Je tiens également à remercier Jean-François HOCQUETT pour m'avoir accueilli au sein de l'Unité de Recherche sur les Herbivores et pour sa collaboration dans diverses communications. Je remercie Daniel TAILLANDIER de l'Unité de Nutrition Humaine de m'avoir appris à réaliser la détection de fluorescence.
COMMUNICATIONS
Analyse des marqueurs de tendreté du bœuf : étude de la fonctionnalité des gènes et analyse du polymorphisme. Comparaison de la tendreté du bœuf basée sur la validation de marqueurs potentiels.
ABREVIATIONS
Figures 31A, 31B et 31C : Prélèvement de poils de queue de bovin en vue de réaliser un test de prédiction de la tendresse animale (Igenity TenderGene).
LISTE DES TABLEAUX
CONTEXTES, ENJEUX, ET OBJECTIFS DE LA THESE
En conséquence, il est difficile pour l’industrie bovine de contrôler les variations de tendreté. Il est donc nécessaire de fournir des outils à l'industrie pour contrôler la tendreté des viandes.
INTRODUCTION BIBLIOGRAPHIQUE
I Les bovins
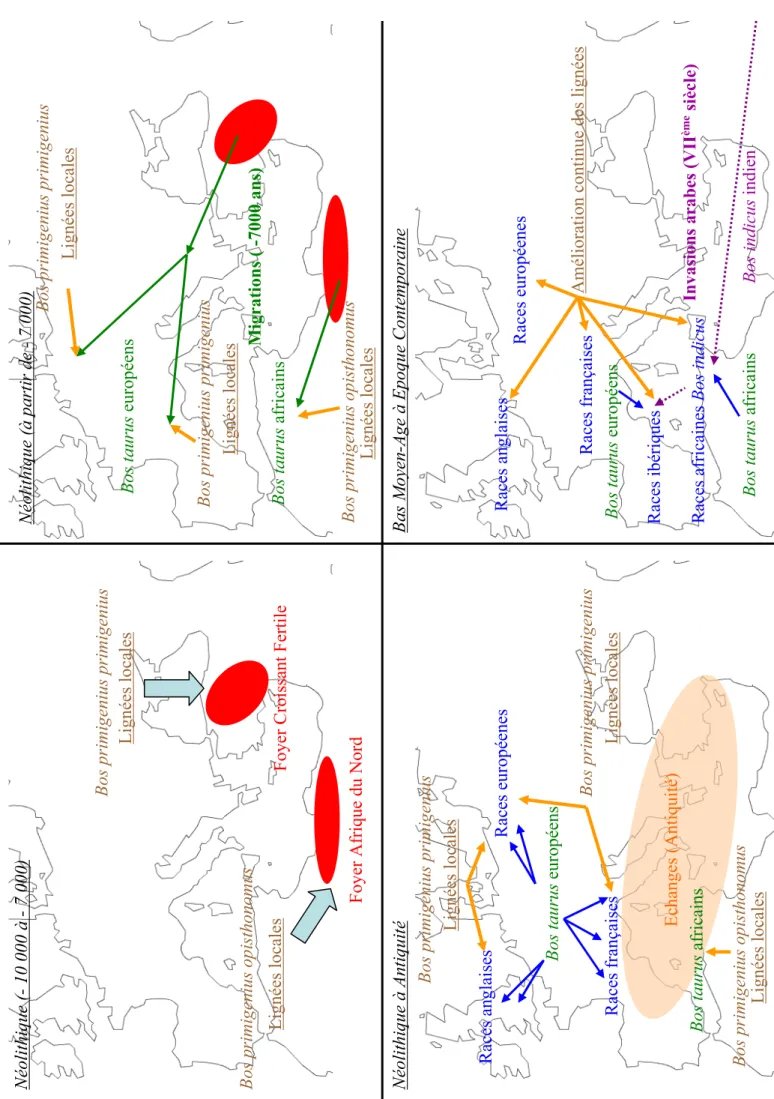
- Les foyers de domestication
- Foyer de l’Inde
- Foyer Nord-Africain
- Influences d’Homo sapiens sur les lignées Bos taurus et Bos indicus
- Bovins africains
- Bovins européens
- Conséquences pour Homo sapiens
- Civilisation
- Génétique et physiologie
- Mythologie
- Les différentes races bovines et l’exception française
Des études sur l'ADN mitochondrial ont montré une séparation nette entre les lignées du Bos indicus indien d'une part, et celles du Bos taurus européen et du Bos indicus africain d'autre part (Loftus et al. 1994, Bradley et al. 1996). Les analyses des haplotypes du chromosome Y montrent une séparation entre les lignées européennes de Bos taurus d'une part, et les lignées indiennes et africaines de Bos indicus d'autre part (MacHugh et al. 1997).
II Le muscle squelettique strié
Par conséquent, les fibres ont été classées selon l'activité de l'ATPase myofibrillaire (mATPase), après incubation des fibres à un pH acide ou basique ( Brooke et Kaiser 1970 ). Ainsi, chez les mammifères, les fibres à contraction lente (type I) sont constituées de MyHC de type I.
Physiologie
Morphologie
Métabolites
Propriétés enzymatiques
Types de fibre
Les myofibrilles
La bande A est caractérisée par des filaments de myosine superposés avec des filaments d'actine. Le chevauchement des filaments fins ne s'étend pas sur toute la longueur des filaments épais, il existe une zone où les filaments de myosine sont seuls : c'est la bande H. Les structures protéiques qui maintiennent les filaments épais forment la raie M. au sein du groupe A.
Les sarcomères
27 La force de contraction produite par le raccourcissement des sarcomères se propage à travers la fibre grâce aux connexions entre la myofibrille et la fibre.
Les costamères
La myomésine et la protéine M sont des composants structurels de la lignée M (Obermann et al 1996). L'obscurine participe à la formation de la lignée M (Kontrogianni-Konstantopoulos et al 2004) et à son ancrage au réticulum sarcoplasmique via l'ankyrine (Lange et al 2006). La protéine MURF-1 liée à la titine régule la dégradation des protéines musculaires et participe au maintien de la lignée M.
Le disque Z
En effet, la lignée M renforce le rôle stabilisant de la titine, une protéine qui traverse le sarcomère en connectant le disque Z à la lignée M et en ramenant ces structures à leur position d'origine après contraction. Les souris KO pour la myomésine ou la protéine M n'ont pas pu être produites à l'heure actuelle, mais des knock-outs conditionnels ont été produits pour le domaine titine qui reconnaît la lignée M. L'activation de KO conduit au démontage des sarcomères et à la mort de la souris, démontrant la nature essentielle de la lignée M.
Les filaments épais
En effet, les têtes de myosine (partie N-terminale de la chaîne lourde) sont orientées vers les filaments d'actine, et les queues (partie C-terminale) vers la ligne M. En fait, le filament d'actine n'est pas fixe, il existe une association /dynamique de dissociation. La régulation de cette dynamique est essentielle pour maintenir une longueur uniforme du filament fin.

Nébuline – CapZ – Tropomoduline – Tropomyosine – Troponine
- Calpaïnes
- Protéasome 20S
- Caspases
- Matrice metallopeptidase
- Peptidase à sérine
- La contraction musculaire
- Le métabolisme énergétique de la fibre musculaire
- La glycolyse
- Production de lactate
- La voie aérobie
- Intégration des différentes voies
- Les protéines chaperonnes
- Hsp20
- Hsp40
- Hsp60
- Hsp70
- Hsp90
- Hsp100
- Quelques mécanismes
- L’apoptose
Cette dernière recouvre alors le site de liaison de la myosine sur l'actine, empêchant ainsi un cycle de contraction. Dans les cellules musculaires, cela implique la fermentation de l’acide lactique, qui a lieu dans le cytosol de la fibre. Le métabolisme énergétique des fibres musculaires ne s’effectue pas par une seule voie.

IIILes qualités sensorielles de la viande bovine
- La couleur
- La flaveur
- La jutosité
- La tendreté
- Caractéristiques de la tendreté
- Les fibres musculaires
Le test sensoriel utilise un jury de dégustation formé pour évaluer la tendreté du bœuf. L'évaluation de la sensibilité est problématique car il existe de nombreux tests différents. Le test WB est en corrélation avec la dureté sensorielle de la viande (r = 0,82) et est donc l'inverse de la tendreté (Peachey et al 2002).

TENDRETÉ
- Le collagène
- Les lipides
- Les systèmes protéasiques
- L’apoptose
- Conclusion
De plus, l’importance du collagène pour la tendreté varie en fonction de la température de cuisson de la viande. Le rôle des cathepsines dans l'établissement de la sensibilité fait l'objet de controverses. L'apoptose serait donc un processus impliqué dans l'établissement de la tendresse (Ouali et al 2006).
IV Génomique et tendreté
Le génome bovin
Le séquençage du génome bovin a été initié en 2003 par le Center for Human Genome Sequencing du Baylor College of Medicine à Houston (Texas, USA) et a été publié le 24 avril 2009 (Elsik et al 2009). Actuellement, le génome bovin a été séquencé et assemblé, mais il est mal annoté. Les bovins étant étroitement apparentés aux moutons et aux porcs, le génome bovin pourrait servir de modèle de base pour l’étude de ces animaux.
Marqueurs de la tendreté
- Marqueurs basés sur des polymorphismes .1 CAPN1
- Tests commerciaux de la tendreté
- Marqueurs basés sur la variation d’expression des gènes .1 DNAJA1
Des études ont vérifié si le gène CAPN1 pouvait être un bon marqueur de susceptibilité (Page et al 2002). L'activité CAST est donc corrélée à une réduction de la tendreté de la viande (Schenkel et al 2006). Les résultats du génotypage sont transmis à l'éleveur avec des prédictions sur la tendreté de la viande de l'animal (Figure 32), à l'aide d'une table de notation selon les génotypes (Tableau 2).

V La protéomique et la tendreté des viandes
Définition
L'étude de Bernard et al (2007) a également mis en évidence d'autres gènes dont le profil transcriptomique était lié à la tendresse (Tableau 3). C'est par exemple le cas du gène HSPB1, dont l'expression est régulée négativement par la douceur. La sous-expression de cette protéine va accélérer la dégradation des filaments et par conséquent augmenter leur douceur.
Gène Nom Rôle dans la tendreté
Recherches sur la tendreté des viandes
- Chez le porc
C'est le cas de la protéine H liant la myosine (MyBP-H), de la chaîne légère de la myosine 1 (MLC1), du régulateur de la chaîne légère de la myosine 2 (MLC2), de la chaîne légère de la myosine 3 (MLC3), ainsi que des chaînes lentes (sTnT) et rapides ( fTnT ) troponine T. En général, les protéines impliquées dans le cycle du calcium semblent jouer un rôle dans la détermination de la douceur. Protéines identifiées dans l'étude de la tendreté de la viande chez trois races bovines, du groupe T+.

VI Bilan de l’état des connaissances
Les analyses protéomiques ont permis d'analyser de multiples profils de muscle bovin et de relier la tendreté à l'expression des protéines. Certaines protéines ont été associées à la douceur dans certaines études (comme la triisophosphate isomérase ou la chaîne légère de myosine). La relation entre l'abondance de ces protéines et la tendreté sera analysée pour établir des équations de prédiction de la tendreté et mieux comprendre les mécanismes permettant d'établir cette qualité de viande.
MATERIELS ET METHODES
I Animaux
II Caractérisation des muscles et de la viande
Données phénotypiques
Analyse mécanique – Test Warner-Bratzler
Analyse sensorielle – Estimation de la note de tendreté de la viande
Une représentation en boîte à moustaches montre que la distribution des scores pour l'année 2003 est plus large que pour l'année 2005 (Figure 34). Les mesures WB (fermeté de la viande) et TG (tendresse de la viande) sont opposées pour le muscle LT avec un coefficient de corrélation de – 0,25 (P < 0,01).

IIIQuantification des protéines
- Démarche expérimentale
- Extraction des protéines et dosage
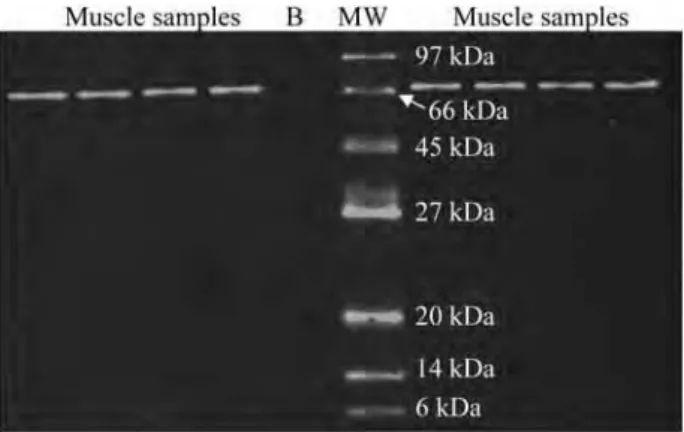
- Validation de la spécificité des anticorps par Western-Blot
- Protéines testées
- Quantification par Dot-Blot
Cette technique permet également de vérifier la spécificité de l'anticorps, c'est-à-dire si l'anticorps ne reconnaît pas d'autres cibles. Anticorps secondaire oxydant fixé à l'anticorps primaire Quantification de l'intensité du signal de bande d'intérêt par ImageQuant A. Détection par chimiluminescence Acquisition du signal à 795 nm par scanner Odyssey Figure 35. Principe de détection des anticorps par chimiluminescence. Emission d'un signal à 795 nmExcitation à 778 nm Quantification de l'intensité du signal de la bande d'intérêt par ImageQuant B. Détection par fluorescence.
IV Analyses statistiques
- Effets muscle et type d’animal
- Analyse de corrélations
- Analyse descriptive
- Analyse prédictive globale
- Analyse prédictive « ciblée »
Cette analyse permet de définir un coefficient dans l'équation de prédiction pour chacune des 24 protéines. La deuxième analyse permet de définir des coefficients dans l'équation de prédiction pour chacune des 24 protéines. Cette approche permet de définir un coefficient pour chaque protéine retenue dans l'équation de prédiction.
V Interactome
Les mesures WB (muscles LT et ST) et les scores TG (muscle LT) constituent les variables dépendantes à expliquer et les quantités des 24 protéines les variables quantitatives explicatives.
RESULTATS ET DISCUSSION
I Choix méthodologiques
Stratégie initiale
Développements techniques
- Mise au point de la révélation d’anticorps primaire par fluorescence
- Validations de la spécificité des anticorps primaires
To achieve this goal, we determined the inter- and intra-assay variability of Dot-Blot and the inter-assay variability of Western-Blot (Study 1), and the Dot-Blot response for proteins with different properties (Study 2) . Finally, we illustrate a Dot-Blot application for the large-scale validation of beef tenderness biomarkers (Study 4). Moreover, the intra-assay variability of Dot-Blot was 9%, similar to 10% of inter-assay variation.

Optimisation du protocole de Dot-Blot
Le Dot-Blot est donc une technique présentant un avantage significatif en termes de rapidité et de performance de l'analyse, tout en étant aussi fiable et reproductible que le Western-Blot, la technique de référence pour la quantification des protéines. Une autre caractéristique intéressante du Dot-Blot est son coût, bien inférieur à celui du Western-Blot en termes de produits et d’anticorps. Ainsi, pour réaliser des quantifications de protéines au cours de la thèse, cette technique Dot-Blot (Figure 38) a été retenue, ainsi que la détection de fluorescence.
II Résultats techniques sur l’utilisation du Dot-Blot
Ceci confirme que le Dot-Blot est une technique très fiable, plus que les techniques classiques de quantification de protéines (ELISA ou gels 2D, avec 25 à 30% d'erreur relative). Ainsi, les quantifications Dot-Blot coûtent environ 4 fois moins cher que le Western-Blot. C'est grâce à cette technique Dot-Blot que toutes les données brutes analysées dans cette thèse ont été obtenues (Tableaux 15 et 16).

IIIEffets muscle et type d’animal
Beef is an important economic asset in various countries such as USA, Australia and France (Veysset et al., 2005). Consumers express interest in high organoleptic quality of meat, especially tenderness (Geay et al., 2001). Many studies have shown that the abundance of some gene transcripts and/or proteins can be affected by the production system, animal type and muscle (Hocquette et al., 2007).
AUTHORS PROOF
For example, they showed that the LDH activity was higher in oxen than in bulls (Brandstetteret al., 2002). For example, Brandstetter et al. 1998) showed that MyHC-IIx was more expressed in oxen than in bulls. Brandstetteret al. (1998) study also showed that the amount of the MyHC-I was constant between oxen and bulls, which is consistent with the results of this study.
IV Relations entre abondance des protéines et la tendreté
Analyse par corrélations
- Muscle LT
- Muscle ST
- Muscles LT et ST cumulés
109 mesure WB), le ST des bouvillons (mesure WB), le ST des taurillons (mesure WB), le ST des bouvillons et taurillons groupés (mesure WB), et enfin sur toutes les données des muscles LT et ST groupés ( Mesure de la WB). Dans le TL du bœuf, la -calpaïne est significativement corrélée à la mesure WB (positive) et au score TG en sens inverse (négatif). Lorsque les deux types d’animaux sont combinés, de nombreuses protéines sont significativement corrélées à la mesure de WB.
Analyse descriptive







