
mostraram suas experiências de vida, nos ensinamentos teóricos e práticos, seja respondendo
referes (“mulher mau amada”), dando ideias, opções de experimentos e desta forma me ajudaram
para a publicação de artigos.
Agradeço sempre ao Professor Doutor Maurício Pereira de Sales (in memoriam), que boa
parte da pessoa profissional que sou hoje é produto do que foi ensinado arduamente por ele.
Agradeço a banca examinadora formada pelos Professores Gabriel da Rocha Fernandes,
João Alexandre R. G. Barbosa, Sonia Maria Freitas e Elizeu Antunes dos Santos que se
disponibilizaram de imediato para participar na contribuição com informações e experiências.
Por fim e não menos importante aos patrocinadores, Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior (CAPES) e ao Conselho Nacional de Desenvolvimento Científico e
“Não é o mais forte que sobrevive, nem o mais inteligente, mas o
que melhor se adapta às mudanças”.
RESUMO
MIGLIOLO, Ludovico. Caracterização molecular e funcional de peptídeos sintéticos multifuncionais derivados de Pleuronectes americanus W. 2012. 170 f. Tese (Doutorado em Ciências Genômicas e Biotecnologia), Universidade Católica de Brasília, Brasília, 2012.
Os peptídeos antimicrobianos (PAMs) podem ser considerados parte da defesa inata de peixes e organismos marinhos, sendo a busca destas moléculas de extrema importância no desenvolvimento de terapias no sistema de saúde pública. O trabalho, portanto, objetiva o desenho racional de peptídeos com características multifuncionais baseado em peptídeos anticongelantes isolado da espécie Pleuronectes americanus. Dez peptídeos ricos em alaninas foram sintetizados, em que quatro deles foram separados em uma primeira geração e desenhados com base direta no peptídeo anticongelante HPLC-8 e após avaliações funcionais da primeira geração um redesenho foi proposto e seis novos peptídeos foram sintetizados (segunda geração) de acordo com modificações de carga variando entre +3 a +6 e momento hidrofóbico com ângulos menores que 100°. Dentre os resultados encontrados para os peptídeos da primeira geração destacou o peptídeo Pa-MAP apresentando atividade contra a espécie Escherichia coli. A atividade antiviral e antifúngica também foi observada para Pa-MAP contra o vírus de herpes humano 1 e 2 e contra os fungos Trichophyton mentagrophytes, T. rubrum e Candida parapsilosis. Por fim, a atividade antitumoral frente à células tumorais coloretais Caco-2 foram também observadas para Pa-MAP. Já na segunda geração de peptídeos podemos destacar Pa -MAP 1.5 e Pa-MAP 1.9 os quais demonstraram MIC de 3,2 e 6 μM, respectivamente contra E. coli. Todos os peptídeos não apresentaram atividade hemolítica e citotóxica frente à células Vero (células fibroblásticas do rim do Cercopitheccus aethiops) e RAW 264.7 (células derivados de monócitos de camundongo), nas concentrações máximas usadas nos ensaios biológicos. Os peptídeos Pa-MAP 1.4 e Pa-MAP 2.0 causaram modificações na geometria do cristal de gelo demonstrando afinidade por uma face do cristal. A conformação do Pa-MAP foi medida em vários pHs (3-11), temperaturas (25-95 °C) e força iônica por dicroísmo circular demonstrando que Pa-MAP foi estável sob diferentes condições. A determinação estrutural teórica para todos os peptídeos foram realizados por meio de modelagem molecular em que se observou a formação da
potenciais no desenvolvimento de ferramentas biotecnológicas contra infecções causadas por micro-organismos patogênicos.
ABSTRACT
Antimicrobial peptides (AMP) are considered essential part of marine organisms and fish innate defense being these peptides important compounds for development of public health system therapies. This work, therefore focus on a rational peptides design with multifunctional properties based in antifreeze peptides previously isolated from Pleuronectes americanus. Ten rich-alanine peptides were synthesized, being four of them classified from first generation with physical-chemical parameters directly linked to HPLC-8 antifreeze peptide. After functional validations, six more peptides were redesigned of physical-chemical improvement parameters including charges around +3 to +6 and hydrophobic moment lower than 100°. First peptides generations were purified by RP-HPLC with a semi-preparative C18 column and its purity were evaluated by mass spectrometry MALDI-ToF. Data showed that Pa-MAP first generation peptide presented antibacterial activity against Escherichia coli. Antiviral activity was also demonstrated by Pa -MAP against human herpes virus 1 and 2 antifungal activity and for Trichophyton mentagrophytes, T. rubrum and C. parapsilosis fungi. In addition, antitumoral activity was observed forward Caco-2 colorectal cells in culture for Pa-MAP. Furthermore, were also observed that second generation peptides Pa-MAP 1.5 and Pa-MAP 1.9 showed deleterious activity toward E. coli. All peptides did not showed hemolytic and cytotoxicity activities against Vero cells isolated from Cercopitheccus aethiops monkey kidney fibroblast and RAW 264.7 cells derived from mouse monocytes which were tested using maximum concentrations used in bioassays in vitro. Interesting results were obtained for Pa-MAP 1.4 and Pa-MAP 2.0 peptides which showed affinity by ice crystal face in antifreeze assay in vitro performed causing ice crystal geometry modifications. Biophysical experiments showed that Pa-MAP structural conformation measured into various pHs (3-11), temperatures (25-95 °C) and ionic strength by circular dicrhoism in vitro was structurally stable. Furthermore, theoretical tridimensional structures were also performed for all designed and redesigned peptides by using molecular
modeling. α-Helical conformations were observed and validated for all peptides. In summary, Pa-MAP and its analogues of first and second generation showed capability to interact on several biological membranes and showed to be potentials candidates in development of biotechnological tools against infections caused by pathogenic microorganisms.
Lista de Ilustrações
Figura 1. Estruturas tridimensionais dos representantes dos AMPs que apresentam conformação em
hélices anfipáticas. ...31
Figura 2. Estrutural tridimensional dos PAMs ricos em cisteínas...33
Figura 3. Estrutura tridimensional dos peptídeos estabilizado por duas ligações dissulfeto na conformação de duas fitas beta antiparalela. ...35
Figura 4. Estrutura química e tridimensional dos peptídeos ricos em aminoácidos regulares ...37
Figura 5. Estrutura tridimensional dos PAMs com ênfase na conformação mista com fitas betas antiparalelas estabilizadas por uma ligação dissulfeto ...39
Figura 6. Estrutura tridimensional do peptídeo composta por uma α-hélice. ...45
Figura 7. Estrutura tridimensional do peptídeo apresentando conformação estrutural composta por α -hélice. ...46
Figura 8. Eventos que ocorrem na membrana citoplasmática de bactérias na sequência de ação dos peptídeos antimicrobianos em geral. ...48
Figura 9. Visão geral do amplo espectro de ação dos peptídeos de defesa. ...49
Figura 10. Figura ilustrativa da espécie de peixe ósseo com os dois olhos situados do mesmo lado. ...55
Figura 11. Estrutura tridimensional do AFP tipo I resolvido por difração de raio-X...56
Figura 12. Alinhamento entre Pa-MAP e o peptídeo anticongelante HPLC-6 (pdb 1j5b) isolado de P. americanus. ...80
Figura 13. Perfil de purificação do peptídeo sintético Pa-MAP ...82
Figura 14. Curva de titulação de Pa-MAP com hidróxido de sódio ...88
Figura 15. Mudanças conformacionais de Pa-MAP avaliada na região Far-UV por meio de dicroismo circular (CD) nos ambientes hidrofílicos: água e hidrofóbicos: TFE e SDS...91
Figura 16. Estrutura tridimensional do peptídeo alvo em estudo modelado. ...93
Figura 17.Avaliação da dinâmica molecular de Pa-MAP ...96
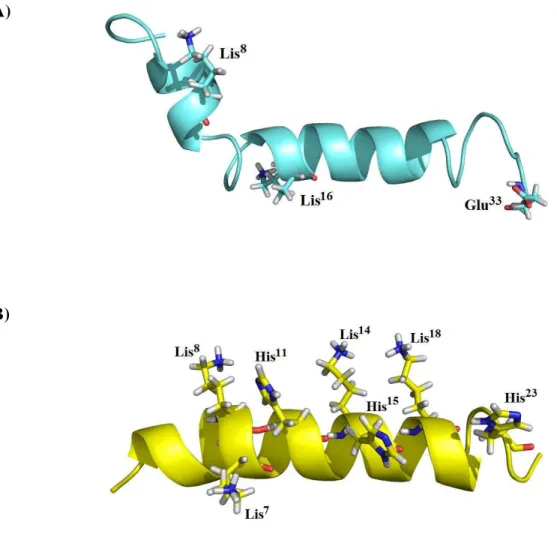
Figura 19. Diagramas de hélices utilizados para comparação do (A) peptídeo anticongelante HPLC-8 demonstrando de cada lado os ângulos de (1) 140° e (2) 220° para os resíduos hidrofílicos e hidrofóbicos respectivamente. Os desenhos racionais dos peptídeos análogos (B) Pa-MAP 1.2 (C) Pa-MAP 1.3 e (D) Pa-MAP 1.4 destacando com a linha pontilhada os ângulos que definem os peptídeos como uma hélice
anfipática. ...100
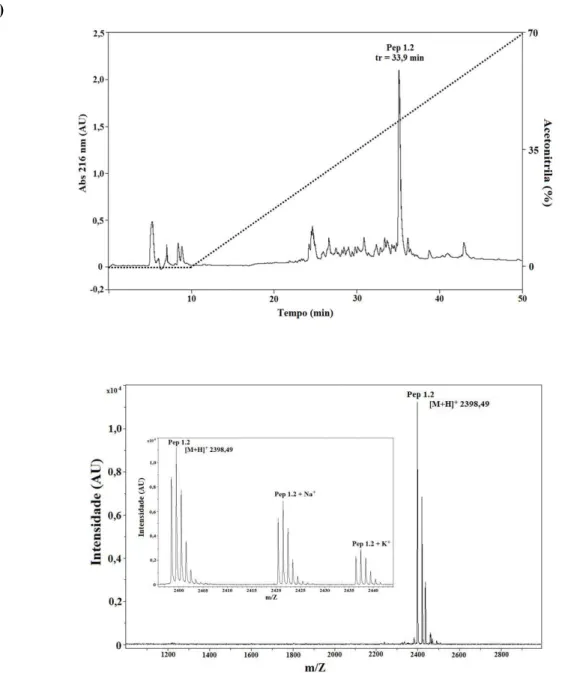
Figura 20. Perfil de purificação do peptídeo sintético Pa-MAP 1.2 por meio de cromatografia em fase reversa, coluna C18 NST, 5 µm, 250 mm x 10 mm semipreparativa. (B) Análise de Pa-MAP 1.2 por espectrometria de massa; massa monoisotópica [M+H+++] = 2398,49. O quadrado interno representa os adutos de Na+ e K+. ...102
Figura 21. Perfil de purificação do peptídeo sintético Pa-MAP 1.3 (A) por meio de cromatografia em fase reversa, coluna C18 NST, 5 µm, 250 mm x 10 mm semipreparativa. (B) Análise de Pa-MAP 1.3 por espectrometria de massa; massa monoisotópica [M+H+] = 2323,68. O quadrado interno representa os adutos de Na+ e K+. ...103
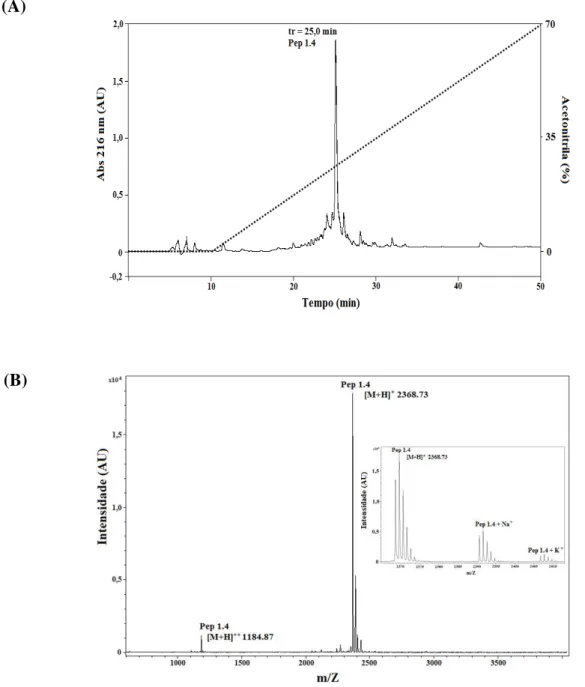
Figura 22. Perfil de purificação do peptídeo sintético Pa-MAP 1.4 (A) por meio de cromatografia em fase reversa, coluna C18 NST, 5 µm, 250 mm x 10 mm semipreparativa. (B) Análise de Pa-MAP 1.4 por espectrometria de massa; massa monoisotópica [M+H+] = 2368,73. O quadrado interno representa os adutos de Na+ e K+. ...104
Figura 23. Diagramas de hélice utilizado para construção dos peptídeos redesenhados com os momentos hidrofóbicos organizados (A) Pa-MAP 1.5, (B) Pa-MAP 1.6, (C) Pa-MAP 1.7, (D) Pa-MAP 1.8, (E) Pa -MAP 1.9 e (F) Pa-MAP 2.0. ...106
Figura 24. Alinhamento, validação e modelo final para Pa-MAP 1.2. ...114
Figura 25. Alinhamento, validação e modelo final para Pa-MAP 1.3. ...115
Figura 26. Alinhamento, validação e modelo final para Pa-MAP 1.4. ...116
Figura 27. Alinhamento, validação e modelo final para Pa-MAP 1.5. ...117
Figura 28. Alinhamento, validação e modelo final para Pa-MAP 1.6. ...118
Figura 29. Alinhamento, validação e modelo final para Pa-MAP 1.7. ...119
Figura 30. Alinhamento, validação e modelo final para Pa-MAP 1.8. ...120
Figura 31. Alinhamento, validação e modelo final para Pa-MAP 1.9. ...121
Lista de Quadros
Quadro 1. Principais características das respostas imunológicas adquiridas Bernstein, 1998. ...26
Quadro 2. Alguns exemplos de peptídeos antimicrobianos isolados de peixes marinho. ...41
Quadro 3. Peptídeos antimicrobianos com outras atividades. ...52
Lista de Tabelas
Tabela 1. Toxicidade e concentração mínima inibitória (MIC) de Pa-MAP contra células de mamíferos, células tumorais, fúngicas (levedura e micélios), bactéria Gram-positiva e -negativa e vírus. ...85
Tabela 2. Avaliação da capacidade de Pa-MAP em inibir o crescimento do cristal de gelo, sendo avaliado o valor de histerese termal. ...87
Tabela 3 Conteúdo de estrutura secundária de Pa-MAP usando o método descrito por Morrisett e colaboradores 1973. As varreduras no comprimento de onda de 222 nm foram realizadas a uma temperatura de 25 °C na concentração de 150 μM de Pa-MAP. ...89
Tabela 4. Toxicidade e concentração mínima inibitória (MIC) dos peptídeos desenhados e redesenhados da primeira e da segunda geração contra células de mamíferos, células tumorais, bactérias Gram-positivas e -negativas. ...110
Tabela 5. Avaliação da capacidade dos peptídeos de primeira (Pa-MAP 1.4) e segunda (Pa-MAP 2.0) geração em inibir o crescimento do cristal de gelo, sendo determinado o valor de histerese termal (HT). Todos as amostras foram solubilizadas com tampão bicarbonato 100 mM, pH 8,1 para o ensaio. ...111
Tabela 6. Resumo dos valores obtidos para as regiões do Mapa de Ramachandran e os valores de g-score para todos os peptídeos de primeira e segunda geração. ...113
Lista de Abreviaturas
AFGP - Glicoproteína anticongelante
AFP - Peptídeo anticongelante
ATCC - The American type culture collection ASA - Área de acessibilidade ao solvente
Blastp - Basic Local Alignment Search Tool protein
CLSI - Clinical and Laboratory Standards Institute
CD - Dicroismo circular CO2 - Dióxido de carbono
DMF - N,N-dimetilformamida
EDT - 1,2-etaneditiol
ED50 - Dose efetiva capaz de inibir 50 % da propagação viral
HPLC - Cromatografia líquida de alta performance
MALDI-ToF - matrix-assisted laser desorption/ionization time of flight mass spectrometry MIC - Concentração inibitória mínima
MAP - Peptídeo com múltiplas atividades
MNTC - Concentração máxima não-tóxica
NCLS - National Committee for Clinical Laboratory Standards
Pa-MAP - Pleuronectes americanus - peptídeo com múltiplas atividades PAM - Peptídeos antimicrobianos
PBS - Tampão fosfato salino
PDB - Banco de Dados de Proteínas (Protein Data Bank)
RPMI-1640 - Meio Roswell Park Memorial Institute-1640
SDS - Dodecil sulfato de sódio SD - Desvio padrão da média
TFA - Ácido trifluoracético
TFE - 2,2,2-trifluoroetanol TIS - triisopropilsilano
SUMÁRIO
1INTRODUÇÃO ... 22
1.1 OCEANO, UMA FONTE DE MOLÉCULAS BIOATIVAS COM POTENCIAL BIOTECNOLÓGICO ... 22
1.2 RESPOSTA IMUNE GERAL ... 23
1.3 SISTEMA IMUNOLÓGICO DOS PEIXES ... 26
1.4 PEPTÍDEOS ANTIMICROBIANOS (PAMs) EM GERAL ... 27
1.5 CLASSIFICAÇÃO DOS PAMS ... 298
1.5.1 PAMs helicoidais ... 298
1.5.2 AMPs ricos em cisteína ... 31
1.5.3 PAMs contendo fitas-β ... 32
1.5.4 PAMs ricos em aminoácidos repetidos ... 35
1.5.5 PAMs contendo aminoácidos raros... 38
1.6 PEPTÍDEOS ANTIMICROBIANOS ISOLADOS DE PEIXES ... 38
1.7 MECANISMO DE AÇÃO PARA PEPTÍDEOS ANTIMICROBIANOS ... 46
1.8 PEPTÍDEOS MULTIFUNCIONAIS ... 49
1.9 PLEURONECTES AMERICANUS ... 53
2 JUSTIFICATIVA ... 58
3 HIPOTESE ... 58
4 OBJETIVO GERAL ...49
5 METODOLOGIA ...60
5.1 ANÁLISE IN SILICO DE SEQUÊNCIAS DE PROTEÍNAS ANTICONGELANTES ... 60
5.2 SÍNTESE DA PRIMEIRA EM SEGUNDA GERAÇÕES DE PEPTÍDEOS ... 61
5.3 PURIFICAÇÃO E DETERMINAÇÃO DA PUREZA DOS PEPTÍDEOS SINTÉTICOS DA PRIMEIRA GERAÇÃO ... 63
5.4 QUANTIFICAÇÃO DOS PEPTÍDEOS ... 64
5.5 TITULAÇÃO POTENCIOMÉTRICA ... 65
5.6 ANÁLISES FUNCIONAIS IN VITRO ... 65
5.6.1 Avaliação da atividade hemolítica... 65
5.6.2 Ensaio de citotoxicidade ... 66
5.6.3 Ensaios para células cancerosas em cultura... 67
5.6.4 Ensaio antifúngico ... 67
5.6.5 Ensaios antibacterianos ... 68
5.6.6 Células e vírus ... 69
5.6.7 Avaliação da atividade antiviral ... 70
5.6.8 Avaliação da atividade anticongelante ... 72
5.7 ESTUDOS CONFORMACIONAIS IN VITRO E IN SILICO DE Pa-MAP ... 73
5.7.1 Dicroísmo circular ... 73
5.7.2 Modelagem molecular ... 75
5.7.3 Dinâmica molecular ... 76
6 RESULTADOS ... 78
6.2 ENSAIOS IN VITRO PARA Pa-MAP ... 82
6.2.1 Testes antibacterianos ... 82
6.2.2 Testes antivirais ... 82
6.2.3 Testes antifúngicos ... 83
6.2.4 Estudos citotóxicos para células tumorais ... 83
6.2.5 Estudos de citotoxidade ... 84
6.2.6 Atividade anticongelante ... 85
6.3 ANÁLISES ESTRUTURAIS DE Pa-MAP IN VITRO ... 86
6.3.1 Titulação potenciométrica ... 86
6.3.2 Dicroísmo circular ... 87
6.4 ANÁLISES ESTRUTURAIS DE Pa-MAP IN SILICO ... 91
6.4.1 Modelagem molecular ... 91
6.4.2 Dinâmica molecular (DM) ... 93
7 RESULTADOS ... 97
7.1 PEPTÍDEOS DESENHADOS E SINTETIZADOS COM BASE EM MOTIVOS ANTICONGELANTES E SEUS REDESENHOS ... 98
7.1.1 Peptídeos sintetizados na primeira e segunda geração ... 98
7.2 AVALIAÇÃO DA ATIVIDADE ANTIBACTERIANA ... 107
7.3 ESTUDOS CITOTÓXICOS PARA CÉLULAS TUMORAIS ... 108
7.4 ESTUDOS DE CITOTOXIDADE ... 108
7.6 ANÁLISE IN SILICO DA ESTRUTURA TRIDIMENSIONAL DOS PEPTÍDEOS DA
PRIMEIRA E SEGUNDA GERAÇÃO ... 111
8 DISCUSSÃO ... 125
9 CONCLUSÃO ... 138
REFERÊNCIAS ... 139
23
1
INTRODUÇÃO
1.1 OCEANO, UMA FONTE DE MOLÉCULAS BIOATIVAS COM POTENCIAL
BIOTECNOLÓGICO
Com pesquisas sendo desenvolvidas em diversas áreas, o ambiente marinho é considerado
fonte de moléculas biologicamente ativas com enorme potencial biotecnológico (Rajanbabu e
Chen, 2011b). Além disso, com as técnicas atualmente usadas na purificação destas moléculas
bioativas abre-se um horizonte enorme no desenvolvimento de compostos a serem usados na luta
contra patógenos humanos. O uso de metodologias de desenho e redesenho racional de peptídeos,
por exemplo, podem vir a ser uma excelente saída na resolução de diversos problemas na saúde,
como são os casos das infecções hospitalares e transplantes de órgãos que nas últimas décadas
apresentam resultados promissores. Para o desenvolvimento destes produtos, o ambiente marinho
aparece como um excelente alvo de estudo com ênfase na exploração das estratégias de defesa
em geral dos peixes (Rajanbabu e Chen, 2011b). Os peixes apresentam uma série de estratégias
desenvolvidas para a defesa que vai desde a defesa contra micro-organismos patogênicos até
mecanismos de sobrevivência em ambientes aquáticos onde as temperaturas se mantêm abaixo de
zero (climas polares, por exemplo) (Amir et al., 2005; Rajanbabu e Chen, 2011b)
Outro ponto de vista importante para o desenvolvimento de produtos biotecnológicos
consiste na busca por moléculas bioativas com múltiplas funções. Estas moléculas bioativas são
24
24
alternativa aos antibióticos utilizados atualmente em infecções adquiridas em serviços de saúde
(IASS). Diante desta visão, o ambiente marinho apresenta um cenário valioso para a exploração
de moléculas bioativas encontradas no sistema imunológico de peixes.
1.2 RESPOSTA IMUNE GERAL
A palavra inflamação vem do latim; “in + flamma + ação” que significa “em + fogacho
(calor, rubor) + atividade”, do grego phlogos: que significa “chama”. Fisiologicamente, consiste
em uma resposta do corpo frente a uma lesão, invasão por fatores externos e ocasionalmente, a
uma auto-agressão (Trowbridge e Emling, 1996). Os processos inflamatórios aparecem após a
ação dos mais diferentes agentes agressores físicos, químicos ou biológicos e consiste em um
fenômeno dinâmico, razão pela qual seu aspecto morfológico se modifica com o tempo (Pereira e
Bogliolo, 1994).
A inflamação se caracteriza por três sinais básicos, descritos por Celsius, no primeiro
século D.C.: calor, rubor, tumor, além da sintomatologia dolorosa. Só mais tarde, no século
passado, deu-se importância às alterações funcionais que quase sempre acompanham uma reação
inflamatória. A maioria das inflamações agudas e crônicas se desenvolve e se amplificam como
resultado do recrutamento de componentes humorais e celulares (Gallin, 1993).
A reação à substâncias estranhas, inclusive macromoléculas e polissacarídeos, é o que se
denomina imunidade. Historicamente, imunidade significa proteção contra as doenças e, mais
25
constituem o sistema imune, e sua resposta coordenada e coletiva à entrada de antígenos
estranhos consiste na resposta imune. Porém, mecanismos que normalmente protegem os
indivíduos de infecções e eliminam substâncias estranhas são capazes de causar lesão tecidual e
doenças em algumas situações (Abbas, Lichtman e Pober, 2000).
A defesa contra micro-organismos pode ser mediada, inicialmente, pela imunidade inata,
seguida pela imunidade adquirida. A imunidade adaptativa, chamada imunidade específica em
função de sua capacidade de reconhecer diferentes micro-organismos e macromoléculas, tem
como componentes os linfócitos e seus produtos. A resposta adaptativa utiliza muitos
mecanismos efetores da imunidade inata para eliminar micro-organismos. As características que
definem essa resposta são: a especificidade, a diversidade, a memória, expansão clonal; a
especialização, a autolimitação (contração e homeostasia) e a tolerância (Quadro 1) (Abbas,
Lichtman e Pober, 2000).
A imunidade inata por outro lado, também chamada de natural ou nativa, provê a primeira
linha de defesa contra micro-organismos. Ela consiste em uma resposta rápida, imediata à entrada
de um dado antígeno (corpo estranho), e ocorre de forma semelhante para diferentes agentes
infecciosos. Dentre os seus principais componentes podemos destacar as barreiras físicas e
químicas; as células fagocíticas tais como neutrófilos, macrófagos e as células NK, “natural
killer”; as proteínas e peptídeos sanguíneos, incluindo o sistema complemento e outros
mediadores da inflamação como as citocinas que regulam e coordenam muitas atividades das
26
26 Quadro 1. Principais características das respostas imunológicas adquiridas Bernstein, 1998.
Característica Significado Funcional
Especificidade Garante que antígenos distintos desencadeiem respostas especificas Diversidade Capacita o sistema imunológico a responder a uma grande variedade de antígenos
Memória Conduz a respostas intensificadas durante as exposições repetidas ao mesmo antígeno
Expansão Clonal Aumenta o número de linfócitos antígeno-específicos para que se mantenham atualizados com os micróbios
Especialização Gera respostas que são ideias para a defesa contra diferentes tipos de micróbios Contração e
homeostasia Permite ao sistema imunológico responder a novos antígenos encontrados Tolerância a
antígenos próprios Evita lesão do hospedeiro durante respostas a antígenos estranhos
1.3 SISTEMA IMUNOLÓGICO DOS PEIXES
Em geral os peixes participam de níveis tróficos maiores ou menores com as cadeias
alimentares dos ecossistemas aquáticos e abrigam um grande número de parasitas. Além disso, os
peixes não representam apenas os mais antigos organismos, mas também a maior classe de
vertebrados (Ravichandran et al., 2010b). O sistema imune destes animais mostra similaridade ao
dos vertebrados superiores, com algumas diferenças importantes. Os peixes possuem sistema
imunológico adaptativo (memória imunológica) menos desenvolvido que o mecanismo
imunológico dos vertebrados superiores. Entretanto desde o início dos anos 90, as pesquisas
sobre a fisiologia, filogenia e ontogenia do sistema imune de peixes vêm aumentando
consideravelmente. Contudo, esse conhecimento ainda é escasso se comparado ao dos
27
Os peixes possuem características únicas em sua anatomia e seu metabolismo devido ao
ambiente onde vivem. O contato permanente com o ambiente aquático e a intensa interelação
com organismos saprófitos e micróbios patogênicos capazes de digerir a matéria orgânica em
decomposição cria um ambiente onde os peixes tem que desenvolver respostas imunológicas
rápidas e eficientes (Plouffe et al., 2005). Esta resposta imune de defesa aparentemente é
essencial para a sobrevivência desta classe e inclui muitos elementos, tais como peptídeos de
ativação do complemento, citocinas, fagossomos e peptídeos antimicrobianos (Cole, Weis e
Diamond, 1997; Ellis, 2001; Fernandes e Smith, 2002; Magnadottir, 2006).
A resposta imune dos peixes também pode ser dividida em dois tipos: resposta inata, que
consiste em impedir que os agentes patogênicos tenham acesso ao organismo hospedeiro,
podendo eliminar os patógenos e bloquear sua entrada; a resposta imune adaptativa, caracterizada
pela especificidade e memória imunológica, induzida por substâncias denominadas de
imunógenos (Bernstein, Schluter e Marchalonis, 1998). Segundo Bly e Clem (1994), o sistema
imunológico inato em peixes possui grande versatilidade, desempenhando um papel importante
na resposta imune, visto que o sistema adaptativo responde lentamente quando comparado ao de
mamíferos, principalmente em temperaturas da água abaixo da comumente encontrada a espécie
(Bly e Clem, 1992).
Igualmente a outras espécies, os peixes são suscetíveis à doenças causadas por parasitas,
vírus, bactérias e fungos, sendo a sobrevivência ou a morte destes, determinadas pela eficácia do
sistema imune em prevenir a infecção inicial e/ou prevenir o crescimento do agente patogênico
após a infecção iniciada (Balfry e Higgs, 2001). Segundo Iwana e Nakanishi (1996), fatores
28
28
proteção externa e interna contra agentes infecciosos. Apesar da distinção na classificação desses
dois sistemas de defesa, deve-se entender que sempre que um agente patogênico ataca o
organismo, este se defende mediante a interação da maioria dos elementos que compõem o
sistema imune, em que vários fatores de cada sistema podem agir separadamente ou em
combinação (Fernandez, De Blas e Ruiz, 2002).
Em resumo, durante a sua evolução, os peixes adquiriram um enorme número de
respostas imune inatas para se defender contra invasores patogênicos, entretanto as moléculas que
apresentam maior potencial biotecnológico são as pequenas moléculas compostas por
aminoácidos denominadas de peptídeos os quais apresentam um vasto espectro de atividade.
1.4 PEPTÍDEOS ANTIMICROBIANOS (PAMs) EM GERAL
Atualmente, mais de 1950 PAMs diferentes foram identificadas em vários organismos
como insetos, plantas e animais, incluindo os seres humanos (Menousek et al., 2012). Além
destes organismos, também foram observados a produção destas moléculas de defesa em
bactérias Gram-positivas e especialmente nos organismos produtores de ácido lático (Luders et
al., 2003). A maioria destes peptídeos antimicrobianos, naturalmente pode ser sintetizado como
um prepropetídeo constituído por uma sequência sinal denominada de peptídeo sinal localizado
na região N-terminal (a qual auxilia no direcionamento para o retículo endoplasmático), um
segmento de “pro” e um segmento posterior na região C-terminal catiônico que demonstra
29
2010a). Estes peptídeos podem ser agrupados com base na sua estrutura primária, composição de
ácidos, tamanho e estrutura tridimensional.
1.5 CLASSIFICAÇÃO DOS PAMS
A análise da estrutura tridimensional destes peptídeos vem apresentando informações que
melhoram o conhecimento da função destas pequenas moléculas bioativas. Com base nas
estruturas tridimensionais obtidas por ressonância magnética nuclear (RMN), juntamente com a
análise das sequências primárias destes PAMs. Estes peptídeos podem ser classificados em cinco
grupos: denominados de PAMs helicoidais, PAMs ricos em cisteínas, PAMs fitas-β, PAMS ricos
em aminoácidos regulares (repetição de um mesmo aminoácido na composição) e PAMs
compostos por aminoácidos raros ou incomuns.
1.5.1 PAMs helicoidais
Polipeptídeos antibacterianos pertencentes a um grupo de ocorrência natural,
compartilhando semelhanças anfipáticas como estrutura em α-hélice, que podem interagir
fortemente e permear membranas fosfolipídicas. Como exemplos, podemos citar a α-hemolisina
descoberta em Staphylococcus. aureus; melitina um potente antibacteriano encontrado na abelha
do mel, paradaxina peptídeo citotóxico isolado do peixe Pardachirus marmoratus (Primor e Tu,
30
30
No início da década de 80 foi identificado e caracterizado um PAM da hemolinfa de
Hyalophora cecropia e assim surgiu a família das cecropinas (Steiner, 1982) (Figura 1A). Desde
então grande parte do trabalho estrutural e bioquímico tem sido focado sobre esses PAMs que
apresentam este tipo de conformação estrutural. Em geral as cecropinas apresentam a
conformação em hélice diante de certos solventes orgânicos como o trifluoroetanol (TFE)
(Cammers-Goodwin et al., 1996). Os estudos iniciais com cecropina-A utilizando RMN
demonstraram um padrão helicoidal quando testado em uma solução com 15 % de álcool
hexafluoroisopropil (Holak et al., 1988). Os resultados permitem inferir a presença de uma hélice
anfipática altamente hidrofóbica e catiônica com superfícies carregadas e um motivo protéico
observado em muitos outros PAMs. Magaininas são peptídeos bem caracterizados compostos de
23 resíduos isolados da pele do sapo Xenopus laevis, uma espécie de anfíbio Africano, estudos de
RMN mostraram que cecropinas como, magaininas também formam estruturas helicoidais
anfipáticas em ambiente hidrofóbicos como em solução de TFE a 25% (Figura 1B) (Marion,
Zasloff e Bax, 1988; Matzusaki, 1999). A foulicidina um PAM catiônico isolado de Gallus
domesticus (galinha) apresenta potente atividade antimicrobiana in vitro e estrutura
tridimensional em α-hélice (Xiao et al., 2006) (Figura 1C). Mastoparam, outro catiônico PAM
isolado do veneno de vespas, Vespula lewisii, também apresenta estrutura tridimensional em α
-hélice em ambientes hidrofóbicos, como detergentes e membranas (Todokoro et al., 2006)
31
Figura 1. Estruturas tridimensionais dos representantes dos AMPs que apresentam conformação em hélices anfipáticas. (A) cecropina (pdb 2igr), (B) magainina
(pdb 2mag), (C) foulicidina (pdb 2amn) e (D) mastoparam (pdb 2czp). Em azul estão representados os resíduos hidrofílicos de lisina e arginina, em amarelo os resíduos hidrofóbicos de isoleucina, leucina, fenilalanina, triptofano, valina e em verde estão representados os resíduos ácido glutâmico, alanina, asparagina, glicina, histidina, metionina, prolina, serina, tirosina e treonina. A visualização foi realizada utilizando o programa PyMol v0.99.
KWKVFKKIEKKWKVFKKIEKAGPKWKVFKKIEK
GIGKFLHSAKKFGKAFVGEIMNS
RVKRVWPLVIRTVIAGYNLYRAIKKK
INWKGIAAMAKKLL (A)
(B)
(C)
32
32 1.5.2 PAMs ricos em cisteína
Os peptídeos de neutrófilos humanos HNP-1, -2 e -3 foram os primeiros peptídeos ricos
em cisteína isolados a partir de granulócitos humanos e foram denominados de defensinas (Ganz
et al., 1985). Estas defensinas consistem em uma família de peptídeos que apresenta em geral
cerca 30 aminoácidos e uma grande quantidade de resíduos de cisteína, estando presente em uma
grande variedade de organismos (Figura 2A). A maioria destas moléculas de defensinas
apresenta um motivo conservado de seis resíduos de cisteína que formam três ligações dissulfeto.
As posições das ligações dissulfeto são em sua maioria entre as Cis1ª-Cis4ª, Cis2ª-Cis5ª e Cis3ª
-Cis6ª. Estudos de cristalografia de raios-X com a HNP-3, em combinação com a centrifugação por
sedimentação, sugerem que o peptídeo possa formar um dímero estável (Ravichandran et al.,
2010a). A estrutura resolvida pela metodologia de RMN destas defensinas mostra a presença de
três fitas beta antiparalelas. A drosomicina, isolada a partir da drosofila (pequena mosca) contém
quatro ligações dissulfeto e são constituídas por três fitas-β antiparalelas com uma α-hélice entre
os dois primeiros eixos estruturais que se assemelham e encaixam em um mesmo grupo de PAMs
33 Figura 2. Estrutural tridimensional dos PAMs ricos em cisteínas (A) HNP-1 (pdb 3hjd) mostrando as 3 ligações dissulfeto características e (B) drosomicina (pdb 1myn) que apresenta quatro ligações dissulfeto, três fitas-β
antiparalelas (amarelo) e uma α-hélice. A visualização foi realizada utilizando o programa PyMol v0.99.
1.5.3 PAMs contendo fitas-β
Alguns dos PAMs conhecidos podem ser estáveis no formato estrutural de um gancho ou
anzol com cerca de 20 resíduos de comprimento, apresentando uma ou duas ligações dissulfeto
com duas fitas-β antiparalelas (Ravichandran et al., 2010a). Os peptídeos taquiplesinas e
polifemusina II isolados de extratos ácidos de hemócitos do Tachypleus tridentatus compartilham
um motivo fita-β harpina estabilizado por duas ligações dissulfeto (Figura 3A). Estudos de
RMN, juntamente com as estruturas tridimensionais indicam que taquiplesina mostra forte
semelhança com protegrinas, peptídeos isolados de leucócitos porcino (Tamamura et al., 1993).
Ambas as moléculas formam fita-β antiparalela ligada por meio de duas ligações dissulfeto
(Figura 3B). Estudos de RNM com o peptídeo tanatina isolado do inseto hemípteros Podisus
maculiventris também demonstram resultados semelhantes aos da taquiplesina, apresentando um
par de fitas-β antiparalela mantido por uma única ligação dissulfeto (Ravichandran et al., 2010a)
34
34
(Figura 3C). Lactoferricina B, um fragmento de 25 aminoácidos obtidos por meio da digestão
proteolítica de lactoferrina um proteína isolada do leite de mamíferos, foi sintetizado
quimicamente tornando-se uma nova molécula análoga devido às novas características
físico-químicas adotadas, as quais regem sua estabilidade conformacional, por exemplo, este fragmento
não sofre mais influência de resíduos adjacentes e pode apresentar outra estruturação em solução.
Portanto, este peptídeo sintético adota em solução uma estrutura de fita-β antiparalela
estabilizado por uma única ligação dissulfeto como mostrado por estudos de RMN (Figura 3D)
35 Figura 3. Estrutura tridimensional dos peptídeos (A) taquiplesina (pdb 1wo1) (B) protegrina 1 (pdb 1zy6), (C) tanatina (pdb 8tfv) e (D) lactoferrina (pdb 1lfc), demonstrando o motivo harpina estabilizado por duas ligações dissulfeto na conformação de duas fitas beta antiparalela. A visualização foi realizada utilizando o programa PyMol v0.99.
(A)
(B)
(C)
36
36 1.5.4 PAMs ricos em aminoácidos repetidos
Alguns PAMs são compostos de um elevado número de aminoácidos iguais. As
conformações estruturais destes peptídeos são em algumas vezes diferentes das normalmente
encontradas como α-hélice e/ou fita-β. Histatina, por exemplo, consiste em um peptídeo isolado
da saliva humana que apresenta abundância de resíduos de aminoácido como a histidina sendo
ativo contra o fungo Candida albicans (Xu et al., 2005) (Figura 4A). Outra interessante família
de peptídeos antimicrobianos são as catelicidinas, peptídeo rico em aminoácidos regulares,
encontrado em lisossomos de macrófagos e leucócitos de mamíferos (Figura 4B). PR-39 é outro
PAM rico em prolina e arginina, isolado do intestino de porcos e demonstra eficiência no
combate contra bactérias (Agerberth et al., 1991). Outros exemplos com aminoácidos
regularmente presentes em sua composição são as tritripticina e as indolicidinas, antimicrobianos
ricos em triptofano isolados de mamíferos, responsáveis pela primeira linha de defesa a invasores
37 Figura 4. Estrutura química e tridimensional dos peptídeos ricos em aminoácidos regulares (A) histatina, rico em histidina, (B) tritripticina (pdb 1d6x) e (C) indolicidinas (pdb 1qxq) ricos em triptofano. A visualização foi realizada utilizando o programa PyMol v0.99.
(B) (C) (D)
38
38 1.5.5 PAMs contendo aminoácidos raros
Alguns peptídeos são compostos de aminoácido modificados e são considerados raros ou
incomuns (Ravichandran et al., 2010a). O melhor exemplo, por ser o melhor estudado, é o
peptídeo produzido pela espécie Lactococcus lactis, antimicrobiano denominado nisina-A
composto de anéis de lantionina, 3-metillantionina, desidroalanina e deidrobutirina, ácido
N-acetilmurâmico, farnesil pirofosfato entre outros (Figura 5A) (De Vos et al., 1993). Este
peptídeo demonstrou atividade deletéria contra bactéria Gram-positiva e não apresenta qualquer
conformação tridimensional definida em água, por outro lado revela estruturação tridimensional
em geral α-hélice quando ligado à micelas de dodecil fosfocolina (DPC). Outro interessante PAM
consiste na leucocina-A, composta por 37 aminoácidos e isolada da espécie Leuconostoc gelidum
uma bactéria produtora de ácido lácteo (Figura 5B) (Fregeau Gallagher et al., 1997). Este
peptídeo apresenta uma conformação anfifílica bem estudada por interagir com membrana.
Nestes peptídeos antimicrobianos podemos observar modificações pós-traducionais que não
resultam em conformações vistas nas outras classes de PAMs descritas acima. Entretanto são
39 Figura 5. Estrutura tridimensional dos PAMs (A) nisina-A isolada de Lactococcus lactis, com ênfase nos resíduos de aminoácidos incomuns como z-dehidrobutireno (DBU) e 2,3-didehidroalanina (DHA), modificação da treonina e da serina respectivamente. (B) Estrutura tridimensional da leucocina (pdb 2leu) apresentando conformação. O peptídeo apresenta conformação mista com fitas betas antiparalelas estabilizadas por uma ligação dissulfeto e uma cauda em
α-hélice na presença de TFE. A visualização foi realizada utilizando o programa PyMol v0.99.
1.6 PEPTÍDEOS ANTIMICROBIANOS ISOLADOS DE PEIXES
PAM isolados de peixe tanto terrestres como marinhos vem recebendo nas últimas
décadas uma atenção especial, principalmente pela atividade demonstrada por estas pequenas
moléculas. O baixo índice de infecção encontrado em peixe é notável e isso tem estimulado cada
vez mais estudos do sistema de defesa inato destes animais (Ravichandran et al., 2010a). Alguns
peptídeos antimicrobianos têm sido identificados em peixes e são responsáveis por este baixo
índice de infecção, no entanto, o número de PAMs isolados e identificados a partir das células da
epiderme ou secreção da pele, guelras e intestino de peixes ósseos (teleósteos) vem nas últimas
40
40
com sequências de proteínas conhecidas, entretanto com outra função, o que sugere uma
derivação de produtos de clivagem de proteínas maiores, tais como histonas e proteínas
ribossomais reforçando ainda mais a ideia de multifuncionalidade (Robinette et al., 1998;
Birkemo et al., 2003; Fernandes et al., 2004). Outro ponto de vista importante e ao mesmo tempo
desafiador consiste em que os PAMs em peixes foram ignorados como potencial fonte de
peptídeos antimicrobianos. Este fato reflete na baixa quantidade de peptídeos depositados no
Antimicrobial Database (APD), o qual apresenta aproximadamente 1950 PAMs depositados a
partir de diferentes fontes, entretanto relativamente uma pequena parcela, cerca de 10 %, tem sido
41 Quadro 2. Alguns exemplos de peptídeos antimicrobianos isolados de peixes marinho.
Peptídeo Peixe Ambiente
Massa Molecular (kDa) Número de aminoácidos
Fonte Referência
Pleurocidina Pleuronectes americanus NOA /
Antártico 2,7 25
pele e
intestino (Mason et al., 2006)
LEAP2 Pleuronectes americanus NOA /
Antártico 2,1 - 2,7 19 – 24
pele e
intestino (Liu et al., 2010)
MGCATH37 Myxine glutinosa NOA 3,5 - 4,6 30 – 37 intestio (Mason et al., 2006)
Pardaxina Pardachirus marmoratus Índico 3,3 33 muco da
pele (Vad et al., 2010)
Piscidina-1 Morone saxatilis NOA 2,5 22 pele, guelra (Moon et al., 2007)
Misgurina Misgurnus anguillicaudatus Pacífico 2,5 21 peixe inteiro (Park et al., 1997)
Hepcidina Oncorhynchus mykiss Pacífico 2,3 21 fígado (Douglas et al., 2003)
Hipposina Hippoglossus hippoglossus NOA 5,4 51 muco da
pele (Birkemo et al., 2003)
Mixinidina Myxine glutinosa NOA 1,3 12 muco da
pele
(Subramanian, Ross e Mackinnon, 2009)
LCRP Petromyzon marinus NOA /
42 NOA: Norte do Oceano Atlântico; Antártico: Oceano Antártico; Índico: Oceano Índico; Pacífico: Oceano Pacífico.
Índico
Parasina-I Parasilurus asotus NOA 2,0 19 pele (Cho et al., 2002)
Epinecidina-1 Epinephelus coioides Índico /
Pacífico 3,0 25 pele (Pan et al., 2007)
β-defensina Oncorhynchus mykiss NOA/
43
As diversas funções dos PAMs de peixes já foram bem relatadas, quase todos estes já
foram isolados e caracterizados demonstrando atividade antibacteriana ou bacteriostática diante
de várias linhagens de bactérias Gram-negativas e -positivas. A epinecidina-1 e seus análogos
sintéticos demonstraram medidas de ruptura da membrana observada contra bactérias
Gram-negativas (Pan et al., 2009). Já em outro estudo utilizando hepcidinas isoladas de Japonese
flounder (JF), nome científico da espécie Paralichthys olivaceus, ficou demonstrado que o
peptídeo JF1, uma isoforma, não exibiu função antimicrobiana. Por outro lado, o peptídeo JF2
com 26 aminoácidos apresentou atividades contra as bactérias Gram-negativas, Escherichia coli e
Gram-positivas como S. aureus e Lactococcus garvieae (Hirono et al., 2005). Transgênicos da
espécie Danio rerio conhecido popularmente como “zebrafish”, expressando hepcidinas de
tilápia (HT)2-3 mostraram resistência contra a infecção causada pela bactéria Gram-negativa,
Vibrio vulnificus (Hsieh, Pan e Chen, 2010). Notavelmente, inúmeras cópias codificando
múltiplos genes de hepcidina em “Japonese flounder” foram observadas em tilápia, salmão do
Atlântico e várias outras espécies (Douglas et al., 2003; Huang, Chen e Kuo, 2007). Entre HTs, o
gene HT 2-3 expresso foi induzido pela endotoxina bacteriana lipopolissacarídica (LPS), mas
duas outras variantes deste gene, HT 1 e HT 2-5-2, não foram fortemente induzidas (Huang,
Chen e Kuo, 2007). É possível, portanto, concluir que as múltiplas cópias de hepcidina evoluíram
em espécies diferentes de peixe com diferentes funções biológicas. Um estudo realizado com
epinecidina-1, HT 1-5, e HT 2-3 demonstrou que os análogos sintéticos destes peptídeos
desestruturava a membrana externa da espécie Riemerella anatipestifer, um patógeno causador de
44
44
Além das funções antibacterianas observadas, os PAMs de peixes também demonstram
outras atividades. O peptídeo β-defensina (BD)-1 mostrou atividade contra a septicemia
hemorrágica viral (SHV), infecção que acomete uma espécie de truta denominada Oncorhynchus
mykiss (Falco et al., 2009). Os PAMs, HT 1-5 e epinecidina-1 isolados de tilápia e garoupa,
respectivamente demonstraram também funções antivirais contra o vírus da necrose do sistema
nervoso (VNSN) (Chia et al., 2010; Wang, Kung e Chen, 2010). Devido à eficiência observada
nos ensaios antivirais utilizando epinecidina-1, a necessidade de investigações sobre o
mecanismo de ação destes peptídeos para o uso terapêutico, no combate à infecções virais
apresenta uma área emergente (Wang et al., 2010). O tratamento com HT 1-5 protegeu embriões
de salmão de uma infecção no pâncreas causada pelo vírus IPNV (infectious pancreatic necrosis
virus), grande causador de perdas econômicas para a indústria de exportação deste rentável peixe
(Rajanbabu e Chen, 2011a). A possibilidade de usar PAMs como modelo para o desenvolvimento
de moléculas no melhoramento da aquicultura também mostra ser uma área promissora.
A ação antifúngica também foi encontrada em alguns PAMs isolados de peixes. O
peptídeo, piscidina-2, secretado pelo robalo, tem a capacidade de romper as membranas de
fungos e agir como fungicida (Sung, Lee e Lee, 2008). Já o peptídeo derivado de histona isolado
do bacalhau do Atlântico, o H2B, foi caracterizado com funções antibacteriana e antifúngica
contra os agentes patogênicos Aeromonas hydrophilia, bactéria Gram-negativa e Saprolegnia
spp. fungo aquático (Bergsson et al., 2005). Outra função observada nos PAMs de peixes foi
demonstrada pela β-hemoglobina, peptídeo com atividade antiparasitária contra a espécie
Ichthyophthirius multifiliis (ich), causador de ictiofitiríase em bagres (Ullal e Noga, 2010).
45
protozoário, anaeróbico flagelado que causa tricomoníase e Candida albicans, um fungo
diplóide, causador de infecções oportunistas orais e genitais em seres humanos (Pan et al., 2009)
(Figura 6).
Figura 6. Estrutura tridimensional do peptídeo piscidina (pdb 2jos) um peptídeo catiônico com potente atividade antimicrobiana, apresentando uma estrutura composta por uma α-hélice. A visualização foi realizada utilizando o programa PyMol v0.99.
Em adição, outros exemplos de PAMs de peixe também são relatados na literatura,
entretanto suas estruturas tridimensionais ainda são pouco exploradas ou não foram resolvidas.
No Protein Data Bank além da piscidina relatada anteriormente, somente pardaxina e
pleurocidina estão depositadas. Pardaxina (pdb 1xc0) consiste em peptídeo antibacteriano ativo
contra bactérias Gram-positiva e -negativa in vitro e secretado por glândulas da mucosa do peixe
Pardachirus pavoninus (Porcelli et al., 2004). Pardaxina biofisicamente interage com membranas
celulares por meio da formação de poros ou disrupção celular de canais iônicos voltagem
46
46
aminoácidos carregados positivamente, mas na somatória geral a carga das isoformas para este
peptídeo varia de 0 a +1 (Figura 7A). Já o peptídeo pleurocidina (pdb 1z64) apresenta carga +4 e
estrutura em α-hélice na presença de TFE ou DPC (Syvitski et al., 2005). Este peptídeo foi
isolado da secreção da pele do peixe Pleuronectes americanus e apresentou atividade
antibacteriana (Figura 7B).
Figura 7. Estrutura tridimensional do peptídeo (A) pardaxina (pdb 1xc0) e (B) pleuricidina (pdb 1z64) peptídeos catiônicos com atividade antimicrobiana contra bactérias Gram-positiva e -negativa, apresentando conformação
estrutural composta por α-hélice. A visualização foi realizada utilizando o programa PyMol v0.99.
(A)
47
1.7MECANISMO DE AÇÃO PARA PEPTÍDEOS ANTIMICROBIANOS
Considerando que os PAMs se comportam como barreiras naturais contra infecções
bacterianas, o conhecimento da enorme variedade de mecanismos de ação destas moléculas
bioativas fornece informações importantes para o desenho de peptídeos com potencial
terapêuticos. Os mecanismos de ação de PAMs contra bactérias são atualmente os melhores
estudados. Inclui a inibição da formação da parede celular, formação de poros na membrana
celular resultando na interrupção do potencial de membrana com eventual lise da célula e
inibição da síntese de ácidos nucleicos, de proteínas, de proteases e da parede celular (Hancock e
Scott, 2000; Zasloff, 2002; Brogden, 2005). Um dos modelos mais aceitos para explicar a
atividade dos PAMs consiste no modelo proposto Shai-Matsuzaki-Huang (SMH), o qual propõe a
interação do peptídeo com a membrana, seguido pelo deslocamento de lipídios, alteração na
estrutura da membrana e em certos casos, a entrada do peptídeo no interior da célula-alvo
48
49
Durante muitos anos tem se discutido sobre o mecanismo de ação dos PAMs.
Acreditava-se que a ação desAcreditava-ses peptídeos era única e exclusivamente sobre a membrana celular. Entretanto,
hoje se sabe que os PAMs, além de suas propriedades permeabilizantes e disruptivas, também
podem operar por meio de interações com alvos intracelulares ou interrupção dos principais
processos celulares promovendo uma modulação no sistema imunológico do hospedeiro
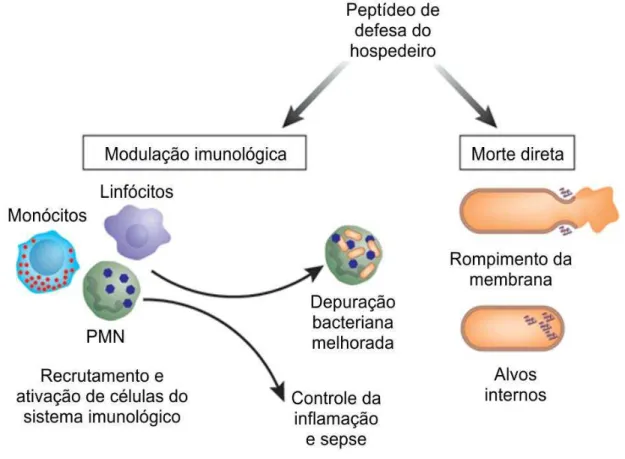
recrutando e ativando células de defesa como linfócitos e monócitos (Figura 9) (Nicolas, 2009).
Figura 9. Visão geral do amplo espectro de ação dos peptídeos de defesa. Os peptídeos de defesa além de sua ação
50
50
Independentemente do mecanismo de ação, a atividade dos peptídeos antibacterianos são
dependentes das interações entre o peptídeo e a membrana celular (Hancock e Rozek, 2002).
Sendo que os mecanismos de ação dos PAMs podem variar de acordo com a célula com a qual
interage, o que depende da composição e característica das membranas celulares. As membranas
bacterianas, por exemplo, são organizadas de tal forma que a porção hidrofílica da bicamada
composta por fosfolipídios é densamente carregada com cargas negativa. Por outro lado, a
membrana externa da bicamada em plantas e animais pode ser composta em geral por lipídeos
com nenhuma carga líquida e na maioria dos lipídios carregados negativamente, estes estão
presentes na membrana interna da bicamada (Matsuzaki, 1999; Zasloff, 2002).
Uma das características que torna os PAMs moléculas bastante atraentes quando se pensa
em seu uso terapêutico consiste na seletividade tóxica apresentada por muitos PAMs. Muitos
deles são completamente inativos a células eucarióticas, e a base para essa seletividade parece
estar relacionada com a composição lipídica da membrana-alvo (fluidez, densidade de carga
negativa, e a ausência e/ou presença de colesterol) (Nicolas, 2009). A presença de colesterol na
membrana, geralmente reduz a atividade dos PAMs, uma vez que o colesterol auxilia na
estabilização da bicamada lipídica, podendo haver ainda interações entre o colesterol e os PAMs
de forma que não haja disrupção da membrana (Nguyen, Haney e Vogel, 2011).
1.8 PEPTÍDEOS MULTIFUNCIONAIS
Nas últimas décadas, apesar de enormes avanços na biologia molecular, os pesquisadores
51
No entanto, nos últimos anos, muitos pesquisadores têm demonstrado que a vida não pode ser
vista como uma via de sentido único, sendo mais bem explicada como uma rede de interações,
um sistema com muitos pontos de partida (Franco, 2011). Esta ideia expansiva também pode ser
extrapolada para estrutura de proteínas. A visão convencional de que as proteínas e peptídeos
possuem uma estrutura única diretamente relacionada a uma função única confronta com a
capacidade destas proteínas em adaptarem e desempenharem novas funções.
A denominação de multifuncionalidade pode ser dada para determinado objeto que realiza
sozinho várias funções. No caso de proteínas e peptídeos multifuncionais essa denominação vem
sendo bastante empregada nos últimos anos para proteínas e peptídeos que sozinhos
desempenham múltiplas funções, tais como interações com membranas de bactérias causando
disrupção e no recrutamento de macrófagos, neutrófilos uma ação definida como
imunomodulação (Franco, 2011). Essas múltiplas funções estão sempre associadas com uma
única estrutura tridimensional, com homólogos semelhantes, e tem vindo a ganhar a atenção em
vários campos de investigação, incluindo a imunologia e enzimologia (Nobeli, Favia e Thornton,
2009). A quadro 3 mostra alguns exemplos e funções de peptídeos multifuncionais isolados de
52
52 Quadro 3. Peptídeos antimicrobianos com outras atividades.
Nome Organismo Função Referência
Mastoparam-X Vespula lewisii AB; CM; IM (Hirai et al., 1979)
Magainina-2 Xenopus laevis AB, AV, AF, AP, AC, IM (Ramos et al., 2012) Foulicidina Gallus domesticus AB, AF, AV, IM (Xiao et al., 2006)
LL-37 Homo sapiens AB, AV, AF, CM, IM (Into et al., 2010)
Cn-AMP1 Cocos nucifera AB, AF, AF, IM (Silva et al., 2012)
Kalata-b2 Oldenlandia affinis AB, UT, IM (Mylne et al., 2010)
Cr-ACP1 Cycas revoluta AB, AF, IM (Mandal et al., 2012)
Antibacteriana: AB; antiviral: AV; antifúngica: AF; anticâncer: AC; antiparasitária: AP; Células de mamíferos: CM; Imunomodulatória: IM e uterotônica: UT.
Além de todas estas atividades diretas bem caracterizadas para PAMs isolados de peixes
sobre agentes patogênicos, estes peptídeos possuem a capacidade de penetrar para o meio interno,
ou ligar-se à receptores na membrana da célula hospedeira como visto anteriormente com alguns
exemplos de PAMs isolados de diferentes fontes. A síndrome de Crohn e Kostmann, uma
deficiência causada pela ausência de defensinas, apresenta avanços no tratamento com PAMs
isolados de peixes (Kindrachuk e Napper, 2010). Outro estudo interessante foi desenvolvido com
o peptídeo isolado da truta arco-íris, a qual foi usada como modelo contra SHV, um vírus que
acomete a aquicultura de peixes comerciais. Os resultados obtidos possibilitaram identificar uma
ação similar à defensina humana alfa-1. HNP1, o peptídeo isolado da truta arco-íris revelou
eficiência no combate a este vírus modulando as vias pró-inflamatórias pelas citocinas,
interleucinas (IL)-1, IL-8, o fator de necrose tumoral (TNF)-α1; e quimiocinas, CK5B, CK6 e
CK7A; e as expressões gênicas de interferon-1 e interferon-3 (Falco et al., 2009).
Outro exemplo é observado nas defensinas de peixes as quais podem servir como
alternativas potenciais em indivíduos com doenças genéticas, como no caso da expressão de
53
resposta imune. O resultado permitiu demonstrar que a modulação da resposta imune foi
ocasionada devido à expressão de genes de citocinas causado pelo efeito do PAM, resultado do
recrutamento de macrófagos, e a modulação da ciclo-oxigenase (COX)-2 (Chiou et al., 2006). O
peptídeo HT 2-3 transgênico da espécie Danio rerio mostrou efeito sobre a expressão diferencial
de genes imunomoduladores relacionados a infecções causadas pela espécie V. vulnificus (Hsieh,
Pan e Chen, 2010). Em particular o peptídeo foi relacionado com o “up-regulated” de 10,
IL-26, receptor de lisozima “toll-like” (TLR)-4a, e MyD88 e com o “down-regulated” de IL-1β e
IL-15 após a infecção bacteriana (Hsieh, Pan e Chen, 2010). A modulação da resposta à patógenos
por meio de genes e expressão de proteínas podem reduzir a gravidade ou permitir a completa
proteção contra infecções por patógenos. Além disso, o tratamento com PAMs contra agentes
patogênicos induz a imunidade adaptativa e reduz a gravidade de re-infecção pelo patógeno.
Outra via também explorada sobre imunomodulação consiste na alteração da transcrição mediada
pela injeção de hepcidina em rato (De Domenico et al., 2010). O pré-tratamento com hepcidina
permitiu a sobrevivência de ratos contra infecção causada pela injeção de LPS, em modelos de
sepse em camundongos realizado pela ligadura e perfuração cecal. Portanto, a hepcidina
demonstrou promissoras possibilidades no uso deste peptídeo como agente profilático para
prevenir infecção bacteriana através da modulação do sistema imune (De Domenico et al., 2010).
As expressões dos genes de AMP H2A e parasina-1 foram relatadas sendo reguladas por
proteínas imuno-relacionadas (Cho et al., 2002). O perfil modulatório de transcrição e tradução
do peptídeo HT 2-3 utilizando a técnica de “microarray” foi avaliada por meio de células de
macrófagos de ratos estimuladas por LPS, neste trabalho ficou demonstrado a “down-regulated”
54
54
2-3-1 e epinecidina mostraram atividade antitumoral in vitro, após a aplicação contra células de
fibrossarcoma humana (Chen, Lin e Lin, 2009; Chen et al., 2009; Lin et al., 2009). HT 2-3-1 e
epinecidina tem ação na lise, inibição da proliferação e na redução da migração de células
tumorais (Chen, Lin e Lin, 2009; Lin et al., 2009). Em certas concentrações de HT 2-3-1 não foi
possível identificar efeitos citotóxicos para linhagens de células normais, mas foi possível inibir a
proliferação e migração de células anormais (Chen, Lin e Lin, 2009). Esta pode ser uma
abordagem promissora na pesquisa contra o câncer e vem sendo outra área de grande interesse
para os pesquisadores. Além desta habilidade observada, uma função antitumoral contra células
leucêmicas humanas U937 e uma função antinecrose para epinecidina-1 foram relatadas em
testes in vitro (Chen et al., 2009). Foi possível identificar que a epinecidina-1 modula a
caspase-3, caspase-8 e caspase-9, e induz a apoptose em células tumorais (Chen et al., 2009). No entanto,
estudos adicionais associados com o desenvolvimento clínico são necessários para demonstrar a
real potencialidade dos PAMs encontrados em peixes no controle de tumores.
1.9 PLEURONECTES AMERICANUS
Pleuronectes americanus consiste em um peixe de inverno conhecido popularmente como
solha (vem do latim e significa sola), este animal apresenta uma característica peculiar, olhos
voltados para o mesmo lado e que compreende quase todas as formas inteiramente ossificadas
(subclasse Teleósteos) (Figura 10A). Esta espécie tem sido classificada como representante da
55
regiões de águas geladas do Canadá e da América do Norte (Figura 10B). São peixes que
apresentam além de seu valor esportivo para pescadores, também a presença de PAMs isolados
de sua pele e intestino. Entretanto, o que também chama a atenção nesta espécie são moléculas
denominadas de peptídeos anticongelantes (AFP) que conferem a este animal a capacidade de
sobreviver em águas onde a temperatura fica abaixo de zero, tornando a espécie P. americanus -
um grande alvo no isolamento destas moléculas para estudos de criopreservação e/ou
crioproteção.
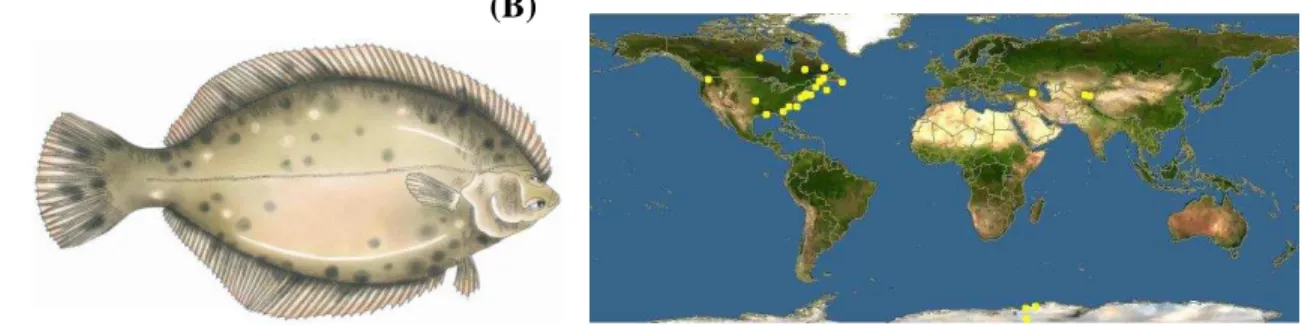
Figura 10. Figura ilustrativa do (A) Pleuronectes americanis, espécie de peixe ósseo com os dois olhos situados do mesmo lado. Fonte: http://narragansett-bay.com/index.cfm. (B) Mapa Mundi, ressaltando em pontos amarelos a localização do P. americanus.
Nas regiões polares, a água do mar fica consistentemente abaixo da temperatura de ponto
de congelamento das soluções fisiológicas, atingindo temperaturas de aproximadamente -1.9 °C
(Davies e Hew, 1990). Os peixes que habitam estas regiões desenvolveram estratégias para a
sobrevivência, sendo capazes de expressarem moléculas que impedem o congelamento ou danos
em seus órgãos. Estudos destes compostos encontrados no sangue destes peixes da Antártica
mostraram que uma molécula de origem proteica era a responsável por este comportamento
incomum (Devries e Wohlschlag, 1969; Devries, Komatsu e Feeney, 1970).