Je tiens également à remercier MM. Jean-Paul Dren, Patrick Gentien et Guy Herrouin de m'avoir accueilli successivement à l'IFREMER à Nantes, au CREMA de l'Houmeau et à l'IFREMER à La Seyne-sur-mer. Merci également à l'équipe de l'Université de Toulon pour leur bonne volonté et leurs conseils : Nathalie, Stéphane et Cédric.
Généralités sur la bioaccumulation des métaux traces
Situation générale des métaux en Méditerranée
Le mercure
- Propriétés fondamentales: physiques, chimiques et biologiques
- Utilisations
- Cycle et sources naturelles et anthropiques
- Propriétés biologiques et toxicité
- Le mercure en méditerranée française
Le cadmium
- Propriétés fondamentales
- Utilisations
- Cycle et sources naturelles et anthropiques
- Propriétés biologiques et toxicité
- Le cadmium en méditerranée française
Le plomb
- Propriétés fondamentales
- Utilisations
- Cycle et sources naturelles et anthropiques
- Propriétés biologiques et toxicité
- Le plomb en méditerranée française
Le cuivre
- Propriétés fondamentales
- Utilisations
- Cycle et sources naturelles et anthropiques
- Propriétés biologiques et toxicité
- Le cuivre en méditerranée française
Le zinc
- Propriétés fondamentales
- Utilisations
- Cycle et sources naturelles et anthropiques
- Propriétés biologiques et toxicité
- Le zinc en méditerranée française
Réglementations, arrêtés
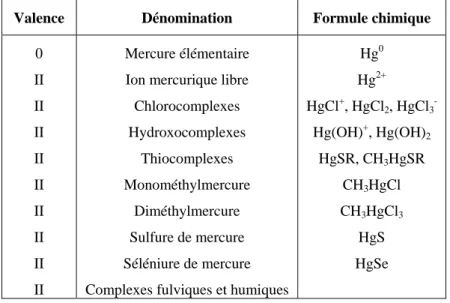
Définitions
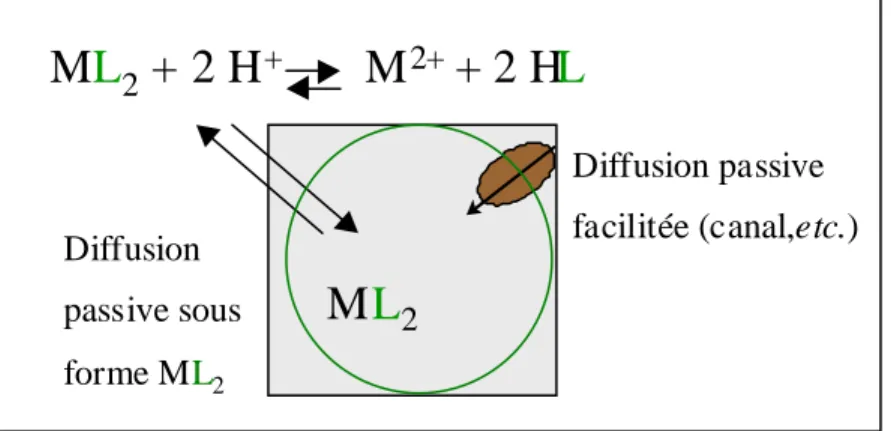
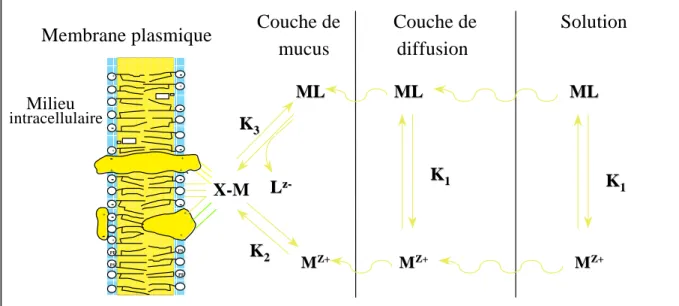
Mécanisme de capture des métaux
- Capture des métaux en solution
- Capture des métaux associés aux particules
Mécanisme d’excrétion des métaux
Mécanisme de stockage des métaux: organotropisme et amplification
- Organotropisme
- Les métallothionéines
- Bioaccumulation et biomplification dans les réseaux trophiques
Bioaccumulation du mercure chez la moule
Bioaccumulation du cadmium
Bioaccumulation du plomb
Bioaccumulation du cuivre et du zinc
Caractéristiques physico-chimiques du contaminant: spéciation et biodisponibilité
Facteurs biotiques
- Les processus de nutrition: action sur entrées/sortie
- Concentration métallique: quantité métallique et poids du bioindicateur
- Cycle de vie de l’organisme
- Composition biochimique et condition physiologique
Caractéristiques physico-chimiques du milieu environnant
- Solubilité dans l’eau, hydrophobicité
- Facteurs physico-chimiques
- Matière organique dissoute et particulaire
Interactions multi-factorielles
Implications pour les programmes de surveillance
Matériel et méthodes: stratégie expérimentale,
Planning expérimental: cinétiques de contamination et de décontamination
Choix des sites
Technique des transplants et mise en stabulation (« caging »)
Caractéristiques des échantillons
Caractéristiques des poches
Campagne de pose et de relèves
Protocole de prélèvement et allométrie
Protocole de traitement du matériel avant usage
Protocole de décoquillage, broyage et lyophilisation
Analyses de la concentration en métaux traces dans la moule
Caractéristiques physiques et physico-chimiques
Caractéristiques chimiques dissoutes
Caractéristiques chimiques particulaires
Analyses de la concentration en métaux dans le milieu: dissous et particulaire
- Performances de la méthode d’analyse des métaux dans la phase dissoute
- Performances de la méthode d’analyse des métaux dans la phase particulaire
Analyses de la concentration en métaux dissous « labiles »: DGT
Données physico-chimiques
- Baie du Lazaret
- Etang de Bages
- Ile de Port-Cros
- Comparaison inter-sites
Teneurs métalliques de chaque site
- Baie du Lazaret
- Etang de Bages
- Ile de Port-Cros
- Comparaison inter-sites
Teneurs métalliques dissoutes totales par la méthode des DGT: étude biodisponibilité
Baie du Lazaret
Etang de Bages
Ile de Port-Cros
Comparaison inter-sites
Cinétiques de contamination
- Cas du mercure
- Cas du cadmium
- Cas du plomb
- Cas du cuivre
- Cas du zinc
- Comparaison inter-métaux des cinétiques d’accumulation
Cinétiques de décontamination: site de Port-Cros
- Cas du mercure
- Cas du cadmium
- Cas du plomb
- Cas du zinc
- Cas du cuivre
Récapitulatif des cinétiques: bilan des observations de bioaccumulation
Interprétation des variations de croissance
- Variations spatiales et temporelles: importance de la quantité nutritive du milieu
- Variations temporelles: saisonnières: cycle de vie de l’organisme
- Relations allométriques et conditions physiologiques
Interprétation des variations de concentration et de quantité: essais de généralisation des
- Variations spatiales: importance de la contamination du milieu
- Variations temporelles: saisonnières: cycle de vie de l’organisme indicateur
- Variations spatio-temporelles: modifications environnementales
- Interactions des différents forçages, couplage et prédominance temporelle
Etat de pseudo-équilibre et facteur de bioconcentration (FBC)
Concept et hypothèses
Un exemple de modèle de bioaccumulation du cadmium: van Haren et al., 1994
Discussion
Généralités sur les modèles de
Généralités
Cette section tente d'expliquer la nature fondamentale du modèle à un compartiment et ses taux constants quelle que soit la situation, et explore ce qui se passe lorsque les hypothèses stables du milieu environnant ne sont pas respectées. Fondamentalement, l'analyse commence par l'identification des compartiments, qui peuvent être n'importe quelle entité traitée de manière homogène par rapport à une entité transportée vers ou depuis ce compartiment. Pour être plus explicite, l'accent est mis sur une problématique écotoxicologique où les compartiments peuvent faire partie de l'environnement (comme un lac ou un site expérimental), ou d'un organisme (comme un poisson ou un bivalve) ou d'une partie spécifique d'un organisme. (comme l'estomac ou le soma).
Le choix de la partition dépend non seulement du système décrit, mais surtout de l'échelle spatiale et/ou temporelle du problème étudié. Par exemple, pour décrire le transfert de polluants à travers une chaîne alimentaire, le poisson peut être décrit comme un seul compartiment. Lorsqu’il s’agit de la cinétique de pollution dans un organe spécifique, il est nécessaire d’ajouter davantage de partitions pour décrire le système.
Concept et formulation du modèle
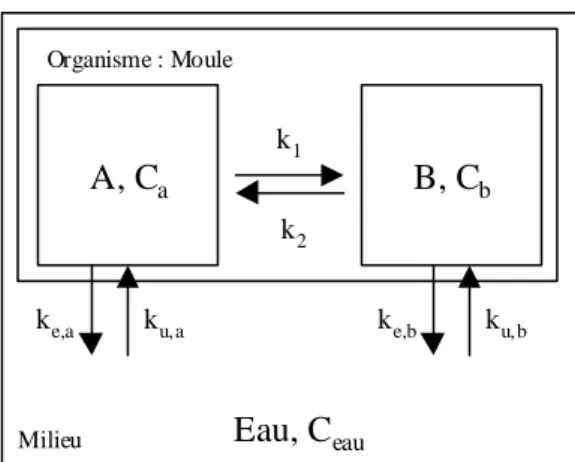
La moule est donc représentée par un compartiment unique où l'accumulation et les pertes sont liées à la concentration de contaminant présent dans le milieu, indépendant de l'état physiologique de cet organisme bio-indicateur (Figure 4.1). La concentration du contaminant dans l'organisme (Cmoule) résulte du bilan net de la capture du contaminant via l'eau (kuCeau) et de l'élimination de l'organisme (keCmoule). A l'entrée sous forme dissoute, peut être ajoutée une entrée du contaminant sous forme particulaire (kpCfood) (avec kp : taux d'entrée du contaminant via les particules, et Cfood : concentration du contaminant sur les particules).
Hypothèses d’application
Etat d’équilibre
Elle renseigne sur l'enrichissement en métaux de l'organisme par rapport au milieu environnant, mais ne permet pas de définir les conditions environnementales et physiologiques (Reinfelder et al., 1998). Le formalisme BCF n'est pas pratique lorsque les concentrations de la source sont inconnues et variables, ou lorsque les phases solides, les sédiments et les aliments contribuent de manière significative au captage.
Extension du concept: les modèles cinétiques à plusieurs compartiments
Le degré de complexité peut devenir très important pour, par exemple, suivre la dynamique des contaminants au sein de différents organes d'un individu afin d'expliquer leur organotropisme. La nature et l'étendue des processus de répartition pour chaque contaminant sont donc variables selon le compartiment et l'organe et sont déterminées par un nombre considérable de propriétés physico-chimiques correspondant à chaque sous-unité (Esser, 1986). Par exemple, la production de gamètes par l'organisme peut constituer une voie d'élimination supplémentaire pour les contaminants.
Les femelles perdront une partie de leur poids lors de la ponte, en fonction des efforts de reproduction de chaque espèce. Dans ce cas, la reproduction peut alors être assimilée à une dilution de la croissance : c'est une production de nouvelle biomasse. Si les polluants ne sont pas transférés, la reproduction affecte alors le poids de la femelle, concentrant le polluant restant.
Le problème de ce type de processus est que, pour de nombreuses espèces, la reproduction n’est pas continue. Par conséquent, l’effet de la reproduction sur la concentration du corps nécessite un lien avec un modèle de croissance et de développement de l’organisme.
Généralités
Concept et formulation du modèle
Extension possible à différents types de nourriture
- Les paramètres d'entrée: voies dissoute et particulaire
- Les efficacités d’assimilation
- Le paramètre de sortie: dissous et particulaire
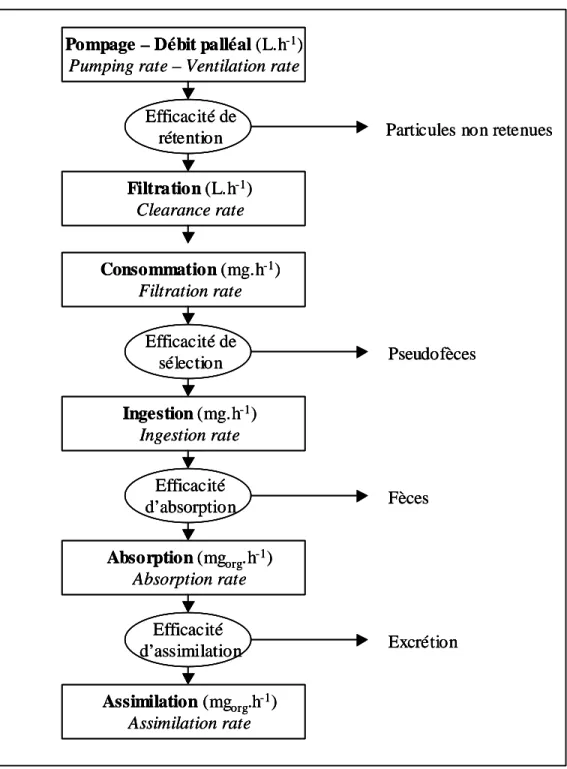
Ainsi, les processus nutritionnels interagissent avec les processus d'assimilation des polluants (Arai et al., 2002). Il existe donc de nombreux travaux sur la relation entre le débit de filtration et le poids corporel des bivalves marins. Certains auteurs ne font varier la filtration qu'en fonction de la taille ou du volume (V) de l'animal, indépendamment des conditions trophiques (Ross & Nisbet, 1990 ; van Haren & Kooijman, 1993).
D'autres considèrent la quantité totale de matière organique particulaire (Smaal & Sholten, 1989; Bayne et al., 1993). Elle est donc exprimée en fonction du débit de filtration, en fonction ou non du poids des tissus somatiques, du facteur de correction induit par la température et de la concentration en particules. D'autres expriment le taux de filtration en fonction de la concentration de particules organiques en carbone (Björk & Gilek, 1997) ou même de la concentration en particules organiques (Grant & Bacher, 1998).
Deux types d’efficacité d’assimilation des métaux se distinguent : l’une par voie dissoute et l’autre par voie particulaire. Cette efficacité d'assimilation des métaux correspond in fine au produit de l'efficacité d'assimilation de la matière organique et de l'efficacité de rétention des métaux sur cette matière absorbée.
Limites d’application et complexité croissante
En effet, l’assimilation des particules nutritives est le résultat de l’absorption des nutriments par l’épithélium gastrique selon les processus digestifs. Cela représente un processus physiologique de premier ordre et peut être comparé entre différents métaux et espèces dans différentes conditions. De nombreuses études ont permis d'estimer quantitativement la biodisponibilité des métaux issus de l'apport (Fisher et al., 1995 ; Reinfelder & Fisher, 1991 ; Luoma et al., 1992 ; Reinfelder et al., 1997 et 1998) ainsi que l'assimilation et la séparation. efficacité des différentes phases de digestion (Decho & Luoma, 1991 et 1994).
Importance du couplage
Théorie des modèles DEB
Le modèle de bioaccumulation de van Haren et al, 1994 est réalisé comme suit. Il existe de nombreux modèles DEB spécialement conçus pour la croissance des moules (tableaux 5.1 et 5.2). Variabilité de la population expliquée par les différences de seston et de dynamique alimentaire.
Testez l’utilisation d’un modèle statistique simple par rapport à un modèle mécaniste pour simuler la croissance des moules. Le maintien se fait au détriment de la croissance, démontrant une compétition naturelle des mêmes cellules somatiques pour l'utilisation de l'énergie. Les deux modèles prédisent une croissance à un niveau acceptable sur trois sites d'essai malgré la simplification de l'écophysiologie des moules.
Van Haren et Kooijman (1993) combinent un modèle pour représenter la croissance et la reproduction des moules en adaptant un modèle appliqué à d'autres espèces (figure 5.3) (van Haren et Kooijman, 1993). Une partie fixe de l'énergie (k) est allouée au maintien et à la croissance du soma ; le reste (1-k) est alloué au maintien de la maturation, au développement et à la propagation.

Application et validation sur d’autres sites: Généricité du modèle
A UTRES FORMES POSSIBLES DES EQUATIONS DU MODELE DE CROISSANCE
Modélisation de la croissance de la moule,
Les modèles de croissance
Cependant, les limites de l'exploitation des ressources conchylicoles sauvages nécessitent l'utilisation d'outils permettant d'améliorer la gestion de la conchyliculture et ainsi d'augmenter la production. Il existe deux types de modèles de croissance : les modèles empiriques simples dont le seul objectif est de décrire et prédire l'évolution temporelle du poids en considérant l'organisme comme un compartiment unique, de type von Bertalanffy ; et des modèles mécanistiques plus complexes, tels que les modèles DEB (Dynamic Energy Budget models), qui déterminent précisément la dynamique du bilan énergétique de la coquille, qui détermine l'état vital de l'organisme, comme la croissance, le développement et la reproduction. En effet, le terme dynamique fait référence au contraste avec les modèles statiques souvent utilisés, où les caractéristiques de l’individu n’évoluent pas explicitement au fil du temps.
Hypothèses générales des modèles DEB
On distingue trois compartiments principaux : la partie structurelle du corps, les réserves pour le stockage énergétique et la partie reproductrice. L'homéostasie est supposée pour chaque compartiment, c'est-à-dire la capacité à maintenir une composition chimique constante malgré les changements de composition dans l'environnement (Figure 5.1). Le bilan énergétique d'un organisme représente l'intégration de tous les processus liés aux gains et aux pertes d'énergie, ce qui permet de déterminer la quantité d'énergie disponible pour sa croissance et sa reproduction (Bayne et Newell, 1983).
Cette information physiologique, en plus d'être un indicateur du taux de croissance (Bayne et al., 1979 ; Riisgard et Randlov, 1981), est également considérée comme un bon indicateur de l'état physiologique de l'organisme (Widdows, 1985), en raison de sa une sensibilité élevée aux changements environnementaux et une précision élevée des taux (Grant, 1996).
Modèles de croissance individuelle
Les points spécifiques de ce modèle sont le détail des processus allant de la filtration à l'ingestion, en passant par l'absorption intégrant un modèle de nutrition optimale (Willows, 1992), la respiration et l'excrétion. Pour les sites intermédiaires où les niveaux de seston et les apports alimentaires sont faibles, les prévisions de croissance ne sont pas bonnes. 1986a) Concevoir un modèle général pour la nutrition, le stockage, la croissance et la reproduction des ectothermes, en fonction de la densité nutritionnelle à température constante.
La croissance de la coquille n'est pas prise en compte. analyse écophysiologique de la moule pour utilisation comme outil de gestion et identification des principaux processus manquants. Dans le modèle de Kooijman modifié, l'énergie assimilée par les moules va directement au compartiment de stockage et est ensuite répartie entre la reproduction et la croissance, l'entretien étant une dépense directe de croissance (Figure 5.2). La principale conclusion des modèles de Ross et Nisbet (1990) est que le seston et la dynamique alimentaire sont essentiels à la croissance et à la reproduction.
Modèle bioénergétique mécaniste utilisé pour prédire la croissance des moules, comprenant deux types d'alimentation (phytoplancton et matière organique particulaire détritique POM convertie en carbone organique particulaire POC) et une description complète des processus d'alimentation. L'entrée d'énergie, proportionnelle à la surface du corps, s'effectue lors de l'alimentation et va directement dans le compartiment de réserve, convertie avec une efficacité constante.

Simulations du modèle
On distingue ainsi différents flux énergétiques : le flux d'apport alimentaire (taux d'assimilation, taux d'utilisation), le flux de croissance, le flux de maintien et le flux de reproduction.
Relation volume/longueur: Shape
Effet de la température
Nutrition
Paramètres de maintenance
Paramètres intervenant dans la croissance et la reproduction
Récapitulatif des différents paramètres du modèle de croissance
Etude biologique et mathématique des différentes équations différentielles
Calcul du poids total de l’organisme
Définition des conditions initiales et des paramètres à estimer
Stratégie 1: fonction nutritive constante
Stratégie 2: fonction nutritive variable
Test des coefficients
Résultats du test sur la stratégie 1
Résultats du test sur la stratégie 2
Validité, pertinence des estimations et de la stratégie de simulation
Comparaison entre sites: interprétation du fonctionnement trophique, simulation long