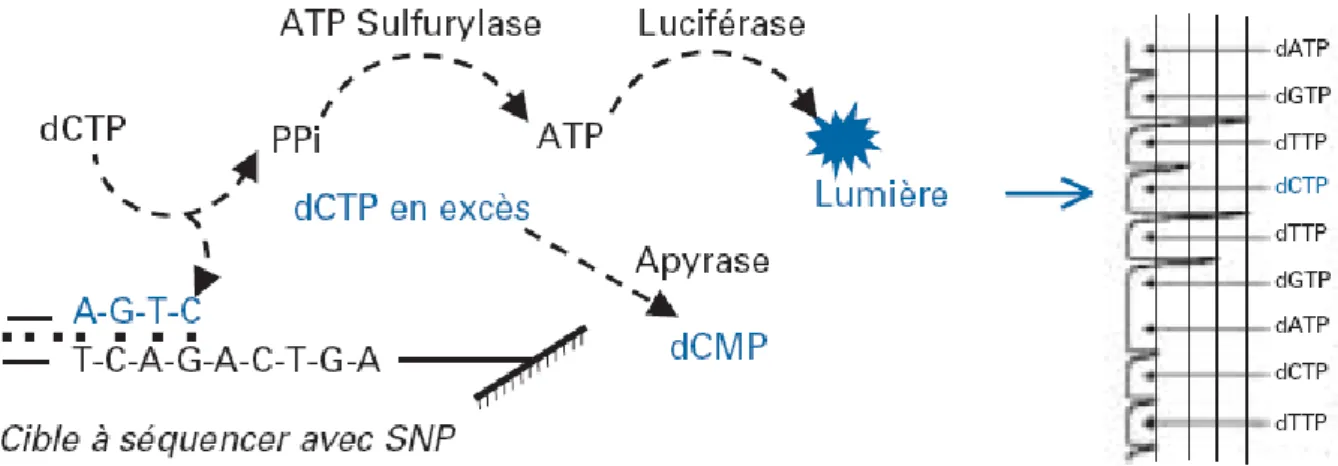
L’intégrité structurelle de l’ADN est nécessaire au succès de la transcription et de la réplication de l’ADN. Un effet similaire a été obtenu via la caséine kinase 2 (CK2) (Schneider et al., 2002). L'expression de la GFP (Green Fluorescent Protein) est analysée par cytométrie en flux (FACS Beckman Coulter).
Résultats de quantification relative de l'expression de gènes plasmidiques (blasticidine-GFP et XPD-V5) intégrés dans le génome de différents clones cellulaires sélectionnés. Pour cela, nous avons choisi un contrôle positif constitué d'un extrait protéique de la lignée K562.
Vérification de la localisation par microscopie confocale 82
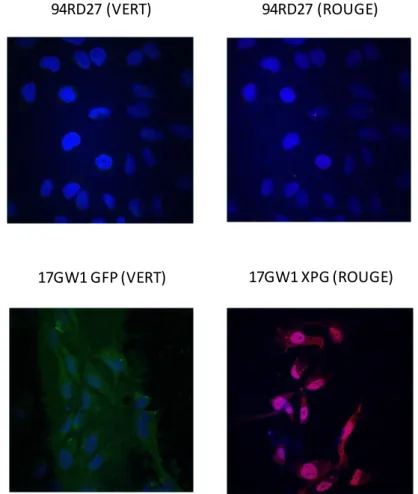
83 De la même manière que nous l'avons fait pour les transfections transitoires, nous avons observé l'expression de la GFP et du transgène dans les clones stables par microscopie confocale (Figure 27) pour compléter l'analyse cytométrique qui a permis d'identifier les clones fluorescents, ainsi que le pourcentage de cellules fluorescentes. La microscopie confocale permet également d'évaluer l'homogénéité des clones en observant la fluorescence de différents champs, et de déterminer si les cellules GFP-positives sont également positives pour le transgène, qui dépend d'un promoteur différent. La blasticidine-GFP dépend d'un promoteur CMV, tandis que le gène d'intérêt dépend d'un promoteur EF1-α.
Microscopie confocale de fibroblastes 94RD27 non transfectés et du clone 17GW1 transfectés de manière stable avec le vecteur pTRACER-ERCC5-WT. Nous avons retenu le clone 17GW1 qui a été obtenu par transfection stable du plasmide pTRACER-ERCC5-WT dans des cellules 94RD27 déficientes en XPG et qui montre une expression homogène du transgène dans les cellules observées. Nous avons fait de même pour tous les clones positifs en cytométrie et en Western blot.
Tests fonctionnels 83
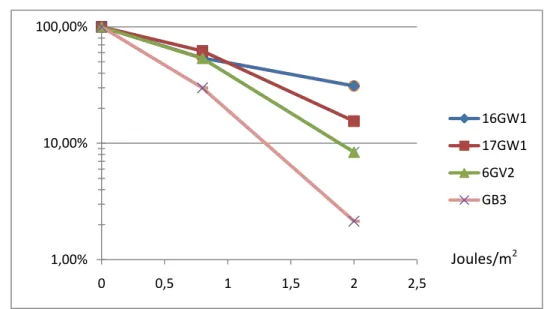
GB3 : population cellulaire transfectée avec le vecteur « vide » ; 6GV2 : clone exprimant la protéine variante XPG. Lors de cette vérification de la fonctionnalité de la complémentation des clones XPG, nous avons observé une différence de survie entre les clones sauvages et le clone variant testé. Pour confirmer cette observation, nous avons répété cette expérience en triple sur deux clones XPG sauvages et deux clones XPG variants (Figure 30).
Nous avons montré que l’activité de réparation de la protéine XPG variante (1104asp) était inférieure à celle de la protéine commune (1104his). 16GW1 et 17GW1 : clones exprimant la protéine XPG générale ; 6GV2 et 7GV2 : clones exprimant la protéine variante XPG. Etude de l'effet des polymorphismes XPD lys751gln et XPG asp1104 sy sur la réponse cellulaire aux agents cytotoxiques.
Étude de l’effet des polymorphismes XPD lys751gln et XPG asp1104his
Test de cytotoxicité par mesure de l’inhibition
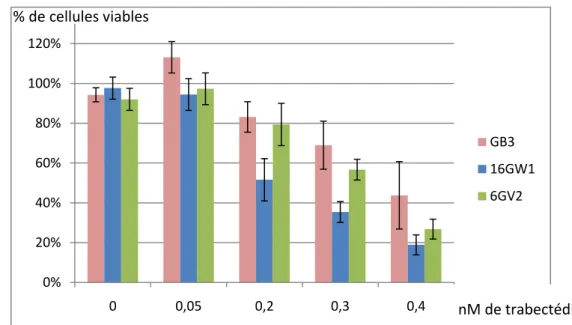
GB3 correspond à une population cellulaire transfectée avec le vecteur « vide » ; 6GV2 correspond à un clone exprimant la protéine variante XPG ; 16GW1 correspond à un clone exprimant la protéine. Courbe d'inhibition de la croissance par la trabectédine de GB3 : population cellulaire transfectée avec le vecteur « vide » ; 6GV2 : clone exprimant la protéine variant XPG ; 16GW1. Le manque de protéine XPG fonctionnelle rend la lignée 94RD27 hypersensible aux UV, mais aussi aux molécules qui créent des adduits à l'ADN comme le cisplatine.
Courbe d'inhibition de la croissance du cisplatine ; GB3 correspond à une population cellulaire transfectée avec le vecteur « vide » ; 7GV2 correspond à un clone exprimant la protéine. 89 Les tests de cytotoxicité du paclitaxel, de la trabectédine et du cisplatine, présentés dans les figures 31, 32 et 33, montrent d'une part que la complémentation de la lignée déficiente par la forme régulière ou variante de la protéine XPG a un effet sur l'inhibition de la croissance provoquée par ces agents anticancéreux ; et d'autre part que l'on observe une réponse cellulaire différente selon le type de complémentation réalisée. Alors que la complémentation avec la protéine commune XPG (16GW1) sensibilise les cellules au paclitaxel et à la trabectédine, l'effet inverse est observé pour le cisplatine, pour lequel cette complémentation est associée à une moindre sensibilité.
La complémentation par la protéine variante XPG (6GV2) est également associée à la résistance cellulaire au cisplatine ; cette résistance est moins importante que celle associée à la complémentation avec la protéine commune. On observe également, dans une moindre mesure qu'avec la complémentation avec la protéine normale, une sensibilisation des cellules à la trabectédine.
Test de cytotoxicité par mesure de l’inhibition de croissance
Il apparaît donc que le polymorphisme du codon 751 n'est pas responsable d'une modification de la stabilité de l'ARNm. Activité CDK1 dans la lignée MCF7, pendant le traitement au paclitaxel, en fonction de l'inactivation de la protéine XPD (C = siARN témoin ; si = siARN XPD). Activité CDK1 dans la lignée XP6BE pendant le traitement au paclitaxel en fonction de l'inactivation de la protéine XPD.
Inhibition de la croissance par le cisplatine de la lignée MCF7 traitée avec des siARN témoins ou ciblant XPD. 99 Nous avons également évalué l'inhibition de la croissance par le cisplatine de la lignée XP6BE après avoir inhibé XPD avec du siARN. La variation du niveau d'expression de la protéine XPD n'a dans ce cas aucune influence sur la réparation (Figure 44).
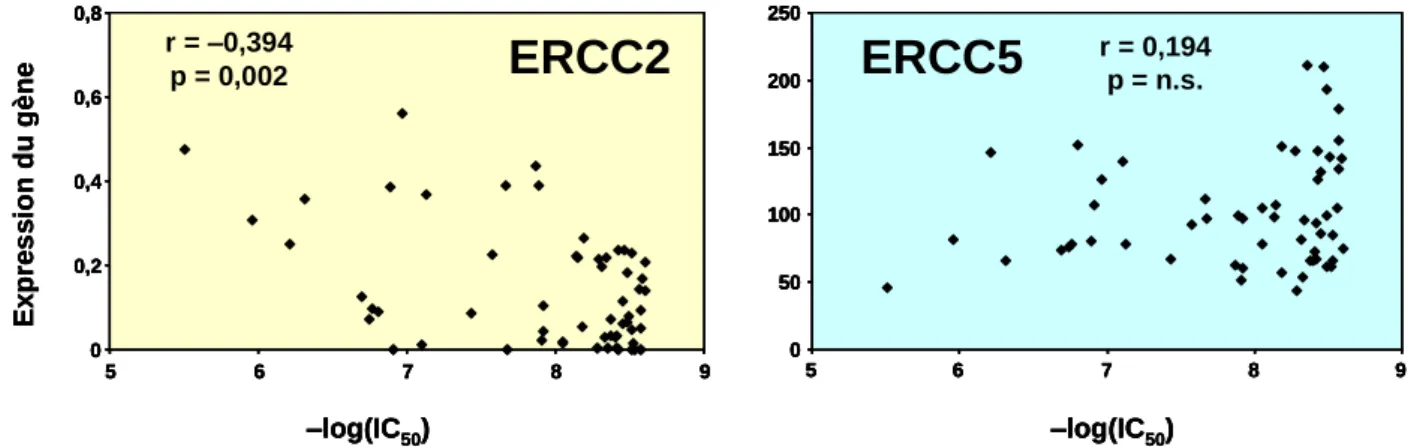
Inhibition de la croissance par le cisplatine de la lignée XP6BE traitée avec du siRNA contrôle ou ciblant XPD. De plus, nous avons montré que le polymorphisme du codon 751 du gène ERCC2 est associé à une diminution de l’expression de la protéine messagère et XPD.
Étude des messagers commun et variant du gène ERCC2 (XPD) 90
Étude de la structure secondaire de l’ARNm de ERCC2 90
Pour étudier la structure secondaire de l'ARNm d'ERCC2, nous avons réalisé une approche in silico à l'aide du logiciel RNAfold. Un changement dans l'unique nucléotide affecté par le polymorphisme lys751gln altère significativement la structure secondaire de l'ensemble du messager ERCC2 (Figure 35), tandis qu'une variation de séquence provoquée par un autre polymorphisme (asp312asn), sans conséquences pharmacologiques sur la sensibilité au taxane dans le modèle NCI, ne provoque aucun changement dans la structure secondaire de l’ensemble du messager ERCC2. Ce travail a été confirmé par la publication d'une étude liant le polymorphisme à une variation significative de la quantité du transporteur ERCC2 (Wolfe et al., 2007) (Figure 36).
Ces résultats sont en faveur de l'hypothèse selon laquelle le polymorphisme du codon 751 serait responsable d'un changement dans la stabilité de ERCC2. Structure secondaire théorique des ARN messagers du gène ERCC2 produits par l'allèle de type sauvage et l'allèle variant du gène ERCC2. Des séquences partielles d'ARNm XPD (140 nt) entourant chacun des SNP ont été analysées.
Il apparaît que l'adénine (A) pour le codon 156, l'adénine (A) pour le codon 312 et la cytosine (C) pour le codon 751 permettent la structure secondaire optimale en termes d'appariement et d'énergie. requise (dG), par rapport à la séquence de référence cytosine (C), guanine (G) et adénine (A) respectivement pour les mêmes codons. Nous avons tenté de valider cette hypothèse en : (1) recherchant une différence dans la stabilité de l'ARN messager dans les lignées présentant un génotype variant commun homozygote, homozygote ou hétérozygote pour le codon ERCC2 751 SNP, (2) en l'évaluant dans des lignées hétérozygotes pour l'ERCC2. gène, la transcription de chacun des deux allèles de ERCC2, recherchant un taux de transcription réduit pour l'allèle 751gln.
Étude de la stabilité des ARNm de ERCC2 92
Nous n’avons donc mis en évidence aucune différence notable dans la stabilité de l’ARNm XPD qui pourrait expliquer la plus faible expression de la protéine variante. Si ce n’était pas le cas, cela signifierait que le SNP au niveau du codon 751 ne serait qu’un marqueur non fonctionnel du niveau d’expression de XPD. Il est possible que la stabilité des messagers soit effectivement impliquée, mais pas de manière différente dans les trois lignées que nous avons étudiées, c'est-à-dire qu'elles ne porteraient pas le SNP responsable du changement de demi-vie de l'ARNm.
Le SNP711 provoque des effets similaires (in silico) à ceux du SNP751 sur la structure secondaire de l'ARNm. Il est également possible que la stabilité du messager ne soit pas en jeu : il y a peut-être un SNP dans la partie promoteur qui serait responsable de la variation de l'expression de XPD et qui serait génétiquement lié au SNP 751. extension de cette étude Il serait nécessaire de connaître avec certitude l’impact de ces polymorphismes sur la stabilité de l’ARNm d’ERCC2.
Étude du rôle cellulaire de la variation du niveau de la protéine XPD 93
- Choix de la méthode d’extinction - Mise au point de l’extinction 93
- Cinétique d’extinction de la protéine 94
- Étude du cycle cellulaire en fonction de l’extinction de XPD 94
- Étude de l’activité de CDK1 (cdc2) en fonction de l’extinction de XPD 94
- Tests d’inhibition de croissance (MTT) des lignées MCF7et XP6BE 96
Western blot montrant la cinétique de disparition de la protéine XPD entre 48 et 96 h après transfection par siRNA dirigé contre le messager ERCC2, dans la lignée XP6BE. Pour étudier l'influence de la protéine XPD sur le cycle cellulaire, nous avons entrepris une analyse du cycle cellulaire dans la lignée MCF7 par silençage en phase G2/M. Ainsi, nous avons voulu vérifier dans des lignées humaines que la disparition de la protéine XPD par la méthode antisens pouvait favoriser l'activation du complexe MPF (CDK1/cycline B).
La régulation de l'activité CDK1 par XPD/CDK7 ne semble pas se produire ici par dégradation plus ou moins tardive de la cycline B. Disparition de . Inhibition de la croissance par le paclitaxel des cellules traitées avec le siRNA ciblant XPD Nous avons montré que le traitement de la lignée MCF7 avec le siRNA XPD la rendait moins sensible au paclitaxel (Figure 45).
De plus, son expression basale de XPD est relativement faible ; ainsi, malgré son génotype XPD commun, il semble difficile de sensibiliser davantage cette lignée au paclitaxel par la perte de la protéine XPD. Courbe d'inhibition de la croissance par paclitaxel de la lignée XP6BE par ciblage siRNA contrôle ou XPD.

Choix de l’étude 100
Description de l’étude clinique 100
Étude statistique 100
Comparison of the global genomic and transcription-coupled repair rates of different lesions in human cells. Impairment of the TFIIH-associated CDK-activating kinase selectively affects cell cycle-regulated gene expression in fission yeast. Binding to DNA of the RNA polymerase II C-terminal domain allows the distinction between Cdk7 and Cdk9 phosphorylation.
Fine-tuning the cell cycle: activation of the Cdk1 inhibitory phosphorylation pathway during mitotic exit. An intronic poly (AT) polymorphism of the DNA repair gene XPC and risk of head and neck squamous cell carcinoma: a case-control study. Cell cycle analysis of the activity, subcellular localization and subunit composition of human CAK (CDK-activating kinase).
XPD DNA repair gene polymorphisms: correlations with risk of basal cell carcinoma revisited. Levels of DNA repair proteins in the face of anticancer drug resistance in human tumor cell lines of the National Cancer Institute Drug Screening Program.