Cependant, l'effet de la leptine (et de l'obésité) sur l'activité de la BO in vivo reste inconnu. Pour évaluer l’effet de la leptine et de l’obésité sur la plasticité olfactive, nous avons choisi le modèle murin ob/ob.
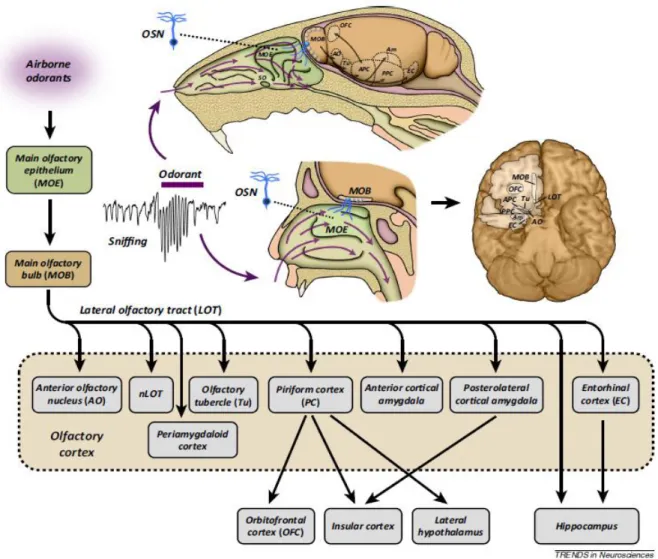
Organisation anatomique et fonctionnelle du système olfactif
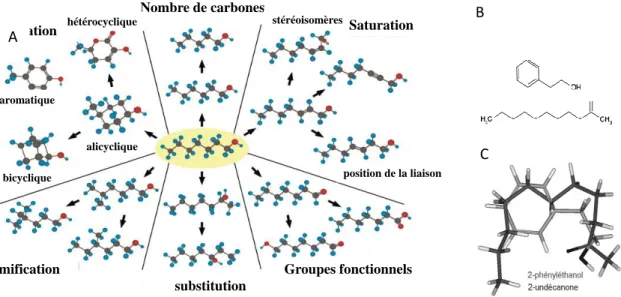
Le stimulus olfactif
Il s’agit d’une question difficile qui a fait l’objet de nombreux articles dans la littérature en raison de la nature chimique complexe, volatile et tridimensionnelle des molécules odorantes. La diversité des molécules odorantes indique la complexité du sens de l'odorat (modifié de (Johnson & Leon, 2007).
La muqueuse olfactive
- Anatomie et organisation cellulaire
- Fonctions des OSN et de leurs récepteurs
Le BM est un épithélium pseudostratifié recouvrant la partie postérieure de la cavité nasale, située au sein de l'os maxillaire. Cette signalisation calcique permet aux cellules encapsulées d'être impliquées dans le couplage neurovasculaire au niveau de la couche nerveuse olfactive (Lohr et al., 2011).
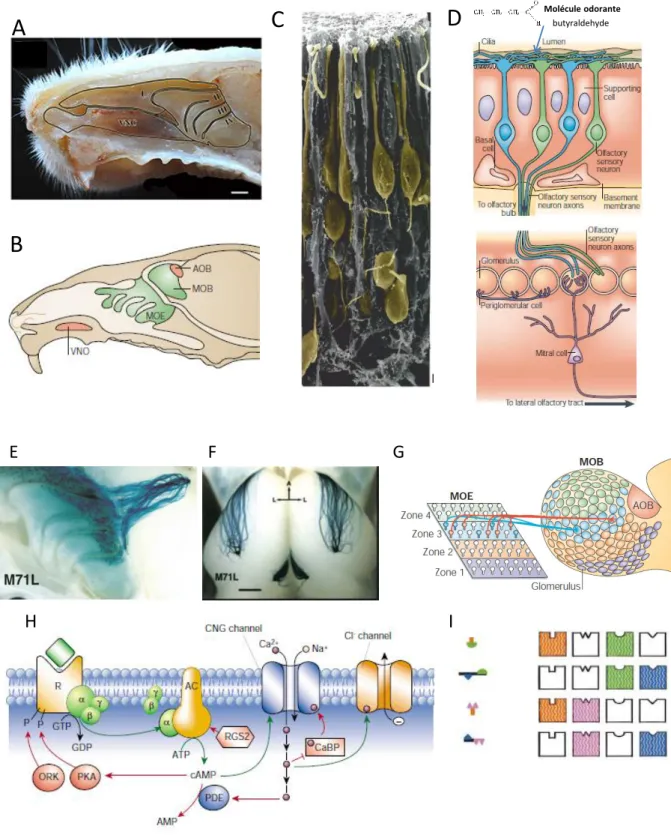
Le bulbe olfactif principal
- Anatomie et organisation cellulaire
- Fonctions du BO : un carrefour olfactif et métabolique
- Le codage temporel dans le BO
Ces projections sont largement étudiées dans le contexte de la mémoire olfactive précoce des rats nouveau-nés (Raineki et al., 2010). Cette technique a également été utilisée dans le cadre de l'étude de l'activation différentielle du BO par les voies rétronasale et orthonasale (Furudono et al., 2013).
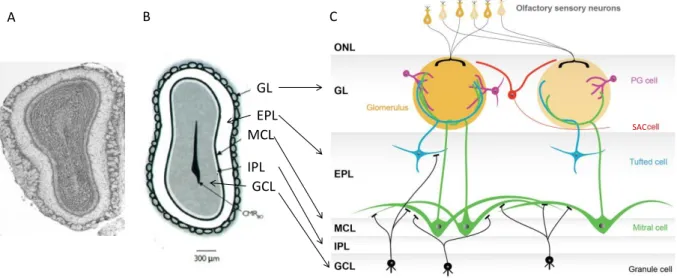
Sortie de l’information du BO vers le PC
Comme cela a été rapporté dans de nombreuses études, l'amplitude des sursauts gamma augmente proportionnellement à l'état d'attention de l'animal (Bressler, 1984 ; Eeckman et Freeman, 1990). Cette attitude d’attention et de motivation est facilement perceptible dès la phase initiale de toute formation. Ils ont conclu que les modèles spatio-temporels trouvés lors de l’analyse des sursauts gamma dépendaient davantage de l’importance de l’odeur pour l’animal que de sa nature chimique.
En fait, le codage des odeurs dans le BO est un processus très dynamique, mais aussi très plastique : la représentation spatio-temporelle des odeurs est fortement influencée par les paramètres respiratoires, par les propriétés intrinsèques des odeurs, mais aussi par la valeur et le sens associés devient cuir avec toutes les odeurs. La suite de l'introduction sera consacrée à la description des différents mécanismes qui contrôlent la prise alimentaire, la régulation du bilan énergétique et de l'obésité, et la manière dont cette dernière interagit avec le système olfactif et affecte la représentation spatio-temporelle des odeurs.
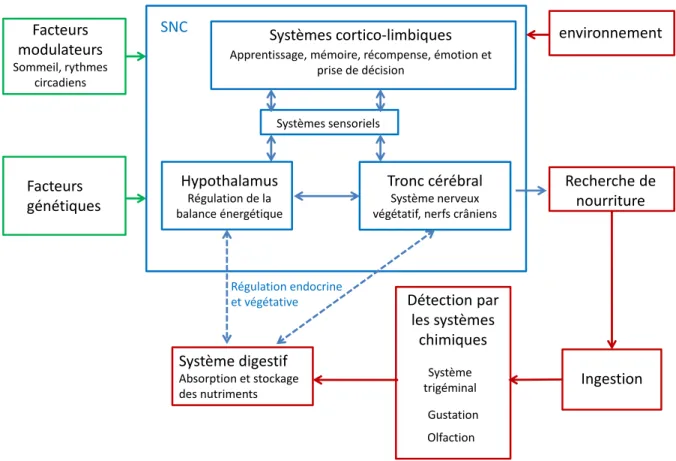
Métabolisme énergétique, prise alimentaire et obésité
Régulation centrale de la prise alimentaire
- L’hypothalamus
- Le tronc cérébral
- Autres structures du système limbique
Le cerveau joue un rôle très important dans la gestion des réserves énergétiques et dans la régulation de l’équilibre énergétique du corps. Le NTS est un noyau du tronc cérébral particulièrement impliqué dans la régulation de la prise alimentaire et du bilan énergétique. CCKA et CCKB par CCK et l'administration de cholécystokinine (CCK) directement dans le NTS ou dans le quatrième ventricule induisent une réduction de la prise alimentaire (Mercer et Beart, 2004).
Le système dopaminergique lié aux idées de plaisir participe au jugement de l'appétence des aliments et au contrôle de la prise alimentaire (Barbano et al., 2009). Cette modulation dépend du type de neurone, de l’appétence de l’aliment et de l’état.
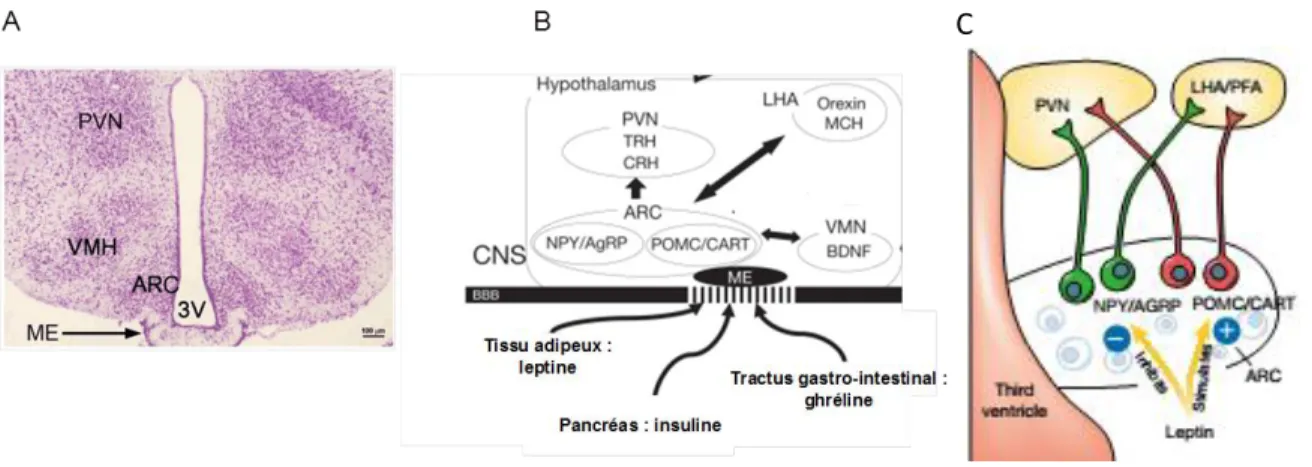
Signaux périphériques de régulation de la prise alimentaire
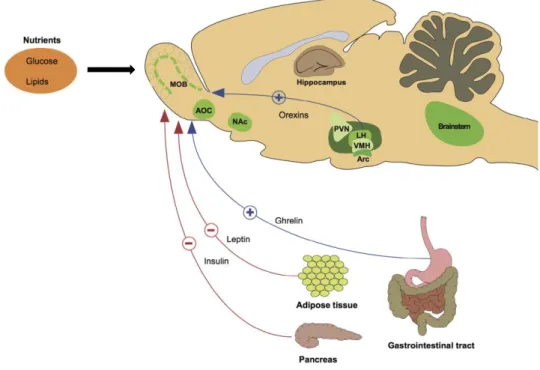
- Les nutriments
- Signalisation par le glucose et les acides gras
- Signalisation par les molécules orexigènes
- Signalisation par les molécules anorexigènes
Dès les années 1960, un effet du glucose sur les neurones hypothalamiques régulant la prise alimentaire a été décrit (Anand et al., 1964 ; Oomura et al., 1969) à partir des idées originales de Mayer (1953) et de sa théorie. "glucostatique" (Mayer, 1996) pour la régulation de la prise alimentaire. L'induction de la prise alimentaire peut être réalisée directement par une seule hormone, la ghréline. Il existe également une régulation centrale de la ghréline : une part importante de l'augmentation de la ghréline circulante dans la période préprandiale, la volonté est due à l'attente du repas par l'animal (Blum et al., 2009 ; Verhagen et al., 2011).
L'insuline est sécrétée par les cellules bêta des îlots de Langerhans du pancréas et est particulièrement impliquée dans la régulation métabolique périphérique et la régulation de la prise alimentaire ; il est très intéressant qu'elle s'exprime également au niveau BM (Lacroix et al., 2008). Il peut également traverser la barrière hémato-encéphalique et agir sur les récepteurs centraux du tronc cérébral et de l'hypothalamus pour provoquer une diminution de l'apport alimentaire (Woods et D'Alessio, 2008).
L’obésité
- Les modèles de l’obésité chez les rongeurs
- L’impact de l’obésité sur l’homéostasie énergétique
La CCK est donc un facteur de satiété : son administration exogène contribue à réduire la taille d'un repas une fois celui-ci commencé (Gutzwiller et al., 2004). Ce n’est que lorsque le produit du gène muté, la leptine, a été identifié par Zhang et al. 1994) qui ont initié des travaux sur les origines génétiques de l’obésité. L'inactivation spécifique du MC4 par une souris transgénique déficiente en MC4R provoque une hyperphagie et une obésité morbide (Huszar et al., 1997).
L'expression réduite ou la suppression des récepteurs de l'insuline dans le cerveau entraînent une hyperphagie et une obésité (Kuliczkowska-Plaksej et al., 2012). Il a également été démontré que chez les patients obèses, l'axe hypothalamo-hypophyso-surrénalien répondait plus efficacement à divers stimuli (Pasquali et al., 2006).
Interactions olfacto-alimentaires et problématique de la thèse
Systèmes orexigènes
Ils sont également présents en grand nombre sur les fibres centrifuges du BO (Soria-Gomez et al., 2014). En BO, l'injection locale d'un agoniste des récepteurs CB1 réduit l'hyperphagie après 24 heures de jeûne chez la souris (Soria-Gomez et al., 2014). Une hypothèse serait que CB1 médie les actions de la leptine, d'une manière similaire à l'hypothalamus (Cardinal et al., 2014).
Dans le BO, le NPY est peu localisé sur les corps cellulaires des neurones, mais également sur l'enveloppe du GL (Jia et Hegg, 2010). Les fibres NPY projetant vers le BO proviennent du locus coeruleus (Bouna et al., 1994) ou plus probablement de l'hypothalamus, mais il n'existe pas de données précises sur les connexions hypothalamo-bulbaires.
Systèmes anorexigènes
Ces récepteurs se trouvent également sur les cellules de soutien et les glandes de Bowman (Getchell et al., 2006). Enfin, certaines cellules de la muqueuse sécrétant l'OBP sont également sensibles à la leptine ( Badonnel et al., 2009 ). Dans le BO, la CCK est détectée dans toutes les couches neuronales sauf celle de l'ANY.
Les récepteurs CCK sont distribués dans le cortex olfactif, et CCK2R est particulièrement présent dans l'AON et le PC ( Honda et al., 1993 ). Dans BO, CCK2R est particulièrement présent dans l'IPL et dans les MC et les cellules juxtaglomérulaires ( Ma et al., 2013 ).
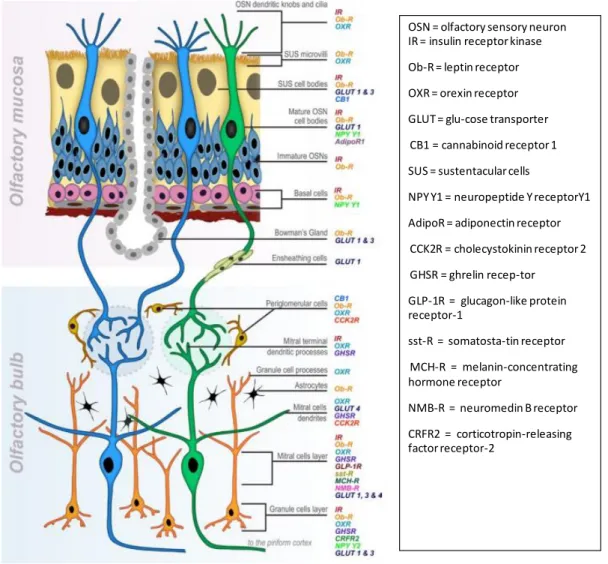
Interactions olfacto-alimentaires : études fonctionnelles
- Interactions entre état nutritionnel et perception olfactive : études comportementales
- Etudes moléculaires, cellulaires et fonctionnelles par électrophysiologie et imagerie
Ainsi, des travaux montrent que l'exploration des odeurs alimentaires augmente au cours d'un jeûne de 48 heures chez le rat (Prud'Homme et al., 2009). En ce sens, ils montrent qu'une injection de leptine réduit les performances olfactives de ces rats (Getchell et al., 2006). La réponse globale de la BM à une substance odorante mesurée par électro-olfactogramme est réduite après l'administration d'insuline (Lacroix et al., 2008).
Des études in vitro ont montré que l'activité de BO MT/C est modifiée par l'application d'orexine dans un bain de perfusion ( Hardy et al., 2005 ). Jeanne Pager (1972) a été la pionnière de l'étude du BO en examinant l'activité M/TC en fonction de l'état nutritionnel dans des conditions physiologiques de jeûne ou de satiété (Pager et al., 1972).
Nouveaux candidats pour la modulation de l’activité olfactive : GLP-1 et PK2
Enfin, la CCK est sécrétée par les cellules TC et excite les cellules MC, créant une boucle qui amplifie les projections de paires de glomérules activées par la stimulation d'un OR (Ma et al., 2013). Ces données suggèrent fortement qu’il existe un lien étroit entre la régulation du bilan énergétique et l’olfaction. En effet, le système olfactif n'est plus considéré uniquement comme un détecteur de signaux chimiques extérieurs au corps, mais aussi comme un détecteur parallèle de signaux chimiques internes liés au métabolisme énergétique (Palouzier-Paulignan et al., 2012b).
Problématique de thèse : étudier la plasticité spatio-temporelle des représentations sensorielles en BO chez des souris ob/ob-déficientes.
Problématique de la thèse : étude de la plasticité spatiotemporelle des représentations
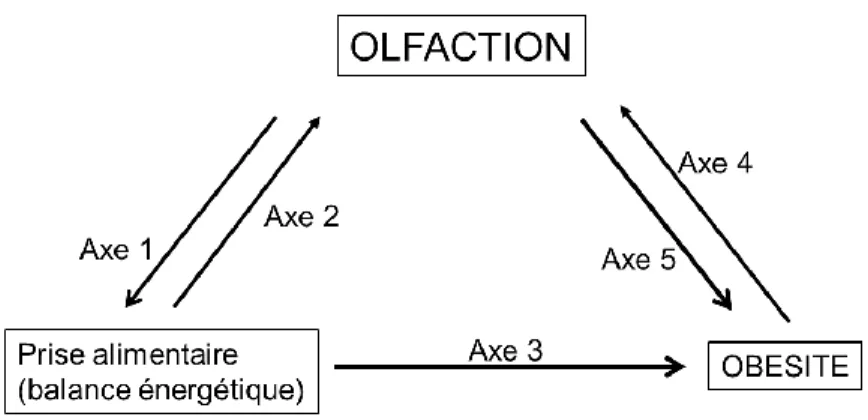
L'axe 1 est caractérisé par l'influence de l'olfaction sur la prise alimentaire et s'appuie historiquement sur les données de Le Magnen (2001) et plus récemment sur les données de Soria-Gomez et al. En contrepartie, l'axe 2 décrit l'influence des mécanismes métaboliques sur l'activité olfactive et s'appuie historiquement sur les données de Pager et al. 1972) démontrant la modification par l'état nutritionnel de l'activité électrophysiologique enregistrée dans le BO chez des rats éveillés. L'axe 3 implique des modèles d'obésité chez les rongeurs, en particulier les souris ob/ob et DIO, et des données chez les rongeurs et les humains suggérant qu'une augmentation de la consommation alimentaire est fortement impliquée dans l'obésité.
Dans ce domaine, l'équipe de Deborah Fadool a étudié la plasticité olfactive de l'OM dans un modèle DIO, avec une obésité induite plus tardivement par rapport à notre protocole (Thiebaud et al., 2014). Enfin, j’aborderai les données de la littérature susceptibles d’étayer cette interprétation (section « discussion ») en posant un regard critique sur les limites de nos approches ainsi que celles des données de la littérature pour suggérer des perspectives de recherche qui peuvent suivre sur mes travaux.
Méthodes expérimentales
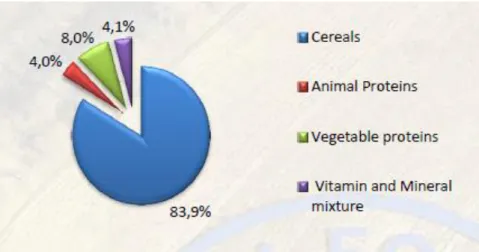
- Animaux, hébergement et alimentation
- Phénotype général des souris obèses
- Pesées
- Glycémie
- Locomotion
- Analyse statistique
- Odeur alimentaire et odeur neutre utilisées pour le MEMRI
- Fabrication de l’odeur alimentaire et choix de l’odeur neutre
- Tests comportementaux d’exploration olfactive spontanées (non opérants)
- Analyse statistique pour le comportement olfactif spontané
- Conditions expérimentales de l’imagerie MEMRI
- Injection de manganèse, stimulation olfactive et enregistrement IRM
- Protocole d’acquisition MEMRI
- Analyse manuelle des images MEMRI
- Analyse des signaux MEMRI par Statistical Parametric Mapping (SPM)
- Marquage anatomofonctionnel complémentaire du signal MEMRI
- Injection de leptine
- Conditions expérimentales de l’Imagerie Optique du Signal Intrinsèque
- Anesthésie des souris témoins et ob/ob
- Chirurgie
- Odeurs
- Stimulation olfactive
- Acquisition des images
- Microscope
- Source lumineuse
- Camera
- Logiciel d’acquisition
- Electrophysiologie chez la souris éveillée
- Implantation des électrodes d’enregistrement à demeure
- Dispositif d’électrophysiologie
- Analyse des signaux LFP
- Test comportemental de discrimination olfactive (Go / No Go)
- Neurogenèse
- Injections des marqueurs de la division cellulaire
- Analyse des tissus
- Protocole d’analyse et de comptage des cellules
- Quantification des marqueurs moléculaires pour estimer l’inflammation intrabulbaire chez les
Images d'activités de recherche spontanées provenant de deux caméras, l'une tournée vers la gauche et l'autre tournée vers la droite. La somme de l'intensité des pixels activés et de leur position ont été enregistrées et comparées entre chaque groupe. L'étude et l'enregistrement de l'activité neuronale nécessitent l'implantation chronique d'électrodes d'enregistrement LFP dans le BO.
Le connecteur de l'électrode implantée a été fixé à la tête de la souris à l'aide de ciment dentaire. A l’inverse, après la présentation de l’odeur non renforcée (15% de butyraldéhyde), l’animal ralentit ou s’arrête, augmentant la latence de réponse.
Résultats
- Phénotype et physiologie des souris obèses
- Souris déficientes en leptine depuis la naissance, souris ob/ob
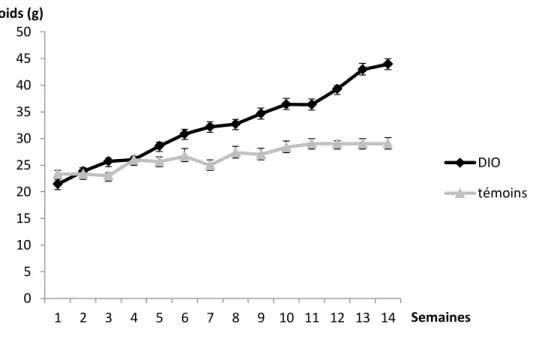
- Physiologie des souris rendues obèses par alimentation hyperacalorique -hyperlipidique
- Activité locomotrice
- Imagerie Optique des Signaux Intrinsèques
- Tests comportementaux d’exploration olfactive spontanée
- Exploration de la pâte odorisée versus la pâte non odorisée
- Exploration de l’odeur d’amande, de l’odeur d’orange et de l’odeur de pentylacétate
- IRM renforcée au contraste de manganèse
- Analyse manuelle de l’intensité brute des pixels
- Comparaison d’images MEMRI analysées par SPM
- Neurogenèse
- RtPCR
- Test de discrimination olfactive et enregistrement des réponses électrophysiologiques en LFP
- Courbes d’apprentissage illustrant la progression des animaux au cours de la tâche Go/NoGo
- Analyse des réponses oscillatoires enregistrées au cours de la tâche de discrimination
- Résumé des résultats
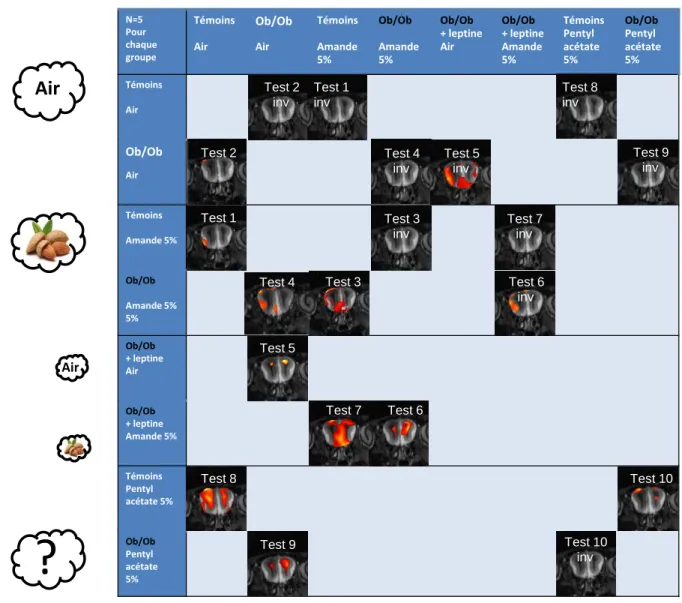
Comparaison intergroupe de l'activité entre des souris ob/ob stimulées avec 5 % d'amande et des souris témoins stimulées avec 5 % d'amande. Ce test compare l'activité de souris ob/ob stimulées avec 5% d'odeur d'amande à celle de souris ob/ob stimulées par de l'air. Réalisé dans l'autre sens, le test n'indique pas de zone d'activité plus élevée chez les animaux témoins stimulés avec 5% d'amande (test inverse 7).
Réalisé dans l'autre sens, le test ne montre pas de zone d'activité plus élevée chez les animaux témoins stimulés par l'air (test inverse 8). Réalisé dans l'autre sens, le test ne montre pas de zone d'activité plus élevée chez les animaux témoins stimulés par l'air (test 9 inv). Comparaison intergroupe de l'activité entre des souris témoins stimulées avec 5 % d'acétate de pentyle et des souris ob/ob stimulées avec 5 % d'acétate de pentyle.
Zone dorsale En Ob/ob, l'activation semble dépendre de l'odeur, mais pas de la leptine.
Discussion
- Choix des modèles et des techniques d’étude des interactions olfacto-alimentaires
- Modèles murins
- Choix de l’anesthésie et de la technique d’imagerie neurofonctionnelle
- Injection de la leptine
- Variations du comportement d’exploration en fonction de l’état métabolique, de l’état
- Implication de la leptine dans la distribution spatiale de l’activité dans le BO
- Plasticité de la représentation temporelle des odeurs chez les souris obèses suite à un
- Leptine, obésité et régulation de mécanismes olfactifs
Fait intéressant, chez le rat, un antagoniste de la leptine provoque une réduction de l'activité locomotrice volontaire dans une roue d'exercice (Matheny et al., 2009). Ces données suggèrent que l’action de la leptine n’est pas spécifique à la présentation des odeurs alimentaires en ob/ob. L'action de la leptine n'est donc généralement pas inhibitrice comme indiqué en électrophysiologie BM (Savigne et al., 2009).
L'inhibition du retour centrifuge, qui supprime les oscillations bêta, provoque également une augmentation de la puissance des oscillations gamma (Martin et al., 2006). Cette étude cellulaire réalisée en ob/ob pourrait également expliquer pourquoi on ne retrouve pas chez les souris ob/ob l'effet inhibiteur de la leptine sur l'activité des couches profondes induite par l'odeur alimentaire visualisée chez les rats à jeun (Prud'homme et al. , 2009).