A variedade de hábitos encontrados nas Leguminosae é grande, pois existem árvores emergentes de grande porte, ervas voláteis, arbustos, subarbustos e cipós (LEWIS et al., 2005; LPWG, 2017). Suas espécies podem ser de hábito herbáceo, arbustivo, lenhoso ou lianescente, e embora ocorram predominantemente em regiões temperadas, existem muitos gêneros que estão restritos aos trópicos (LEWIS et al. 2005; LPWG, 2017).

Anatomia da madeira na identificação taxonômica das
Identificação molecular de madeira
Diferentes regiões do genoma nuclear ou citoplasmático (mitocondrial ou plastidial) de organismos vegetais podem ser acessadas por meio de marcadores moleculares específicos ou universais (KALIA et al., 2011). O genoma do cloroplasto (cpDNA) é uma molécula circular haplóide e é predominantemente herdado pela mãe na maioria das angiospermas (ENNOS et al., 1999, PROVAN et al., 1996).
Objetivo geral
Objetivos específicos
Explorar a capacidade dos marcadores microssatélites do genoma do cloroplasto (cpSSRs) para diferenciar e identificar geneticamente nove espécies da família Leguminosae; Comparar dados anatômicos e genéticos em análises de variação interespecífica em espécies arbóreas da família Leguminosae na Mata Atlântica.
Obtenção do material botânico
A partir de Xilotecas
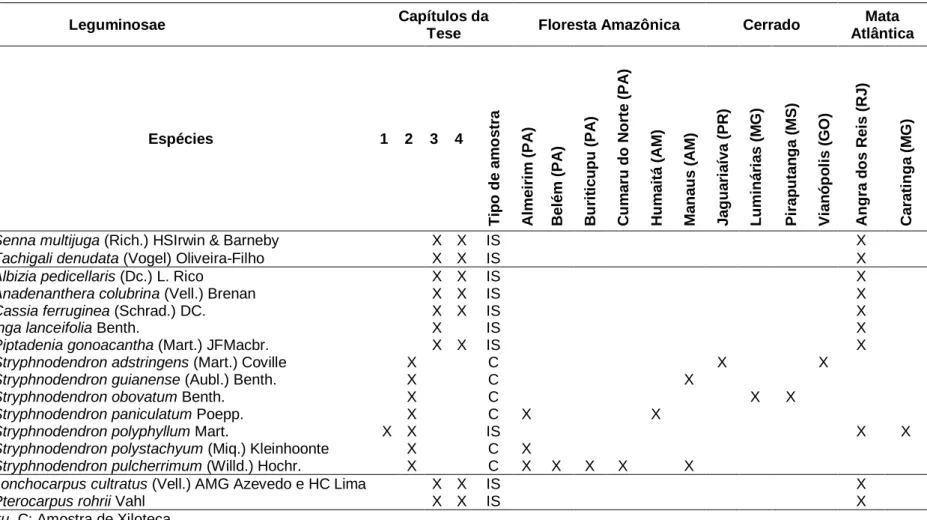
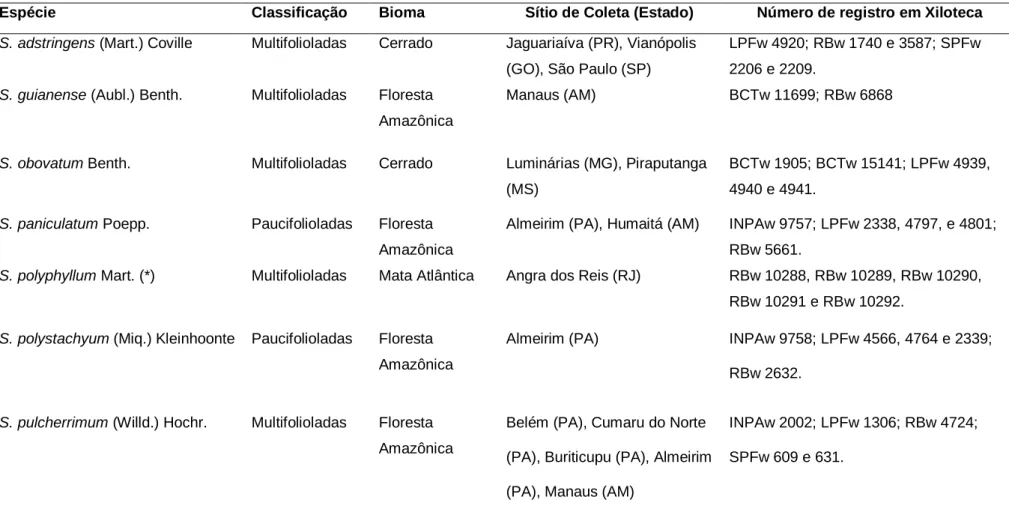
Amostras de madeira de Stryphnodendron astringens, Stryphnodendron guianense, Stryphnodendron obovatum, Stryphnodendron paniculatum, Stryphnodendron polystachyum e Stryphnodendron pulcherrimum, foram obtidas de Xylothecas brasileiras registradas no Index Xylariorum (https://www.kew.org/files/files/index Xylariorum. Harry). van der Slooten, Biblioteca de Madeira do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro e Biblioteca de Madeira Nanuza Luiza de Menezes.
A partir de remanescentes de Mata Atlântica
- Reserva Particular do Patrimônio Natural Feliciano Miguel
- Parque Estadual da Ilha Grande
As amostras de madeira estão registradas e depositadas na Xiloteca do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro (RBw) e na Xiloteca Dra. Legenda: Não.: Número de registro no Herbário da Universidade do Estado do Rio de Janeiro (HRJ); DAP.: diâmetro a 1,30 m do solo em centímetros;

Anatomia da madeira
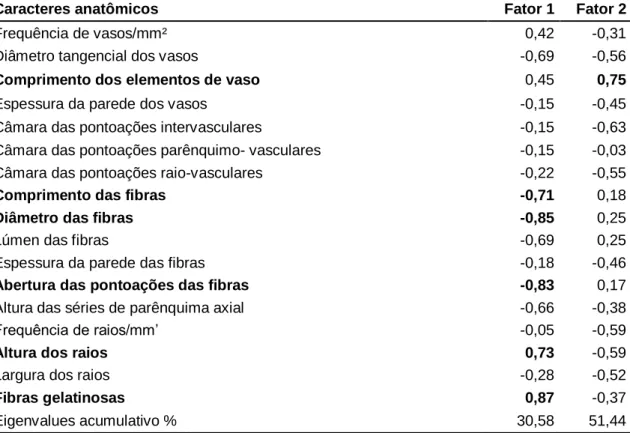
As características anatômicas foram submetidas ao teste de Shapiro-Wilk para avaliar a normalidade dos dados. Quando não foi observada distribuição normal, a não normalidade dos dados foi obtida por transformação logarítmica. As características anatômicas foram submetidas à análise de componentes principais (PCA) para avaliar o valor diagnóstico da anatomia das árvores na separação de espécies ou grupos de espécimes utilizando o programa Statistica versão 12.
Extração de DNA
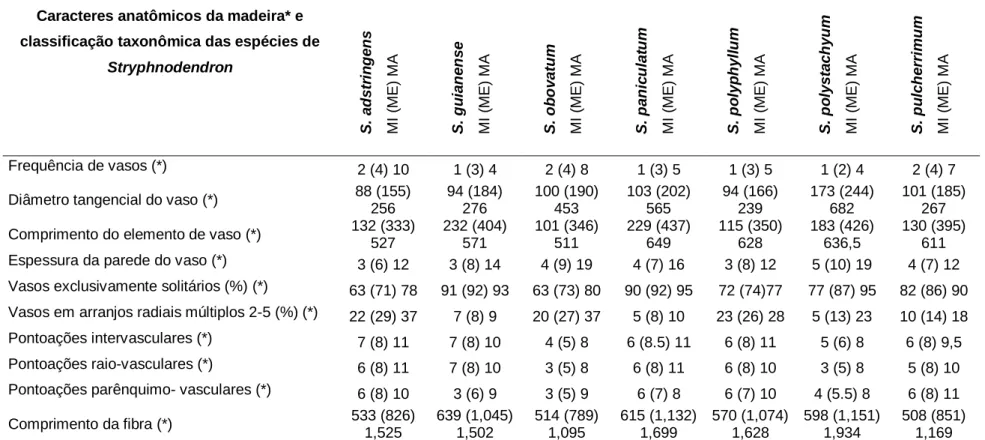
As medidas e contagens dos elementos celulares foram realizadas utilizando o sistema de processamento Image Pro Express, versão 6.0 para Windows, utilizando imagens obtidas por uma câmera de vídeo Olympus Q Color 5 acoplada ao microscópio Olympus BX41-BF-I-20. As descrições anatômicas, medidas, contagens de elementos celulares da madeira, bem como as terminologias utilizadas, seguiram geralmente as recomendações do Comitê IAWA (1989).
Amplificação e análise dos locos microssatélites do genoma
Para estimar os tamanhos dos fragmentos de DNA amplificados (alelos), foi utilizado o padrão interno GeneScan LIZ 600 (Life Technologies, Inc.). As estimativas dos tamanhos dos produtos fluorescentes amplificados foram realizadas utilizando o software GeneMapper versão 4.1 (Life Technologies, Inc.).
Amplificação e análise das Regiões não-codificadoras do
Dois protocolos (ROGSTAD 1992; RACHMAYANTI et al. 2006) e uma extração testada permitiram a obtenção de DNA para nove espécies. Estes resultados confirmam os obtidos por RACHMAYANTI et al. 2006), utilizando os mesmos marcadores para identificar espécies lenhosas de Dipterocarpaceae. Estudos que avaliaram o código de barras do DNA utilizaram diferentes métodos para diferenciar espécies (YAN et al., 2015).
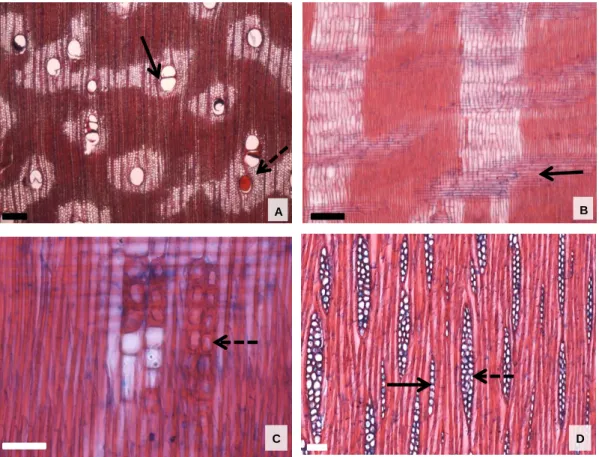
Parênquima axial: paratraqueal vasicêntrico (Figura 15B) e raramente em forma de diamante; séries de 2 a 8 células de altura, com média de 388,7 μm (± 91,1). Parênquima axial: vasicêntrico paratraqueal (Figura 19 A e B), raramente em forma de diamante e faixas confluentes; séries de 1 a 5 células de altura, com média de 331,2 μm (± 84,9). Parênquima axial: estratificado (Figura 21C); paratraqueal aliforme linear e confluente; parênquima axial em linhas de até 3 células de espessura (Figura 21A);
Embora Machado (2010) também tenha descrito a ocorrência de raios exclusivamente homocelulares na análise da madeira Tachigali denudata, Macedo et al. 2014) observaram a presença de raios heterocelulares. Entretanto, não há relação taxonômica entre a ocorrência de parênquima axial e espécies do mesmo grupo ou gênero (SILVA et al., 1989; EVANS et al., 2006).
Análises de dados genéticos…
Resultados e Discussão
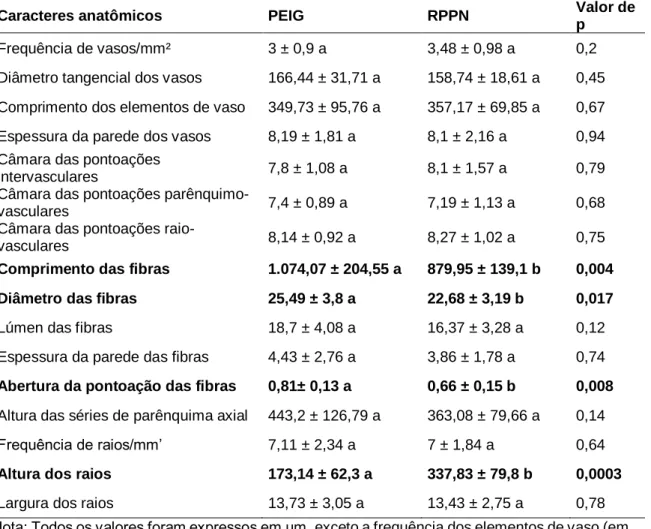
Fibras gelatinosas foram observadas em indivíduos da Floresta Estacional Semidecidual da Mata Atlântica, em Minas Gerais (Figura 1C). As variáveis que contribuíram para a separação das populações foram: comprimento e diâmetro das fibras, pontos de fibras e presença de fibras gelatinosas (Tabela 7 e Figura 2B).
Conclusão
Análise de componentes principais (PCA)
As características anatômicas de maior contribuição, em ordem decrescente de contribuição para PCA (Figura 5 B e Tabela 9), são: parênquima axial aliforme em forma de diamante, parênquima axial confluente, parênquima axial difuso, parênquima axial em linhas ou faixas estreitas, raio largura, raios com 1 a 3 células de largura, raios exclusivamente unisseriados, lúmen das fibras e diâmetro do vaso. O Grupo 1 foi dividido devido ao diâmetro do vaso, presença de parênquima axial difuso, parênquima axial aliforme e confluente em forma de diamante.
Discussão
A PCA mostrou a formação de dois grupos distintos, confirmando a divisão proposta por SCALON (2007) e SIMON et al. polystachyum, foram individualizados de espécies multifolioladas pela presença de parênquima axial paratraqueal difuso e parênquima axial aliforme em forma de diamante. polystachyum foram individualizados em PCA. A largura do raio no número de células é destacada por Evans et al. 2006) como importante fonte de diagnóstico para outras espécies e gêneros de Mimosoideae, e Marchiori (1990) para espécies e gêneros de outras subfamílias de Leguminosae. A falta de demarcação entre as espécies policotiledôneas observada na análise de PCA confirma os estudos filogenéticos de Simon et al.
Isto pode explicar a ausência de caracteres diagnósticos na anatomia da madeira das espécies estudadas neste trabalho e também na anatomia da casca relatada por Sanches et al.
Resultados
Extração de DNA
Amplificação dos locos microssatélites e regiões intergênicas do
- Microssatélites do genoma do cloroplasto (cpSSR)
- Espaçadores intergênicos
Um total de 34 alelos foram observados para os quatro locos cpSSR e o número de alelos por loco variou de 6 a 10 para as nove espécies de Leguminosae analisadas. As relações entre os haplótipos, baseadas na análise conjunta dos quatro locos cpSSR, inferidas através de uma análise de rede utilizando o algoritmo Median-Joining, demonstram a inexistência de um haplótipo ancestral comum do qual os demais haplótipos são derivados das nove espécies . estudado (Figura 8). A análise conjunta das duas regiões intergênicas do cpDNA permitiu a identificação de 11 haplótipos para as nove espécies (Anexo II).
O método de análise filogenética de vizinho utilizado para avaliar a capacidade de discriminação de espécies e a relação filogenética entre gêneros com base nos loci psbA-trnH, trnL-F e na combinação psbA-trnH+trnL-F permitiu a recuperação de grupos monofiléticos agrupando amostras pertencentes para cada uma das nove espécies analisadas.

Discussão
- Sucesso na extração e amplificação de DNA a partir de amostras
- Identificação de espécies utilizando marcadores microssatélites
- Identificação de espécies utilizando regiões intergênicas do
- Identificação de espécies utilizando regiões intergênicas do
- A identificação de Pterocarpus rohrii
O DNA extraído da madeira geralmente está bastante degradado (DEGUILLOUX et al., 2002) e no caso da madeira processada a degradação das moléculas de DNA é ainda maior (RACHMAYANTI et al., 2009). A alta taxa de recuperação de espécies em psbA-trnH pode ser explicada pela grande variação no comprimento das sequências entre as espécies de Leguminosae analisadas, o que permite maior divergência genética entre as espécies, como também observado por Yan et al. 2015) principalmente pela presença de inserções e exclusões. Nesse sentido, Bhagwat et al. 2018) que realizaram análises de DNA barcoding com os gêneros Pterocarpus e Dalbergia, respectivamente, encontraram melhores resultados ao combinar marcadores, em abordagem multilocus, utilizando diferentes combinações com o marcador matK, outra região não codificadora do Genoma Cloroplasto das Angiospermas.
Conforme sugerido por YAN et al., 2015, é importante que métodos complementares sejam incluídos na análise, pois o poder de resolução de espécies é um parâmetro importante na avaliação do método de código de barras do DNA.
Conclusão
Embora seja descrito que o método TaxonDNA possui maior poder de discriminação, uma vez que este método se baseia na identificação de espécies por meio do emparelhamento de sequências analisadas com base na distância genética observada e comparação com as sequências do banco de dados (MEIER et al. , 2006; BOLSON et al. ., 2015; HARTVIG et al., 2015; AUSTERLITZ et al., 2009; HUANG et al., 2015), outros métodos utilizados (árvores de rede e de junção de vizinhos) também apresentaram alta eficiência nas espécies identificação. Em um estudo filogenético do gênero Pterocarpus, Saslis-Lagoudakis et al. 2011) propôs uma revisão taxonômica para Pterocarpus rohrii por ser uma espécie altamente variável e polifilética. Embora baixas taxas de recuperação de identificação tenham sido observadas em Pterocarpus rohrii usando o método de código de barras para todas as espécies, o locus trnL-F provou ser mais informativo que o locus trnH-psbA. Portanto, trnL-F pode ser um locus promissor para estudos com a espécie, conforme mencionado por Klitgard et al. 2018), que analisou seis espécies de Pterocarpus, apresentou a combinação multilocus matK + ndhF-rpl32 + ITS2 como a mais adequada para análises de código de barras de DNA em espécies de Pterocarpus.
Finalmente, a abordagem multilocus utilizada no presente estudo, considerando marcadores microssatélites cpDNA e tecnologia de código de barras de DNA, foi bastante eficaz para identificação.
Resultados
Descrições anatômicas
- Subfamília Caesalpinioideae
- Subfamília Caesalpinioideae - Clado Mimosoide
- Subfamília Papilionoideae
Inclusões minerais: presença de cristais prismáticos em câmaras formando arranjos cristalinos no parênquima axial (Figura 15E). Inclusões minerais: presença de cristais prismáticos em câmaras formando arranjos cristalinos no parênquima axial (Figura 16E). Inclusões minerais: presença de cristais prismáticos em câmaras formando arranjos cristalinos no parênquima axial (Figura 19E).
Inclusões minerais: presença de cristais prismáticos em câmaras formando séries cristalinas no parênquima axial (Figura 20D).
Análise de componentes principais (PCA)
Chave de identificação
Discussão
Entretanto, não observamos a ocorrência de vasos bloqueados por tilos em nenhuma das oito espécies analisadas. Metcalfe & Chalk (1950) e Baretta-Kuipers (1981) descreveram caracteres típicos da subfamília Papilionoideae; estratificação de todos os elementos celulares; presença de parênquima axial abundante nos tipos paratraqueais vasicêntrico e aliforme, confluente, em alguns gêneros em faixas marginais e raramente ocorrência de fibras septadas. Segundo Baretta-Kuipers (1981), a ocorrência de raios uniformes e inferiores pode indicar maior especialização entre Leguminosae.
As oito espécies analisadas apresentaram grande variação no tipo de parênquima axial, sendo possível observar o aparecimento do parênquima em faixas e listras com maior frequência em Lonchocarpus cultratus e Pterocarpus rohrii.
Conclusão
Proceedings of the International Workshop "Fingerprinting Methods for the Identification of Timber Origins", Bonn, Germany. Journal of Systematics and Evolution, v. Wood anatomy of the tribe Swartzieae with comments on related Papilionoid and Caesalpinioid Leguminosae. editor), Advances in Legume Systematics 10, Higher Level Systematics: pp. DNA barcoding for the identification of the endangered species Aquilaria sinensis: comparison of data from heated or aged wood samples.
DNA barcoding of the endangered Aquilaria (Thymelaeaceae) and its application in species verification of marketed agarwood products. A comparison of the wood anatomy of 11 species from two cerrado habitats (cerrado s.s. and adjacent gallery forest). DNA barcoding of Rhododendron (Ericaceae), the largest Chinese plant genus in biodiversity hotspots of the Himalayan Hengduan.