THÈME : Etude de la croissance de Chlorella vulgaris en photobioréacteurs discontinus et continus, en présence de fortes concentrations de CO2. Calcul de la productivité et de la biofixation de Chlorella vulgaris en cours de culture.
Etude bibliographique
Matériels et méthodes
C. Modélisation de la croissance de Chlorella vulgaris
Absorption de la lumière dans le réacteur en fonction de la concentration cellulaire en C. Modélisation de la fraction massique d'azote dans la cellule y, après trois jours pour C. vulgaris cultivé avec une intensité lumineuse de 180 µmol.m-2.s - 1 p.106 Figure III.10. Comparaison entre les résultats obtenus par l'expérience et le modèle pour a) l'évolution de la concentration cellulaire, b) l'évolution de la fraction azotée intracellulaire et c) l'évolution de la concentration en azote dans le liquide pour C. vulgaris.
Perspectives de développement industriel
Annexes
Etude de l’influence du taux de dilution et de la
Introduction générale
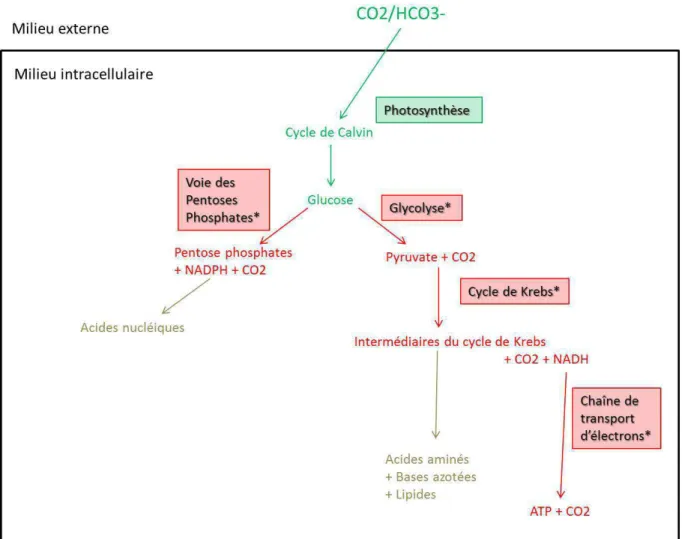
Selon certains auteurs, la production de 100 tonnes de biomasse algale permettrait de capter 180 tonnes de CO2 (Chisti, 2007). L'objectif de cette thèse est d'étudier la faisabilité d'un procédé de captage du CO2 grâce à la culture de la microalgue Chlorella vulgaris.
Etude bibliographique
A Le système carboné
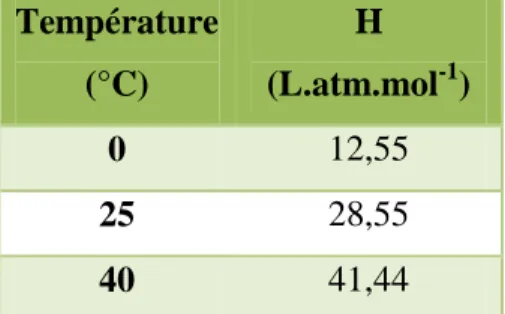
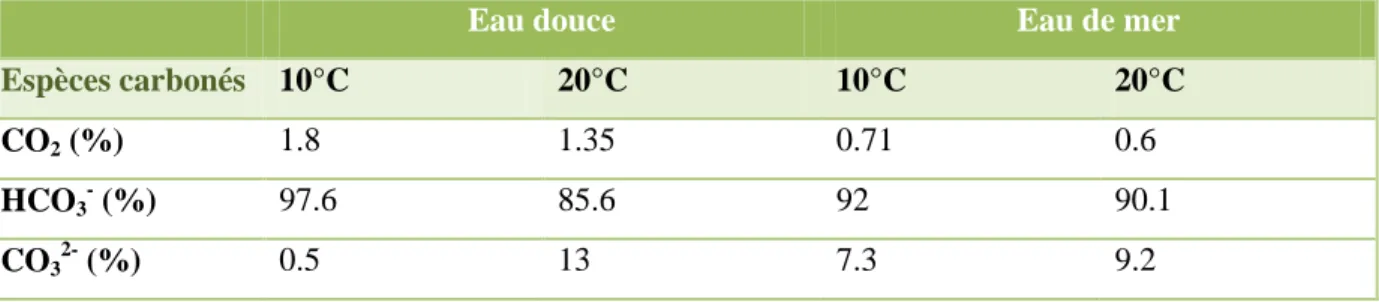
La dissolution du CO2 dans l'eau dépend de la pression partielle du CO2 dans le gaz et de la température. La constante de Henry dépend de la température (tableau I.1) : elle diminue avec la température.
B Présentation du modèle d’étude
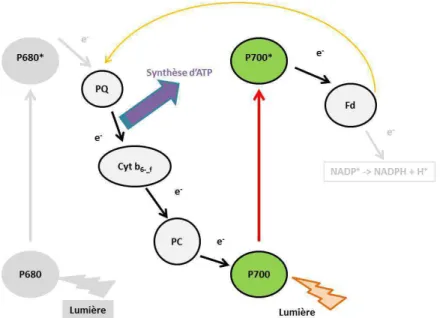
Le transport est alors dit acyclique et implique une dissociation de l'eau et la synthèse de NADPH et d'ATP. Cela est dû en partie à la capacité de la cellule à accumuler du CO2 (Falkowski et Raven, 2007).
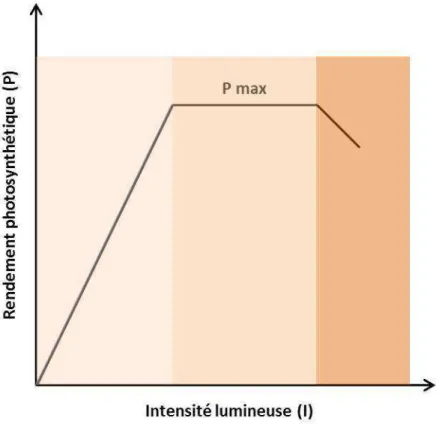
C La culture en photobioréacteur : le cas de la colonne à bulle
Le but de la culture continue est de maintenir stable la concentration cellulaire dans le réacteur. L'assimilation de cet élément limitant par la cellule est cependant dépendante de la concentration de l'élément limitant dans le milieu de culture (Droop, 1973).
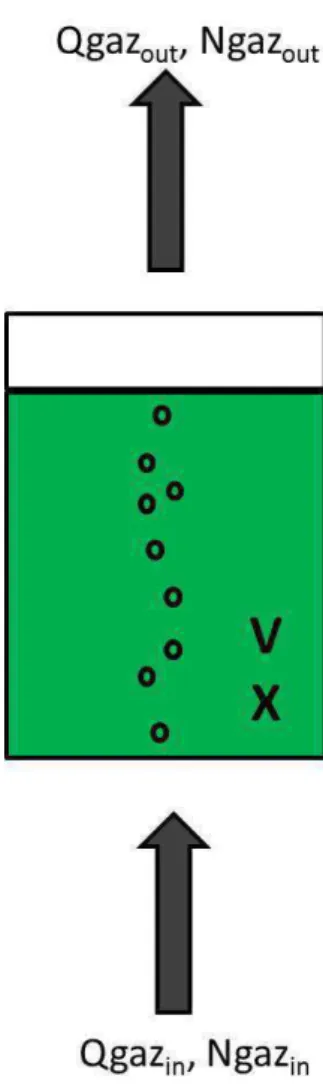
D L’impact des paramètres de culture
Il convient de souligner que la loi des quotas cellulaires reste l'approche privilégiée dans le domaine de l'étude de la physiologie (Bougaran et al., 2010). Contrairement à l'azote, une limitation du phosphore dans l'environnement entraîne une réduction de l'activité du CCM (Beardall et al., 1998). En revanche, la diminution de l'azote dans l'environnement entraîne une diminution de la capacité photosynthétique de la cellule (Yuong et Beardall, 2004 ; Richardson, 1969, Osborne et Geider, 1986).
55 une diminution de l'assimilation du CO2, de la capacité photosynthétique, de la synthèse de chlorophylle et une augmentation des xanthophylles (Huner et al., 1998). Chez Spirulina platensis, la température affecte l'ultrastructure, la morphologie et l'activité photosynthétique de la cellule (Gao et al., 2008).
Matériels et méthodes
B. Analyses
L'échantillon est d'abord soniqué pendant 10 minutes pour éliminer les agrégats, puis dilué avec de l'eau distillée avant d'être compté avec un granulomètre. Les analyses de carbone organique particulaire (POC) et d'azote organique particulaire (PON) sont effectuées à l'aide d'un analyseur élémentaire CHNS/O série FLASH 2000 (Thermo Scientific). Pour cette mesure, on utilise le surnageant des échantillons, à partir duquel est effectuée la mesure du carbone et de l'azote intracellulaires.
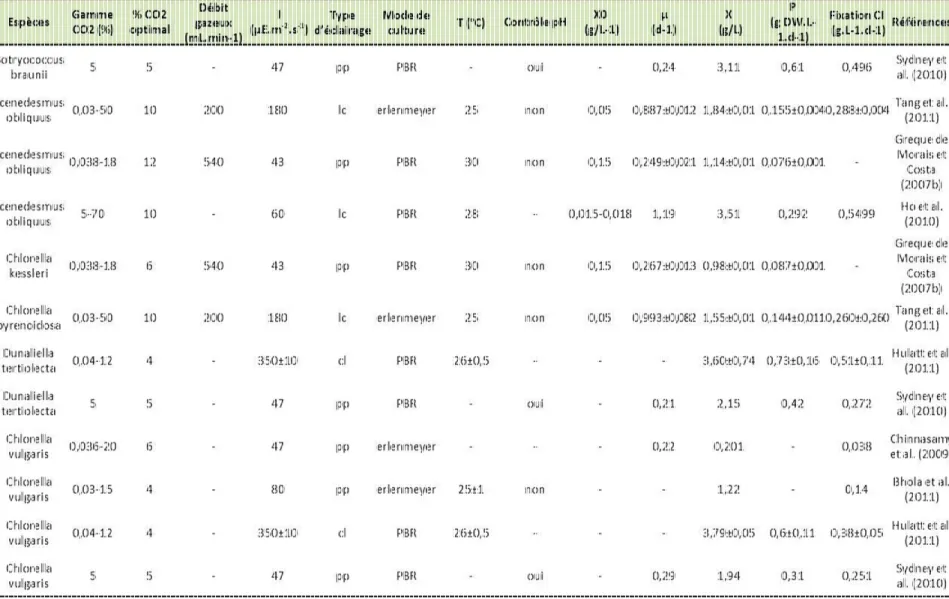
Les échantillons sont dilués avec de l'eau ultra pure pour correspondre à la plage de concentration de l'instrument. Enquête sur l'influence de la concentration de CO 2 et de l'intensité lumineuse sur la croissance de.
- Abstract
- Introduction
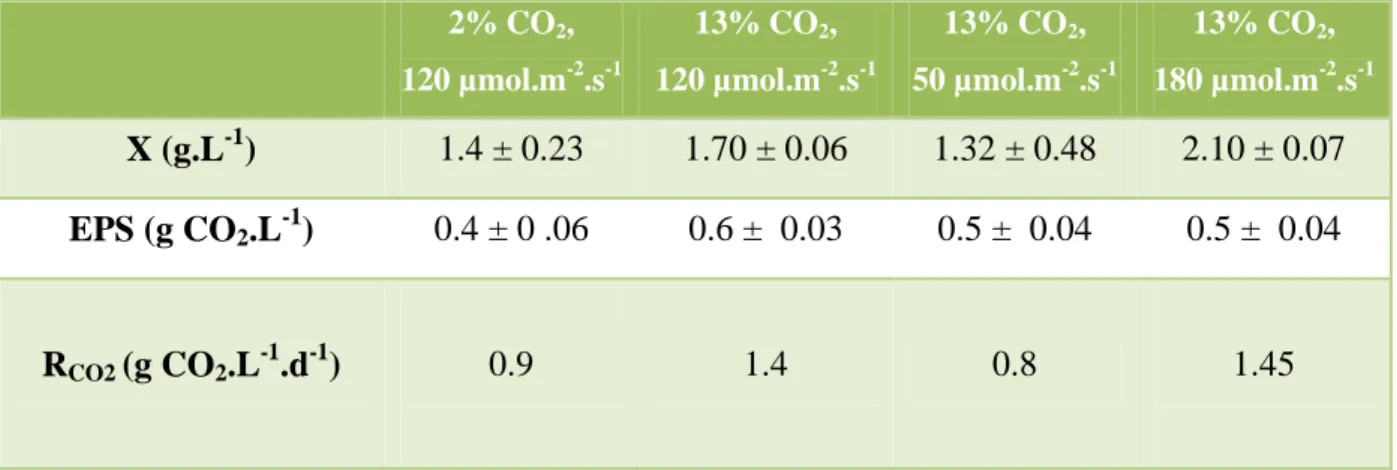
Nitrogen deficiency also seemed to affect growth. The algae showed different physiological adaptations under the different culture conditions. A sample of the culture (10 ml) was removed from the reactor and centrifuged (20 minutes; 4000 rpm) and the pellet was washed with deionized water (10 ml). With a low supply of CO2. 2% v/v), the non-exponential growth phase ended on day 6, leading to the stationary phase.
For a cellular concentration of approximately 2×1010 cell.L-1, the entire incident light was absorbed in the reactor, regardless of the culture conditions. The decrease in chlorophyll-a content and the increase in cellular C:N ratio on days 5 and 6 for the cultures grown at a light intensity of 120 µmol.m-2.s-1 and 180 µmol.m-2 .s-1 .
D. Conclusion
114 Pour les cas présentés, à l'exception de la figure III.12.a, la modélisation est en accord avec les valeurs expérimentales. Les cellules qui consomment du CO2 ont l'avantage car, bien que la limitation des nitrates puisse affecter l'activité de Rubisco, l'assimilation du carbone inorganique sous forme de CO2 nécessite moins d'énergie, permettant à la cellule d'allouer préférentiellement ces molécules d'énergie à d'autres métabolismes, maintenant ainsi l'activité cellulaire. Pour modéliser la croissance des algues dans ces conditions, nous avons recherché la valeur permettant le meilleur accord entre le modèle et les valeurs expérimentales.
Il apparaît que pour valider le modèle dans ces conditions, le terme de Droop doit être égal à 1, et donc la croissance de C. Etude de l'influence du taux de dilution et de la concentration en nitrates sur la croissance et la
A. Matériel et méthodes
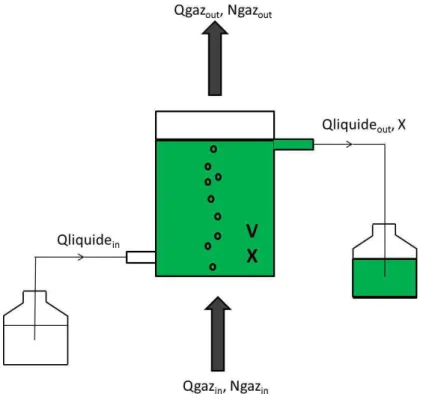
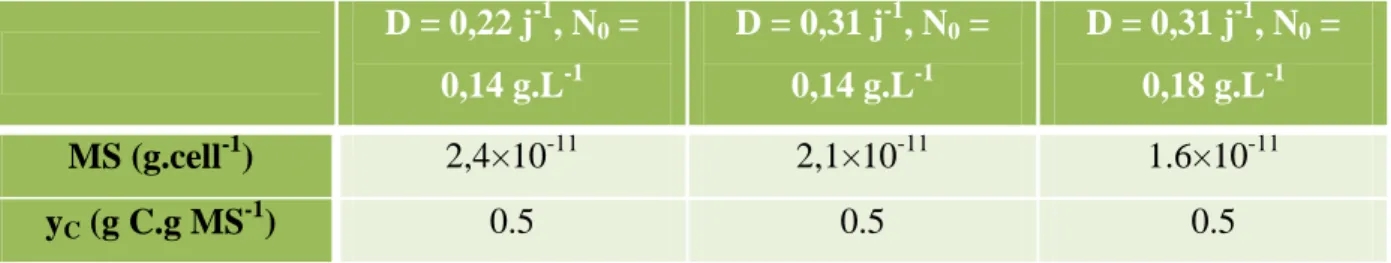
Il a été démontré dans le chapitre précédent que le réacteur se comportait comme un réacteur parfaitement agité (III.A). De plus, une fois l'état d'équilibre atteint, des échantillons en double sont prélevés pour mesurer la fraction intracellulaire de carbone (yC) et d'azote (yN), la concentration d'exopolysaccharides (EPS), la concentration de carbone organique dissous (DOC), la matière sèche et la concentration de nitrates dans le réacteur. La concentration en nitrates dans le milieu de culture est mesurée lors du changement du flacon de milieu.
Cette mesure est donc obtenue différemment de la p-value présentée dans le chapitre précédent ; pour éviter toute confusion, on l'appellera MS dans l'étude présentée ici. Bilan carbone dans le réacteur à débit constant de CO2 perdu par le gaz (mol/h) :. IV.8) Flux de CO2 inclus dans le carbone intracellulaire (mol/h).
B. Résultats
Dans cette étude, la concentration en exopolysaccharides dans le milieu de culture (EPS) a également été mesurée. Effet de la concentration de nitrates sur la biomasse et l'efficacité de fixation du CO2 chez Chlorella vulgaris. Un bilan carbone a été réalisé à partir des mesures du CO2 dans le gaz, de la concentration en DOC et POC.
Valeurs optimisées de la constante de demi-saturation pour la lumière (KE), de la constante de vitesse maximale pour la consommation de nitrates (fS) et de la constante de demi-saturation pour les nitrates (KN). Comparaison entre les valeurs expérimentales et les valeurs déterminées par le modèle pour la concentration en nitrate (NO3-.-N) dans le réacteur et le quota d'azote intracellulaire (y) à l'état d'équilibre pendant la culture continue de Chlorella vulgaris en fonction des différentes conditions de croissance (A, B et C).
![Tableau IV.2. Valeurs de la concentration de biomasse (X), de la productivité (R X ), de la biofixation du CO 2 (R CO2 ), du quota de carbone interne (POC), de la concentration en carbone organique dissous ([DOC]), de la concentration](https://thumb-eu.123doks.com/thumbv2/1bibliocom/463466.69230/147.892.92.812.646.1032/tableau-valeurs-concentration-productivité-biofixation-concentration-organique-concentration.webp)
C. Discussion
Ce phénomène aurait pu intensifier la limitation de la lumière, réduisant encore davantage la pénétration de la lumière dans la culture et augmentant ainsi la zone sombre dans le réacteur. En effet, la biomasse est une mesure de matière sèche dans le réacteur, sa diminution peut donc s'expliquer soit par une diminution de la concentration cellulaire, soit par une diminution de la matière sèche par cellule. Collos et ses collègues (2005) rapportent que certaines espèces de phytoplancton sont capables d'adapter leur capacité d'assimilation de l'azote en fonction de sa concentration dans le milieu de culture.
Smith et al (2009) rapportent également ce phénomène de modification du KN en fonction de la concentration en nitrate dans le milieu et le relient au nombre de sites de fixation présents sur la cellule, qui varie en fonction de la concentration externe en nitrate. L’augmentation de la valeur KN pourrait donc être une adaptation des algues à une faible quantité de lumière disponible.
D. Conclusion
Cependant, lors de l’étude en série, nous avons vu que le modèle était capable de prédire l’évolution de différents paramètres lors d’une double limitation lumière et nitrates, ce qui n’est plus le cas en continu. De plus, à mesure que le taux de dilution augmente, le modèle représente correctement la concentration cellulaire (Figure IV.3 B et C) lorsque celle-ci est prise en compte. Le modèle Droop a été utilisé pour simuler la croissance des algues dans le réacteur.
140 Le modèle défini lors de l'étude batch n'a pu être appliqué que dans le cas d'un taux de dilution de 0,22 j-1. Lorsque le taux de dilution a été augmenté, il a fallu ajuster le modèle et tenir compte du fait que la croissance cellulaire n’est limitée que par la lumière.
Perspectives de développement industriel
B. Résultats
Variation de la concentration cellulaire en sortie du réacteur (Xf), du débit liquide (Ql) et du volume du réacteur (V) pour différents taux de dilution (D). Dans un réacteur continu à volume fixe, l’augmentation de l’intensité lumineuse entraînera une augmentation de la concentration cellulaire. Pour réduire significativement le volume (V), il faut accepter une concentration plus élevée de cellules dans le réacteur.
L'augmentation de deux des paramètres, l'intensité lumineuse et la concentration cellulaire, permet de réduire beaucoup plus efficacement le volume du réacteur (tableau V.5). Lorsque les valeurs d'intensité lumineuse et de concentration cellulaire doublent, le volume du réacteur est réduit de moitié.

C. Discussion
Au cours de l'étude de groupe, il a été démontré que la quantité d'EPS excrétée variait en fonction de la concentration de CO2 dans le gaz d'alimentation. Par conséquent, une concentration cellulaire élevée, comme prévu dans un processus à l’échelle industrielle, peut entraîner la production d’une grande quantité d’EPS. Augmenter l'intensité lumineuse ou la concentration cellulaire permettrait de diminuer le volume du réacteur.
161 Plus la lumière augmente, plus le volume du réacteur diminue pour une même concentration de cellules. Il est possible de réduire plus efficacement le volume requis du réacteur en augmentant l’intensité lumineuse et la concentration cellulaire dans des proportions égales.
D. Conclusion
Cependant, une intensité lumineuse trop élevée pourrait provoquer une photoinhibition cellulaire et ainsi altérer le processus. Des études sont menées pour optimiser la géométrie des photobioréacteurs afin que des intensités lumineuses élevées soient utilisées sans apparition de photoinhibition ou que les cultures à densité cellulaire élevée ne souffrent pas de dominance lumineuse. prononcée (Moline grima et al., 1998 ; Cornet, 2010). Cette stratégie pourrait également empêcher les dommages cellulaires dus à une trop grande quantité de lumière reçue.
Cependant, comme mentionné précédemment, avant de pouvoir envisager une concentration cellulaire élevée dans le réacteur, il faut anticiper une éventuelle limitation par le carbone inorganique dissous. 162 déjà qu'en se mettant dans des conditions optimales de croissance, de nombreuses questions restent encore sans réponse quant à la mise en œuvre du procédé de captage du CO2 à l'échelle industrielle, notamment quant à la capacité d'utilisation industrielle d'un tel procédé.
Conclusions et Perspectives
L’augmentation de l’intensité de la lumière incidente a entraîné une augmentation de la consommation de biomasse et de CO2. L'étude ne permet pas de déterminer si ces effets sont directement liés à la concentration de CO2 dissous dans le milieu ou indirectement à la valeur du pH résultant de la concentration de CO2 dissous. Des tests en continu avec un gaz à 13% de CO2 (proche des concentrations industrielles) ont permis de mesurer les effets du taux de dilution et de la concentration en nitrates dans le milieu de culture sur la concentration cellulaire et la biofixation du CO2 par C.
La modification du taux de dilution (D) a entraîné une légère diminution de la concentration de cellules et de biomasse, ainsi qu'une augmentation de la biofixation du CO2. Par ailleurs, il serait intéressant de poursuivre les travaux menés sur la modélisation de la croissance de C.