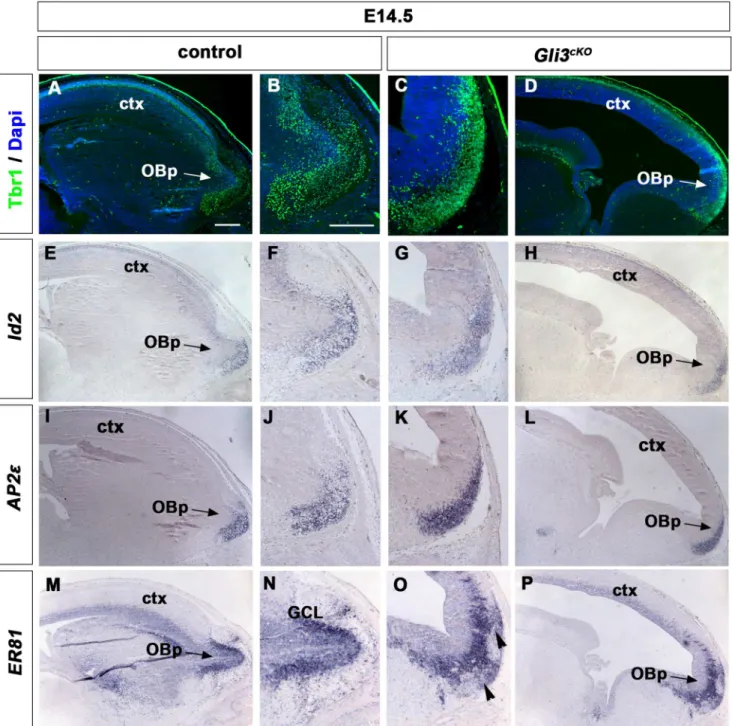
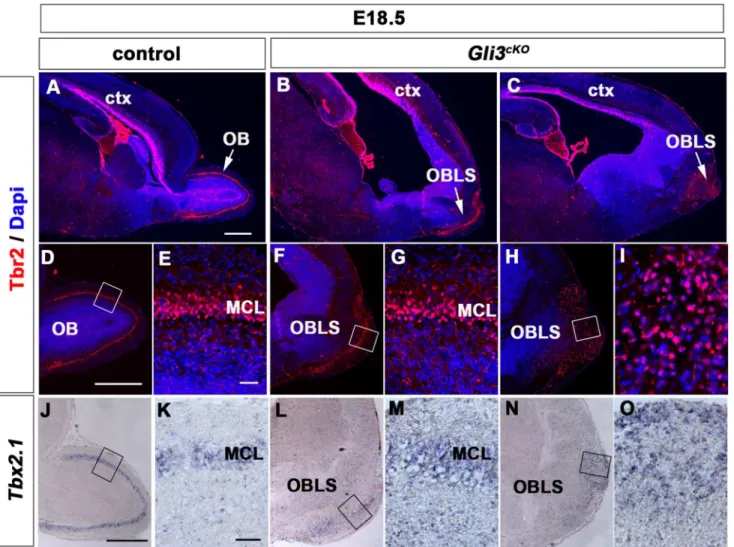
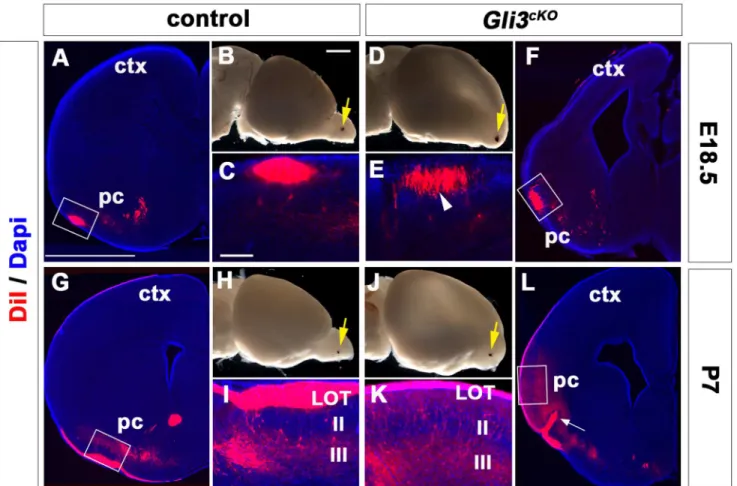
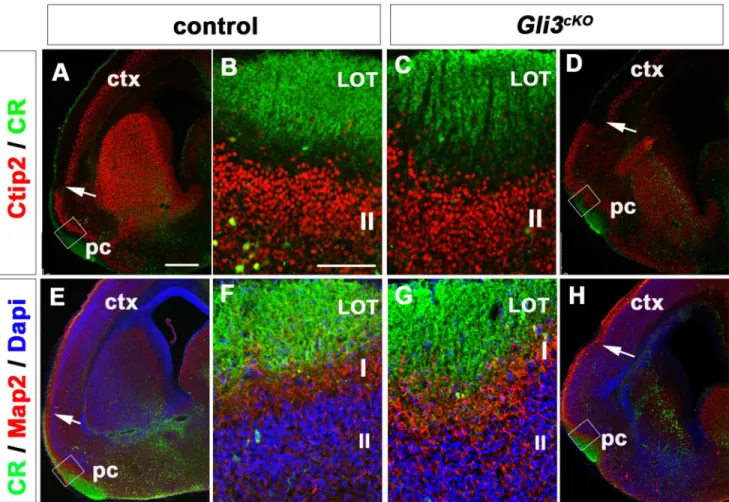
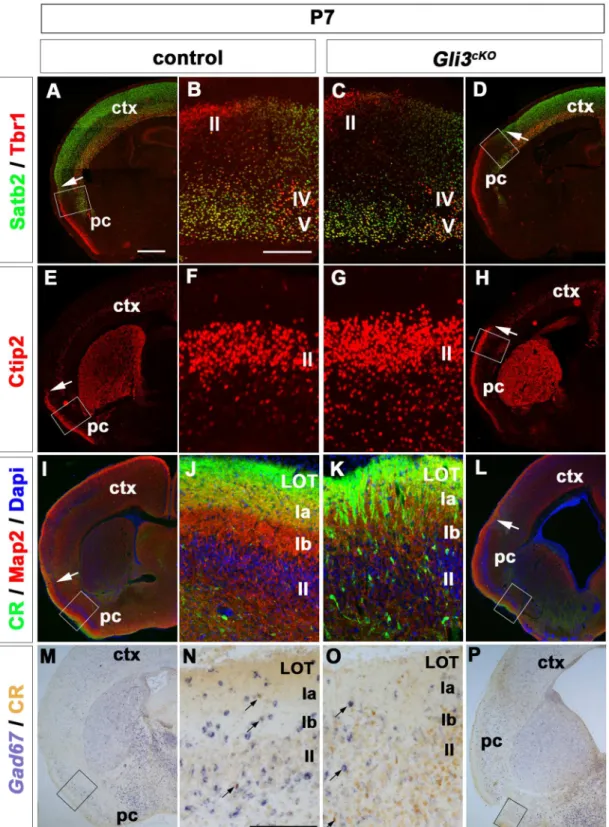
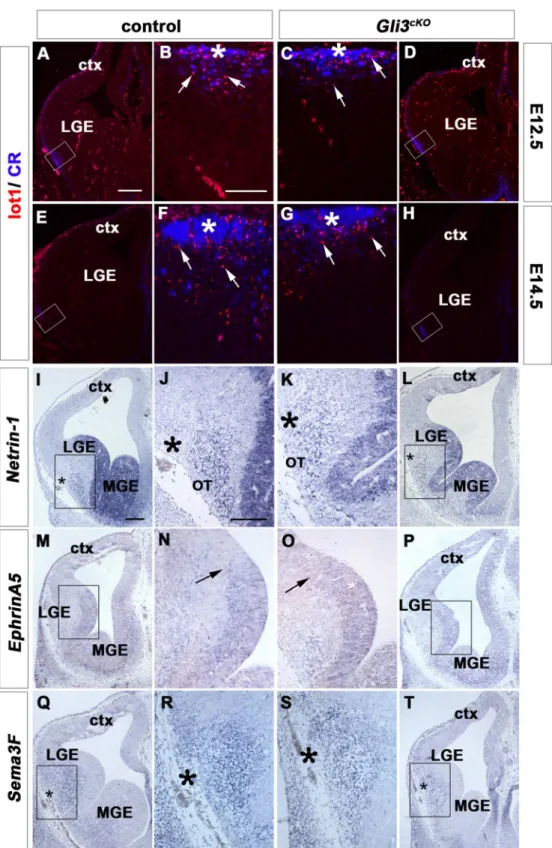
Cerebral Cortex Expression of Gli3 Is Required for Normal Development of the Lateral Olfactory Tract.
Texto
Imagem

Documentos relacionados
Our finding that Jun was strongly expressed in the OFT endocardial cushion mesen- chyme (Fig. 1D, E) together with the prominent endocardial cushions noted in Jun null embryos
Taken together, these data indicate that ATRA provides a stimulus for differentiation of t(15;17) APL cells and this results in upregulation of SIRP a expression to a level that
Patients with OSCC, particularly alcohol drinkers, current smokers, and those with the ALDH-2 allele and multiple LVL of the oesophageal mucosa, have an increased risk of
Taken together, these results showed that downregulation of Oct4 expression is clearly required to activate Stra8 and Sycp3 expression during the onset of meiosis in developing
At the first stage of the measurements results analysis, the gear wheel cast surface image was compared with the casting mould 3D-CAD model (fig.. Next, the measurements results
The structure of the remelting zone of the steel C90 steel be- fore conventional tempering consitute cells, dendritic cells, sur- rounded with the cementite, inside of
Responsiveness to activation of Shh- signaling was concluded with the following observations (i) accumulation of GLI1 protein and concurrent inhibition of GLI3 repressor formation
Taken together, our results show that the function of the ESCRT-0 complex is required for efficient FGFR signalling in different aspects of tracheal cell development,
