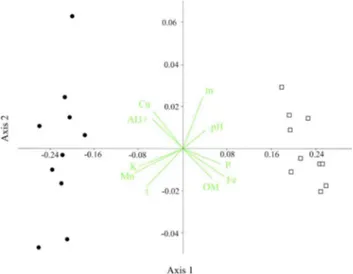
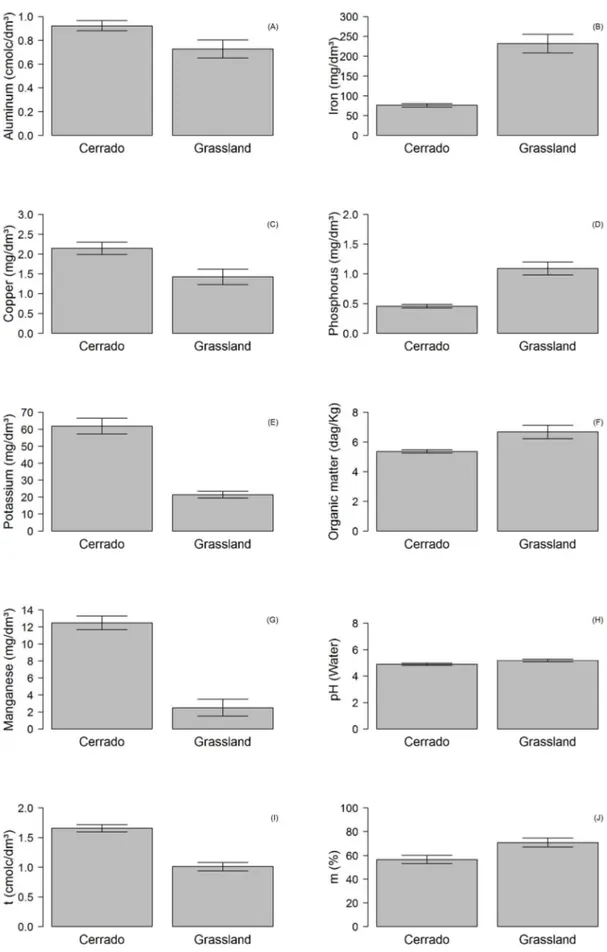
Together yet separate: variation in soil chemistry determines differences in the arboreal-shrub structure of two contiguous rupestrian environments
Texto
Imagem

Documentos relacionados
A participação na escola refere-se à participação em atividades no seio da instituição, que no entanto, não estão diretamente relacionadas com as aulas de Educação Física,
Regarding the Mediterranean biogeographical region, 53 SCIs have been provisionally adopted within Natura 2000 in Mainland Portugal (Commission of the European Communities 2006)..
Para determinar o teor em água, a fonte emite neutrões, quer a partir da superfície do terreno (“transmissão indireta”), quer a partir do interior do mesmo
Extinction with social support is blocked by the protein synthesis inhibitors anisomycin and rapamycin and by the inhibitor of gene expression 5,6-dichloro-1- β-
[Objective] In this work, we propose a Requirements Engineering (RE) approach for DSPL, ConG4DaS (Contextual Goal models For Dynamic Software product lines), which
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
