George Harisson Felinto Sampaio
Universidade Federal do Rio Grande do Norte
Centro de Ciências da Saúde
Programa de Pós-Graduação em Ciências Farmacêuticas
Natal, RN
2010
Soroprevalência da infecção pelo
Trypanosoma cruzi
no município de Caicó,
RN, e caracterização genética por RAPD do
parasito isolado de humanos e de
George Harisson Felinto Sampaio
Orientadora: Profa. Dra. Lúcia Maria da Cunha Galvão Pesquisadora Bolsista DCR/CNPq/FAPERN
PPgCF/CCS/UFRN
Coorientador: Prof. Titular Dr. Egler Chiari
Universidade Federal de Minas Gerais Instituto de Ciências Biológicas
Departamento de Parasitologia
Colaboradores
Prof. Dra. Antonia Cláudia Jácome da Câmara
Departamento de Análises Clínicas e Toxicológicas, DACT/CCS/UFRN
Prof. Dr. Paulo Roberto Medeiros de Azevedo Departamento de Estatística, CCET, UFRN
Dra. Daniella Alchaar D’ Ávila
Departamento de Parasitologia, ICB/UFMG
Natal, RN 2010
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Ciências Farmacêuticas (PPgCF), Departamentos de Farmácia e Análises Clínicas e Toxicológicas, Centro de Ciências da Saúde (CCS) Universidade Federal do Rio Grande do Norte, como requisito parcial para obtenção do grau de Mestre.
Instituições envolvidas na realização do trabalho
O trabalho experimental desta dissertação foi realizado no Laboratório de Biologia de Parasitos e Doença de Chagas, Departamento de Análises Clínicas e Toxicológicas, Centro de Ciências da Saúde, UFRN, e Laboratório de Biologia do Trypanosoma cruzi e doença de Chagas, Departamento de Parasitologia, Instituto de Ciências Biológicas, UFMG.
Universidade Federal do Rio Grande do Norte
Universidade Federal de Minas Gerais
Secretaria Municipal de Saúde Pública, Caicó/RN
Instituto Chico Mendes de Conservação e Biodiversidade Estação Ecológica do Seridó (ESEC SERIDÓ-RN)
Apoio Financeiro
Editais: Universal 02/2006; MCT/CNPq/MS-SCTIE-DECIT 25/2006-Estudo de Doenças Negligenciadas;MCT/CNPq/CT-Saúde/MS/SCTIE/DECIT 34/2008 Doenças Negligenciadas; Bolsas Pesquisador Visitante CNPq e DCR/CNPq/FAPERN
PPSUS/FAPERN PROJ_430_19969656 e
Agradecimentos
A DEUS, por me dar força nos momentos mais fáceis e, principalmente nos mais difíceis.
Aos meus Pais, LINDA E BÔSCO, por acreditarem que a melhor maneira de investir nos filhos é por
meio da educação.
À Profa. Lúcia Maria da Cunha Galvão, pela orientação, por ter acreditado em mim e também por
ter me dado à oportunidade de conviver ao lado de uma pessoa com tamanha sabedoria.
Ao Prof. Egler Chiari, por ter colocado a disposição as facilidades do seu Laboratório Biologia do
Trypanosoma cruzi e Doença de Chagas, Departamento de Parasitologia, ICB/ UFMG, para a
realização dos experimentos com RAPD.
À Dra. Daniella D’ Ávila, pelo acolhimento no Laboratório de Biologia do Trypanosoma cruzi e
Doença de Chagas, Departamento de Parasitologia, ICB/ UFMG, pela ajuda na realização da técnica
de RAPD e também pelos conhecimentos compartilhados.
À Simone Silveira Dias, bolsista do Laboratório de Biologia do Trypanosoma cruzi e Doença de
Chagas, Departamento de Parasitologia, ICB/ UFMG, pela ajuda durante o estágio na Instituição.
À Profa. Antônia Claúdia Jácome da Câmara, pela valiosa colaboração e companheirismo na rotina
do laboratório, pela paciência e pelos conhecimentos.
Ao Prof. Geraldo Barroso, pelos conselhos e incentivos.
Aos Professores do Programa de Pós-Graduação em Ciências Farmacêuticas, Centro de
Ciências da Saúde, UFRN, representados pela Coordenadora, Profa. Adriana Augusto de Rezende,
pelos ensinamentos transmitidos.
Aos meus amigos Diêgo Sollon, César Rodrigues, Danielle Delgado e Ramon Brito pela ajuda e
conselhos no decorrer dessa jornada.
Aos funcionários do IBAMA, Juan e George, pela atenção e colaboração.
As funcionárias Fábia Freire e Aureliana Bezerra do Programa de Pós-Graduação em Ciências
Farmacêuticas, Centro de Ciências da Saúde, UFRN, pelo exemplo de dedicação e profissionalismo.
Às pessoas que aceitaram participar deste trabalho, minha grande gratidão.
LISTA DE GRÁFICOS, TABELAS E FIGURAS
Gráfico 1 - Percentual de amostras soro reativas, não reativas e indeterminadas
dos moradores de diferentes comunidades rurais do município de Caicó, RN... 27 Gráfico 2 - Percentual de indivíduos com sorologia reativa ou indeterminada e não
reativa por gênero das diferentes comunidades rurais do município de Caicó, RN... 31 Tabela 1 - Iniciadores aleatórios utilizados nas reações de RAPD-PCR... 23
Tabela 2 - Características dos isolados do Trypanosoma cruzi obtidos de indivíduos infectados e de triatomíneos naturalmente infectados... 26 Tabela 3 - Reatividade sorológica da infecção pelo Trypanosoma cruzi em
moradores das diferentes comunidades rurais do município de Caicó, RN... 28 Tabela 4 - Reatividade sorológica para anticorpos anti-T. cruzi por faixa etária dos
390 moradores das comunidades rurais no município de Caicó, RN... 32 Tabela 5 - Reatividade sorológica para anticorpos anti-T. cruzi por escolaridade
dos 390 moradores das diferentes comunidades rurais do município de Caicó, RN.. 33 Figura 1 – Mapa do Estado do Rio Grande do Norte em destaque os municípios estudados e localização da Estação Ecológica do Seridó, área de 1.166ha (ESEC, criada pelo decreto 87.222 de 21/05/1982, Lei No. 6.092 de 27/04/1982).
Fonte: IBGE mapas... 25 Figura 2 - Gel poliacrilamida a 5% representativos dos perfis de RAPD do T.
cruzi, obtidos de indivíduos e triatomíneos naturalmente infectados, com o
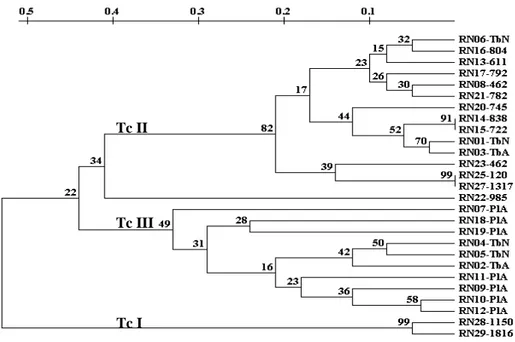
iniciador M13-40. Nas canaletas das extremidades, o marcador de peso molecular (PM), e na penúltima, controle negativo da PCR (branco). As cores representam grupos de isolados com elevado compartilhamento de bandas entre eles... 35 Figura 3 - Fenograma correspondente aos perfis de RAPD de isolados do T. cruzi
obtidos de indivíduos e triatomíneos naturalmente infectados, construído pela UPGMA usando matriz de distância genética obtida com o iniciador M13-40. As percentagens correspondem aos valores bootstrap (percentagens de vezes em que o agrupamento se manteve durante 1000 reamostragens)... 36 Figura 4 - Fenograma correspondente aos perfis de RAPD de isolados do T. cruzi
Figura 5 - Fenograma correspondente aos perfis de RAPD de isolados do T. cruzi
obtidos de indivíduos e triatomíneos naturalmente infectados, construído pela UPGMA, usando matriz de distância genética obtida com o iniciador λgt11-F. As percentagens correspondem aos valores bootstrap (percentagens de vezes em que o agrupamento se manteve durante 1000 reamostragens)... 40 Figura 6 - Fenograma correspondente aos perfis de RAPD de isolados do T. cruzi
LISTA DE ABREVIATURAS
ANVISA Agencia Nacional de Vigilância Sanitária CRP Complement Regulatory Protein
RAPD Random amplified polymorphic DNA
EDTA Ácido etlilenodiamino tetracético
ELISA Enzyme-Linked Immunosorbent Assay
HAI Hemaglutinação indireta
kDNA DNA do cinetoplasto
LIT Liver Infusion Tryptose
min Minuto
mL Mililitro
mM Milimolar
ng/µl Nanograma/microlitros
pb Pares de base
RIFI Reação de Imunofluorescência Indireta
RN Rio Grande do Norte
rpm Rotações por minutos
rRNA RNA ribossomal
SDS Sodium dodecyl sulphate
TESA Trypomastigote excreted/secreted antigen
µ L Microlitro
UPGMA Unweighted PairGroup Mean Average
RESUMO
ABSTRACT
Trypanosoma cruzi infection was evaluated in 390 resident individuals in different rural
SUMÁRIO
1. INTRODUÇÃO 12
1.1 Trypanosoma cruzi e a doença de Chagas 12
1.2 Diagnóstico laboratorial 15
1.3 Random amplified polymorphic DNA (RAPD) 17
2. OBJETIVOS 20
2.1 Objetivo geral 20
2.2 Objetivos específicos 20
3. MATERIAL E MÉTODOS 21
3.1 Seleção dos indivíduos 21
3.2 Isolamento e cultura acelular do Trypanosoma cruzi 22 3.3 Preparação do DNA genômico do Trypanosoma cruzi 23 3.4 RAPD (Random Amplified Polymorphic DNA) 24 3.5 Eletroforese em gel de poliacrilamida 25 3.6 Análise dos dados e construção dos fenogramas 25 3.7 Caracterização genética das amostras do T. cruzi 25
4. RESULTADOS E DISCUSSÃO 28
4.1 Soroprevalência da infecção pelo T. cruzi 28 4.2 Avaliação da variabilidade intraespecífica das populações do T.
cruzi pela técnica de RAPD
35
5. CONCLUSÕES 46
6. REFERÊNCIAS 47
7. ANEXOS 57
7.1 Termo de consentimento livre e esclarecido - Adultos 57 7.2 Termo de consentimento livre e esclarecido – Crianças 7-12 anos 60 7.3 Termo de consentimento livre e esclarecido – Pais de
adolescentes 13-17 anos 63
7.4 Termo de consentimento livre e esclarecido – Adolescentes de
13-17 anos 66
7.5 Questionário para identificação individual 69
7.6 Compartilhamento de bandas dos isolados do T. cruzi de indivíduos chagásicos e triatomíneos obtidos pelo programa
DNA-POP com o iniciador M1340 70
indivíduos chagásicos e triatomíneos obtidos pelo programa DNA-POP com o iniciador L15996
7.8 Compartilhamento de bandas dos isolados do T. cruzi de indivíduos chagásicos e triatomíneos obtidos pelo programa DNA-POP com o iniciador gt11-F
1 INTRODUÇÃO
1.1 Trypanosoma cruzi e a doença de Chagas
A tripanossomíase americana é uma infecção de evolução essencialmente crônica causada pelo hemoflagelado Trypanosoma cruzi e transmitido naturalmente ao homem e a outros animais por insetos vetores da ordem Hemíptera e família Reduviidae. Esta enfermidade é originalmente confinada a pobreza nas áreas rurais das Américas Central e do Sul. Um fator preocupante é que esta doença está gradativamente se tornando um problema de saúde global devido o fluxo migratório de indivíduos de 17 países da América Latina para os países desenvolvidos como, por exemplo, Austrália, Canadá, Espanha e Estados Unidos da América (SCHMUNIS, 2007). Após os bem sucedidos programas de controle ao vetor nas décadas de 80 e 90, em 21 países onde ocorre a endemia, o número de indivíduos infectados atualmente é estimado em 7,7 milhões e 41.200 casos novos/ano ocorrem por transmissão vetorial. No Brasil o número estimado de indivíduos infectados é aproximadamente dois milhões (SALVATELLA, 2007).
No Brasil foram realizados dois inquéritos sorológicos para verificar a prevalência da infecção pelo T. cruzi. No primeiro inquérito (1975-1980) a prevalência para a população geral rural brasileira foi estimada em 4,2% e 1,8% para o Estado do Rio Grande do Norte (CAMARGO et al., 1984). O segundo inquérito foi realizado (1989-1997) em escolares de 7-14 anos de idade e demonstrou que a média de prevalência da infecção chagásica no Brasil foi de 0,14%, e, 0,20% para o Estado do Rio Grande do Norte (RN) em um total de 116 municípios analisados (SILVEIRA & VINHAES, 1998). A área endêmica para a doença de Chagas no Estado do RN compreende aproximadamente 115 municípios e destes, apenas 12 passaram à fase vigilância epidemiológica em 1995 (FUNASA, 1989-1995). Observou-se ainda que, no período entre 1993-1996, não houve alteração em relação ao número de triatomíneos capturados nas unidades domiciliares, permanecendo prevalentes as espécies
Triatoma brasiliensis e T. pseudomaculata, independente das ações realizadas em todos esses
anos do programa de controle (SILVEIRA & VINHAES, 1998).
áreas onde o vetor silvestre pode invadir domicílios, pelo custo médico e social no cuidado de indivíduos infectados com o tratamento específico (MOREL & LAZDINS, 2003).
A doença de Chagas é caracterizada por duas fases distintas: a fase aguda e a fase crônica. A primeira com duração e intensidade variáveis tem sua definição baseada na elevada parasitemia detectável por exames parasitológicos diretos do sangue, sendo uma infecção predominante em crianças na primeira década de vida. Na passagem da fase aguda para a crônica estão envolvidos importantes fenômenos de imunomodulação, ocorrendo um declínio acentuado de parasitemia e redução progressiva dos fenômenos inflamatórios. A fase crônica é caracterizada por apresentar longa duração com parasitemia subpatente e, elevado e consistente aumento de anticorpos da classe IgG. Essa fase geralmente inicia-se com a forma indeterminada também denominada latente, subclínica ou inaparente na qual a maioria dos pacientes (60-70%) permanece sem qualquer manifestação clínica, eletrocardiograma, RX do coração, cólon e esôfago normais (COURA et al., 1984).
A resposta imune na fase aguda é constituída de hipersensibilidade celular no sítio de entrada e reação inespecífica inicial, por meio de macrófagos e neutrófilos. Anticorpos heterófilos inatos encontrados no soro de indivíduos normais, reconhecem o T. cruzi e a eles se ligam (TOWBIN et al., 1987). Estes anticorpos reconhecem um epítopo terminal contendo o dissacarídeo galactosil 1-3 galactose (anti-Gal), e possuem atividade lítica contra tripomastigotas, independentemente de complemento. A formação de anticorpos específicos da classe IgM é relativamente precoce, iniciando-se ao fim da primeira semana e mantendo níveis elevados durante toda a fase aguda. A invasão de macrófagos do hospedeiro por tripomastigotas inicia a resposta inata pelo recrutamento e ativação de células natural killer (NK) e linfócitos T, envolvendo a secreção de IL-12 e fator de necrose tumoral pelos macrófagos. Já as células NK atuam sobre os macrófagos, induzindo a produção de óxido nítrico e a eliminação de formas intracelulares do parasito e/ou promovendo o desenvolvimento da imunidade celular adquirida (SCOTT & TRINCHIERI, 1995). Vale destacar também que, essas células são importantes produtoras de IFN- e TNF- , duas citocinas relevantes na ativação de outras células, como, macrófagos, tornando-as mais eficientes na destruição de microorganismos intra e extracelulares. Todavia, também tem sido demonstrado que, macrófagos infectados por T. cruzi secretam IL-12, levando ao aumento da produção de IFN- e TNF- , e o consequente controle da parasitemia e da mortalidade (ALIBERTI et al., 1996; HUNTER et al., 1996).
(D’IMPÉRIO LIMA et al., 1986). Nessa fase a resposta imune anti-T. cruzi é mista, uma vez que diferentes componentes do sistema imune como, por exemplo, anticorpos, linfócitos T CD4+ e CD8+ e macrófagos são essenciais para o controle da infecção em camundongos, além disso, é caracterizada por uma sólida resposta humoral protetora que mantém uma parasitemia sub-patente (RUSSO et al., 1988; TARLETON, 1990). As citocinas desempenham papel importante na regulação da resposta imune, e estão envolvidas tanto na resistência quanto nos mecanismos relacionados com a imunopatologia da infecção pelo T.
cruzi. O IFN- tem sido considerado uma citocina protetora, uma vez que ao estimular
macrófagos, estes produzem metabólitos tóxicos para o parasito, por outro lado IL-4, IL-10 e TGF- são capazes de suprimir a ativação dos macrófagos induzida por IFN- , inibindo tanto a liberação dos metabólitos tóxicos quanto a diferenciação de células Th1 (BRODSKYN & BARRAL-NETO, 2000).
Apesar de, os mecanismos efetores do sistema imunológico contra o T. cruzi, uma série de evidências tem demonstrado que diversos mecanismos de evasão à resposta imune são acionados durante a infecção contribuindo para a cronicidade e para as alterações patológicas associadas à infecção chagásica. Dentre eles ressaltamos a resistência das formas infectantes tripomastigotas à ação do complemento, uma vez que, esse parasito possui glicoproteínas capazes de bloquear a via alternativa e/ou clássica do sistema complemento, inibindo a formação da C3 convertase, enzima central da cascata do sistema (JOINER et al., 1988; TAMBOURGI et al., 1993; NORRIS et al., 1991; FISCHER et al., 1988). O ácido siálico presente na superfície das formas tripomastigotas, também contribui para a resistência do parasito ao complemento (SCHENKMAN et al., 1991).
parte, a eficácia dessa resposta imune e aos aspectos genéticos dos indivíduos infectados, mas tem sido relatado que se deve primariamente a complexidade da estrutura populacional do T.
cruzi (MILES et al., 1981). Em países como, o Panamá e a Venezuela há predominância da
forma cardíaca. No Brasil, Argentina e Chile as formas clínicas são bastante variadas (COURA et al., 1983; GUEVARA et al., 1977). Em alguns Estados do Brasil como, a Bahia, predomina a forma cardíaca, em Goiás, a forma digestiva, em Minas Gerais observa-se uma elevada frequência de formas mistas (cardíaca e digestiva) e, no Rio Grande do Sul e Rio de Janeiro a maioria dos indivíduos infectados é assintomática (COURA et al., 1983).
1.2 Diagnóstico laboratorial
O diagnóstico laboratorial da infecção pelo T. cruzi pode ser realizado por diferentes métodos de acordo com as fases da infecção. Na fase aguda, o diagnóstico parasitológico é prioridade, pela identificação das formas tripomastigotas do parasito ao exame direto do sangue periférico. Os exames diretos do sangue são realizados rapidamente, sendo o resultado imediato, por isso devem ser empregados em toda suspeita de fase aguda ou de reativação, por qualquer forma de transmissão (CERISOLA, 1974; CANÇADO, 1980). A utilização de métodos sorológicos para pesquisa de anticorpos anti-T. cruzi da classe IgM no sangue periférico nessa fase da infecção pelo T.cruzi, tem encontrado dificuldades devido a falta de
kits comerciais registrados na ANVISA (Agencia Nacional de Vigilância Sanitária) e carência
de controles positivos para IgM (MS, 2005).
Na fase crônica, devido à parasitemia subpatente, os métodos parasitológicos são de baixa sensibilidade, enquanto, os métodos sorológicos detectam anticorpos anti-T. cruzi da classe IgG e devem ser realizados dois métodos de princípios distintos ou com diferentes preparações antigênicas com sensibilidade e especificidade elevadas (MS, 2005). O xenodiagnóstico e a hemocultura são métodos parasitológicos indiretos altamente específicos e podem ser empregados tanto na fase aguda como na fase crônica da infecção, mas não são viáveis para aplicação na rotina laboratorial uma vez que, são laboriosos e demandam um tempo mais longo para obtenção do resultado (CHIARI & DIAS, 1975).
Os métodos sorológicos convencionais como, imunofluorescência indireta (IFI),
Enzyme-Linked Immunosorbent Assay (ELISA) e hemaglutinação indireta (HAI) são capazes
1993) apresenta elevada sensibilidade na fase crônica da doença e baixa na fase aguda (MATSUMOTO et al., 1993; UMEZAWA et al., 1993; UMEZAWA et al., 1996). Uma vez que, a fração antigênica das formas epimastigotas é composta de moléculas complexas que favorecem a ocorrência de reações falso-positivas e reações cruzadas com soros de indivíduos infectados com Leishmania sp. e T. rangeli (CHILLER et al., 1990; ARAÚJO, 1986; SCHECHTER et al., 1983). Esta heterogeneidade antigênica inviabiliza o diagnóstico diferencial entre as fases aguda e crônica e, entre as formas clínicas da doença de Chagas (UMEZAWA et al., 2001).
Os ensaios sorológicos são bastante sensíveis, entretanto seu desempenho pode variar em função dos reagentes, procedimentos técnicos e diferentes critérios de avaliação dos resultados (CAMARGO et al., 1984). Resultados falsos positivos podem ocorrer em função da variação nos títulos de anticorpos na fase crônica ou transitoriamente negativos e, nos casos de indivíduos sorologicamente negativos, porém parasitologicamente positivos (BRENIERE et al., 1989). Além disso, eles ocorrem também por reatividade antigênica cruzada com outros protozoários flagelados ou ainda, a presença de auto-anticorpos capazes de reagir com lipídeos, carboidratos e epítopos polipeptídicos comuns aos parasitos e ao hospedeiro (VELÁSQUEZ et al., 1993).
Os métodos sorológicos na fase crônica da infecção contribuem para: (i) realizar diagnósticos individuais; (ii) levantar níveis de endemicidade; (iii) avaliar medidas de controle de transmissão; (iv) selecionar doadores de sangue; (v) avaliar a atividade de drogas; e (vi) acompanhar a resposta imune (GOMES, 1996).
A IFI constitui um dos métodos mais utilizados no diagnóstico laboratorial da infecção chagásica crônica devido à relativa facilidade de obtenção de reagentes padronizados, elevada sensibilidade e praticidade para o uso em larga escala. Contudo, a leitura é subjetiva e sua especificidade é baixa já que favorece a reações cruzadas com outras parasitoses. A reação de ELISA apresenta vantagens, tais como, a sua elevada sensibilidade, a utilização de quantidade mínima de soro, a estabilidade dos seus reagentes, a facilidade de pré-sensibilizar as placas com antígenos e congelá-las até o momento de uso, o processamento de várias amostras em uma só placa, a utilização de espectrofotômetros portáteis para a leitura dos resultados no campo, fácil execução e a possibilidade de automação. No entanto, a especificidade é variável e pode não ser a ideal com antígenos brutos ou semipurificados (ZICKER et al., 1984). Os antígenos recombinantes originados de frações citoplasmáticas e flagelar (KRIEGER et al., 1992), a proteína recombinante de 24kDa rTc24 (KRAUTZ et al., 1995) tem sido testados com sucesso revelando elevada especificidade. O antígeno TESA (Trypomastigote
ELISA num grupo de 120 pacientes chagásicos nas fases aguda e crônica demonstrando 100% de sensibilidade e especificidade nas duas fases da doença (UMEZAWA et al., 2001). A glicoproteína de 160kDa (GP160), uma proteína específica das formas tripomastigotas do
T. cruzi pertencente à família das proteínas reguladoras da cascata de complemento
(Complement Regulatory Protein–CRP) foi descrita como antígeno indutor dos anticorpos líticos e, devido à elevada especificidade e sensibilidade apresenta potencial para ser utilizada no diagnóstico e como critério de cura de pacientes chagásicos e tratados (MARTINS et al., 1985; NORRIS et al., 1994; MEIRA et al., 2002).
1.3 Random amplified polymorphic DNA (RAPD)
O T. cruzi é constituído por populações heterogêneas contendo um grande número de clones naturais, que circulam nos ambientes domésticos e silvestres entre seres humanos, reservatórios e vetores (DEVERA et al., 2003). Essa heterogeneidade tem sido comprovada por meio de abordagem biológica, bioquímica, patológica, imunológica e genética entre as populações isoladas de diferentes hospedeiros e vetores (CHAGAS, 1909; BRENER & CHIARI, 1963; TIBAYRENC & AYALA, 1988; MACEDO et al., 1992; STEINDEL et al., 1993). A elevada variabilidade biológica e genética encontrada nessa espécie pode ser explicada pelos múltiplos contatos entre os vetores e reservatórios nas áreas endêmicas, os quais propiciam infecções com mais de uma população do T. cruzi, com distintas propriedades biológicas, que coexistiam dentro de um mesmo hospedeiro, em sua maioria sem recombinação entre eles, constituindo as populações monoclonais ou multiclonais (TIBAYRENC & AYALA, 1988).
Na última década, com o avanço tecnológico, o polimorfismo genético em várias espécies de microorganismos, inclusive do T. cruzi, tem sido unificado por técnicas moleculares, dentre elas podemos destacar a técnica de RAPD (Random Amplified
Polymorphic DNA) utilizada inicialmente para examinar amostras de DNA humano
(WILLIAMS et al., 1990). Essa técnica identifica loci amplificados pela PCR usando iniciadores curtos, com sequências aleatórias, que se ligam em sítios desconhecidos e dispersos do genoma, muito dos quais são polimórficos gerando produtos de DNA de tamanho e intensidade variáveis e padrões complexos de bandeamento. As informações obtidas podem ser utilizadas como marcadores genéticos de populações e de relações filogenéticas sendo utilizada tanto na diferenciação de espécies, quanto na detecção de polimorfismos de DNA (TIBAYRENC et al., 1993; STEINDEL et al., 1993).
rápido e simples e não requer trabalho preliminar. É uma técnica de fácil manuseio onde várias amostras podem ser processadas ao mesmo tempo, além de requerer quantidades mínimas de DNA para sua realização. Estas características mostram que RAPD pode ser utilizada na hibridização, identificação de clones, no desenvolvimento de marcadores ligados a determinação do sexo, e muitos outros estudos (WOLFE & LISTON, 1998). Essa técnica também revela níveis relativamente elevados de polimorfismo e tem um custo baixo quando comparada com outras técnicas, como, isoenzimas e RFLP (Restriction Fragment Lenght
Polymorphism) tornando-a ideal também para o mapeamento genético, vegetal e animal
(DEVERA et al., 2003). Vale salientar que, em muitos casos, apenas um pequeno número de iniciadores são necessários para identificar o polimorfismo entre as espécies, destacando que a simplicidade da técnica pode levar a automatização do mapeamento genético de organismos que necessitariam de um amplo número de marcadores genotípicos para descrever completamente o seu genoma (WILLIAMS et al., 1990). Todavia, apresenta algumas limitações, dentre elas, pequenas mudanças nas condições da reação podem interferir na reprodutibilidade dos produtos de amplificação, como, a temperatura de anelamento que pode interferir nas ligações do iniciador com sítios não totalmente complementares gerando o aparecimento de bandas falso-positivas (PENNER et al., 1993). A reprodutibilidade dos resultados pode ainda ser influenciada por fatores como, variações de pureza do DNA, peso molecular, quantidade da enzima Taq DNA polimerase, a concentração de cloreto de magnésio e o tamanho do iniciador. Portanto, essa técnica deve ser padronizada com os diferentes reagentes para obtenção de resultados satisfatórios antes do seu uso (STEINDEL et
al., 1993). Outro fator crítico é a dependência da interpretação pessoal do pesquisador,
tornando difícil a reprodutibilidade dos achados. Para testar a confiabilidade do RAPD, três laboratórios independentes usaram as mesmas amostras de DNA e os mesmos iniciadores e observaram que as matrizes de dados resultantes foram substancialmente diferentes com algumas conclusões divergentes, embora também tenha apresentado resultados semelhantes em laboratórios diferentes (WOLFE & LISTON, 1998).
Desde a sua introdução em 1990, o RAPD tem sido direcionado para estudos taxonômicos e para caracterização de microorganismos, demonstrando ser uma ferramenta útil tanto em tripanossomatídeos como em outros protozoários parasitos, uma vez que tem a capacidade de revelar importante informação genética variável até mesmo entre isolados de uma mesma espécie (STEINDEL et al., 1993, TIBAYRENC et al., 1993; DEVERA et al., 2003). E também tem sido utilizada para a análise genética de diversos parasitos como,
Schistosoma, Giardia, Leishmania, Trypanosoma rangeli e T. cruzi (DIAS NETO et al.,
marinhos o RAPD foi utilizado para analisar a variabilidade em ostras (LIU & DAÍ, 1998), mexilhões (KIMURA et al., 1997), entre outras espécies, e mostrou-se uma ferramenta adequada para identificar polimorfismo de populações em uma mesma espécie.
Estudos sobre genética de populações do T. cruzi com RAPD demonstraram que vários isolados do parasito provenientes de humanos, triatomíneos e gambás foram bastante heterogêneos, evidenciando que, as cepas dos zimodemas Z2 e Z3 mostraram padrões mais polimórficos (STEINDEL et al., 1993). Os perfis de RAPD gerados por 61 isolados obtidos de 47 pacientes chagásicos mostraram a existência de uma população geneticamente bem correlacionada, mas não permitiu estabelecer diferenças entre o perfil molecular gerado para cada isolado e as manifestações clínicas da doença, identificando apenas que populações do T.
cruzi isoladas de pacientes chagásicos crônicos apresentaram elevada homogeneidade
genética (D’ ÁVILA et al., 2006).
2 OBJETIVOS
2.1 Objetivo geral
Avaliar a soroprevalência da infecção pelo Trypanosoma cruzi em moradores rurais do município de Caicó, e a variabilidade genética intraespecífica de populações do parasito isoladas de indivíduos e de triatomíneos procedentes de Caicó, Serra Negra do Norte, Angicos, Governador Dix Sept Rosado e Caraúbas no Estado do Rio Grande do Norte.
2.2 Objetivos específicos
Avaliar a prevalência da infecção pelo T. cruzi em moradores de diferentes comunidades rurais do município de Caicó, RN, por métodos sorológicos e sua associação com dados epidemiológicos;
Isolar o T. cruzi de indivíduos sorologicamente reativos por hemocultura e de triatomíneos naturalmente infectados por xenocultura para estudos epidemiológicos moleculares;
3 MATERIAL E MÉTODOS 3.1 Seleção dos indivíduos
O desenho da amostra para o inquérito sorológico se baseou em “amostragem aleatória simples”, sendo que, para efeito de definição do cálculo amostral, a variável considerada é p = proporção de indivíduos com sorologia reativa no município de Caicó. Assim, de acordo com estimativa preliminar, tem-se pˆ = 0,037. Neste caso, o cálculo do tamanho da amostra é definido por:
(1 )
( 1) (1 )
Np p n
N D p p
Onde n = tamanho da amostra; N = número total de habitantes; D = 2 /4, sendo um limite para um erro estimado de p, que satisfaz: P( p- pˆ < ) = 0,95 e pˆ como o estimador da proporção de indivíduos com sorologia reagente no município de Caicó.
Considerando-se = 0,02 dado que N = 6960 e substituindo p por pˆ= 0,037, pela expressão acima obtém-se n = 339. Assim, foi realizando o sorteio das localidades com dez ou mais residência, até atingir-se um número igual ou maior a 339 indivíduos. Desta forma, alcançamos uma amostra de 358 indivíduos, onde o número que ultrapassou o obtido no cálculo amostral corresponde a 5,6% do calculado que é um percentual de acréscimo usualmente considerado nas definições de planos amostrais.
Os moradores de comunidades rurais do município de Caicó, localizado na mesorregião central do estado do Rio Grande do Norte foram previamente consultados individualmente em sua residência e obtivemos a concordância escrita por meio do Termo de Consentimento Livre e Esclarecido (TCLE) aprovado pelo comitê de Ética da Universidade Federal de Minas Gerais (UFMG), sob o Nº 312/06 - Anexos 1, 2, 3 e 4. A faixa etária desses indivíduos variou de seis a 89 anos. As amostras de sangue periférico foram coletadas por punção venosa e acondicionadas em tubos de Eppendorf (Becton Dickinson and Company, USA) devidamente identificados e congelados até o momento de uso. Os métodos utilizados para o diagnóstico da infecção pelo T. cruzi foram ELISA (Enzyme-linked immunorbent assay) empregando o kit Chagastest (Wiener Laboratórios S. A I. C., Rosário Argentina) e o de Imunofluorescência Indireta (IFI) com o antígeno de formas epimastigotas da cepa Y do T. cruzi fixadas com formaldeído a 20% (CAMARGO, 1966), usando como conjugado a imunoglobulina anti-IgG humana marcada com isotiocianato de fluoresceína (Sigma Chemical Company, Missouri,
USA). As reações de ELISA foram consideradas reativas quando a densidade óptica foi
Consenso Brasileiro em Doença de Chagas (MS, 2005). Todos os indivíduos com sorologia reativa foram encaminhados para acompanhamento clínico pela Equipe de Saúde da Família de suas localidades.
3.2 Isolamento e cultura acelular do Trypanosoma cruzi
O isolamento do parasito foi realizado nos indivíduos com sorologia reativa pela técnica de hemocultura (CHIARI et al., 1989). De cada indivíduo foram coletados 30mL de sangue venoso em tubos a vácuo contendo o anticoagulante heparina e transferidos para um tubo plástico cônico de 50mL (Falcon, Sigma). Os tubos contendo o sangue foram centrifugados a 300 × g por 10min a temperatura ambiente seguido da remoção do plasma para um tubo plástico cônico de 15mL (Falcon, Sigma). Ao sedimento de hemácias foram adicionados 10mL de meio de cultura acelular Liver infusion tryptose-LIT (CAMARGO, 1964) e centrifugados a 900 × g por 20min a 4 C. Descartado o sobrenadante, ao sedimento de hemácias foram adicionados 6mL de LIT, homogeneizado e distribuído em alíquotas de 3mL em cada tubo de 15mL contendo 3mL de meio LIT, totalizando seis tubos. O tubo contendo o plasma foi centrifugado nas mesmas condições e, após a centrifugação, transferido para novo tubo e armazenado a -20 C. Ao sedimento obtido do plasma foram adicionados 5mL de LIT. Os tubos foram incubados a temperatura de 28 C, agitação lenta duas vezes por semana e alíquotas de 10µL examinadas ao microscópio entre lâmina e lamínula com aumento de 400×, mensalmente até 120 dias. As hemoculturas positivas foram cultivadas em meio LIT por no máximo três passagens na tentativa de evitar seleção de populações do parasito por prolongada manutenção em meio de cultura. Em seguida, as culturas foram mantidas em fase de crescimento na concentração de 1 106 epimastigotas/mL em 60-80mL de LIT. A cultura foi então lavada por três vezes em tampão Krebs-Ringer-Tris (KRT) pH 7,2 e concentradas por centrifugação 1.800 × g por 15min a 4oC. O sedimento (massa úmida) foi armazenado à temperatura de -20oC até o momento da extração do DNA.
contendo solução salina estéril. Após a maceração, a suspensão foi utilizada para a xenocultura em meios de cultura acelular LIT + McNeal Novy e Niccole (NNN) ou Agar sangue, incubada a 28oC e examinada 30-60 dias (BRONFEN et al., 1989). Os tubos positivos para o T. cruzi foram semeados em meio LIT e mantidos por duas passagens sucessivas. Após esse período, o parasito foi mantido em cultura para o crescimento até a concentração de 106/ml e obtenção da massa úmida para extração do DNA.
3.3 Preparação do DNA genômico do Trypanosoma cruzi
O DNA foi obtido pela digestão do sedimento de cada isolado do T. cruzi com proteinase K (Promega, Madison, WI, USA), desproteinizado com fenol (v/v), fenol clorofórmio e clorofórmio, seguido da precipitação em etanol absoluto (MACEDO et al., 1992). Os sedimentos foram resuspendidos em 1mL de tampão de lise (NaCl 80mM/ EDTA 45mM, pH 8,0/ SDS 1%) contendo 0,1mg/mL de proteinase K e incubado a 37°C durante 12h. A extração do DNA foi realizada por desproteinização com fenol (v/v) e a fase aquosa foi recuperada e submetida a extrações com igual volume (v/v), fenol: clorofórmio: álcool isoamílico (25:24:1) e clorofórmio: álcool isoamílico (24:1). A precipitação do DNA ocorreu na presença de dois volumes de etanol absoluto gelado incubado a -70°C por 2h ou a -20°C durante a noite. Após a centrifugação e volatilização do etanol, o precipitado obtido foi tratado com 1mL de tampão da ribonuclease (NaCl 80mM/ EDTA 5mM, pH 8,0, 10U/ L de ribonuclease - Promega, Madison, Wiscosin, USA ) e incubado a 37ºC por 2h. A seguir, foram realizadas novas etapas de extração e precipitação do DNA, como descrito anteriormente. O DNA obtido foi diluído em 300µL de tampão Low TE (10mM Tris-HCl e 0,1mM EDTA pH8,0) e estocado a -20°C (MACEDO et al., 1992). As amostras de DNA foram quantificadas em espectrofotômetro a 260nm sendo o grau de pureza das amostras determinado pela relação das absorbâncias a 260/280nm. A concentração aproximada do DNA total foi determinada com auxílio da fórmula:
A concentração final de cada amostra de DNA foi ajustada para 3ng/µ L e as amostras armazenadas a 4ºC até o momento do uso.
3.4 RAPD (Random amplified polymorphic DNA)
A amplificação aleatória de polimorfismos de DNA em T. cruzi foi realizada seguindo o procedimento previamente descrito (STEINDEL et al., 1993). Cada reação foi processada em um volume final de 10µL, contendo 10mM Tris-HCl (pH 9,0), 0,1% Triton X-100, 1,5mM MgCl2, 50mM KCl, 125µM de cada dNTP, 1,0 unidade de Taq DNA polimerase, 6,4pmol do
Concentração DNA= Leitura a 260nm × Fator 50 × Fator da diluição = g/ l
iniciador e 1ng/µ L de DNA, adicionados de 20µL de óleo mineral. A PCR foi realizada em termociclador (PTC-100 MJ Research Inc. Minnesota, USA) seguindo programa de amplificação: desnaturação inicial de 95° (5min), seguida de dois ciclos térmicos para anelamento, alongamento e desnaturação respectivamente a 30°C (2min), 72°C (1min) e 95°C (30s) e 33 ciclos com temperatura de anelamento a 40°C e extensão final a 72°C por 5min. As sequências e o número de oligonucleotídeos (Sigma) estão listados na tabela 1.
TABELA 1
Iniciadores aleatórios utilizados nas reações de RAPD-PCR
Iniciadores Sequências Nt % de G
+ C
Referência
L-15996 5’CTCCACCATTAGCACCCAAAGC 3’ 22 54.5 AlphaDNA
Montreal
M13-40 (5’-GTTTTCCCAGTCACGAC-3’) 17 52,9 Idem
GT11-F (5’-GACTCCTGGAGCCCG-3’) 15 73,3 Idem
Nt=nucleotídeos; %=percentagem; G=Guanina e C=citosina
3.5 Eletroforese em gel de poliacrilamida
3.6 Análise dos dados e construção dos fenogramas
Os perfis de bandas obtidas por eletroforese em géis de poliacrilamida foram analisados visualmente. Os padrões observados foram inseridos manualmente no computador e submetidos à análise de proporção de bandas compartilhadas entre as cepas do programa DNA-POP (PENA & NUNES, 1990) e construção de fenogramas pelo programa Treecon for
Windows (VAN der PEER & Der WACHTER, 1994). O programa DNA-POP usa como
princípio básico a divisão vertical do gel em níveis de mobilidade, no qual um nível é definido pela presença de, pelo menos, uma banda em qualquer uma das canaletas do gel. Para cada gel, a análise foi limitada a fragmentos de DNA entre 250 a 3000 pares de bases. De acordo com o número da canaleta e as posições das diferentes bandas foi calculada a proporção de compartilhamento entre as mesmas. O programa analisa os dados em comparações de pares, fornecendo o número total de bandas e a proporção de bandas compartilhadas por cada par e, no final, para todos os pares. A fórmula do coeficiente de similaridade de NEI & LI (1979) foi utilizada para quantificar os polimorfismos de DNA detectados. Para o cálculo da distancia genética, o parâmetro DGxy= 1-[2nxy/(nx +ny)] foi empregado o qual representa a unidade menos o coeficiente de similaridade (NEI & LI, 1979), onde nxy é o número de bandas comuns aos isolados X e Y, e nx e ny são o número de bandas especificas ao perfil de cada um dos isolados. Os fenogramas, representando as matrizes de distância genética, foram construídos pelo método da média aritmética não ponderada- UPGMA (Unweighted Pair Group Method using Arithmetic Average), utilizando-se o programa Treecon for Windows (VAN DER PEER & DER WACHTER, 1994). O grau de reprodutibilidade e confiabilidade dos agrupamentos e das relações taxonônicas obtidas nos fenogramas foi testado pelo Bootstrap, com 1000 reamostragens (FELSENTEIN, 1985).
3.7 Caracterização genética das amostras do T. cruzi
Foram analisadas 27 amostras do T. cruzi correspondentes a 14 indivíduos com sorologia reativa e, 13 de triatomíneos de diferentes espécies (FIG. 1). A tabela 2 mostra a relação das amostras do T. cruzi, os municípios de origem e os hospedeiros dos quais foram isolados.
classificar esses isolados dentro dos grupos genéticos T. cruzi I, II, e III (CÂMARA et al., 2010).
A principal abordagem do presente trabalho foi verificar a variabilidade genética dessas amostras usando a técnica de RAPD. Este marcador também tem sido bastante utilizado na análise de polimorfismo do DNA genômico do T. cruzi e de outros parasitos, tais como,
Giardia, Entamoeba histolytica, Leishmania, Trypanosoma rangeli, Toxoplasma gondii,
Trichomonas vaginalis, demonstrando ser uma ferramenta útil para a análise da variabilidade
genética e capaz de revelar importante informação genética até mesmo entre isolados de uma mesma espécie, além de apresentar boa correlação com a análise isoenzimática (WELSH & MCLELLAND, 1990; WILLIAMS et al., 1990; STEINDEL et al., 1993; TIBAYRENC et al., 1993; OLIVEIRA et al., 1994; GOMES et al., 1995; GOMES et al., 2000; D’ ÁVILA et al.,
2006).
FIGURA 1 – Mapa do Estado do Rio Grande do Norte em destaque os municípios de procedência dos isolados do T. cruzi e localização da Estação Ecológica do Seridó, área de 1.166ha (ESEC, criada pelo decreto 87.222 de 21/05/1982, Lei No. 6.092 de 27/04/1982). Fonte: IBGE mapas
Governador
Dix-Sept
Rosado
TABELA 2
Características dos isolados do Trypanosoma cruzi obtidos de indivíduos e de triatomíneos naturalmente infectados
AMOSTRAS HOSPEDEIROS ORIGEM
T cruzi I - -
RN22-985 Humano Caraúbas
RN28-1150 Humano Angicos
RN29-1816 Humano Caicó
T. cruzi II
RN01-TbN T. brasiliensis Caicó
RN03-TbA T. brasiliensis Serra Negra do Norte RN06-TbN T. brasiliensis Serra Negra do Norte
RN08-462 Humano Serra Negra do Norte
RN13-611 Humano Caraúbas
RN14-838 Humano Caraúbas
RN15-722 Humano Caraúbas
RN16-804 Humano Caraúbas
RN17-792 Humano Caraúbas
RN20-745 Humano Caraúbas
RN21-782 Humano Caraúbas
RN23-462 Humano Serra Negra do Norte
RN25-120 Humano Caicó
RN27-1317 Humano Gov. Dix SeptRosado
T. cruzi III - -
RN02-TbA T. brasiliensis Serra Negra do Norte RN04-TbN T. brasiliensis Serra Negra do Norte RN05-TbN T. brasiliensis Serra Negra do Norte
RN07-PlA P. lutzi Serra Negra do Norte
RN09-PlA P. lutzi Serra Negra do Norte
RN10-PlA P. lutzi Serra Negra do Norte
RN11-PlA P. lutzi Serra Negra do Norte
RN12-PlA P. lutzi Serra Negra do Norte
RN18-PlA P. lutzi Caraúbas
RN19-PlA P. lutzi Caraúbas
NOTA: Código das amostras do T. cruzi e abreviaturas: RN (Rio Grande do
4 RESULTADOS E DISCUSSÃO
4.1 Soroprevalência da infecção pelo T. cruzi
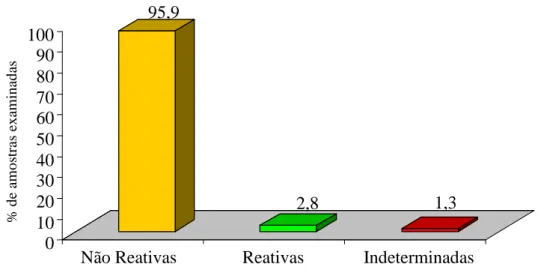
Dos 390 indivíduos, moradores de 28 comunidades rurais no município de Caicó, RN, 11 foram reativos sorologicamente para o T. cruzi pelos métodos de ELISA e RIFI demonstrando 2,8% de soroprevalência da infecção. Em cinco casos os resultados da sorologia foram considerados indeterminados (1,3%) com reatividade obtida por apenas um dos métodos sorológicos e 95,9% não demonstraram reatividade (GRAF. 1). Nas 28 comunidades analisadas, a prevalência da infecção pelo T. cruzi variou de 0,25 a 0,8% (TAB. 3).
GRÁFICO 1 - Percentual de amostras soro reativas, não reativas e indeterminadas dos moradores de diferentes comunidades rurais do município de Caicó, RN
95,9
2,8 1,3
0 10 20 30 40 50 60 70 80 90 100
Não Reativas Reativas Indeterminadas
% d
e
am
o
str
as e
x
am
in
ad
TABELA 3
Reatividade sorológica da infecção pelo Trypanosoma cruzi em moradores das diferentes comunidades rurais do município de Caicó, RN
Comunidades Sorologia não
Reativa
Sorologia
Reativa
Sorologia
indeterminada
Total
Nº % Nº % Nº % Nº %
Alagoinha 4 1,0 - - - - 4 1,0
Alegre de Duda 6 1,5 - - - - 6 1,5
Barra de cima 54 13,9 2 0,5 2 0,5 58 14,9
Barra de Clemente 7 1,8 - - - - 7 1,8
Barra Verde 8 2,05 - - - - 8 2,0
Bela Flor 14 3,6 1 0,3 - - 15 3,9
Dominga 25 6,4 - - - - 25 6,4
Dominga de
Sr.Paulo 5 1,3 - - - - 5 1,3
Esperança 6 1,5 - - - - 6 1,5
Extrema 3 0,8 - - - - 3 0,8
Furna da Onça 13 3,3 - - - - 13 3,3
Inês Velha 2 0,5 - - - - 2 0,5
Lajinha 116 29,7 1 0,25 2 0,5 119 30,5
Monte Alegre 4 1,0 - - - - 4 1,0
Morada Nova 13 3,4 1 0,25 1 0,25 15 3,9
Mundo Novo 4 1,0 - - - - 4 1,0
Nova Olinda 1 0,25 - - - - 1 0,3
Pai Bastião 2 0,5 1 0,25 - - 3 0,8
Recanto 1 0,25 - - - - 1 0,3
Retiro 22 5,6 3 0,8 - - 25 6,5
Riacho 2 0,5 - - - - 2 0,5
Riacho do Fundo 3 0,8 - - - - 3 0,8
Riacho do Meio 4 1,0 1 0,25 - - 5 1,3
Riacho Fundo 3 0,75 - - - - 3 0,8
Saco do Miguel 28 7,2 - - - - 28 7,2
Salgado 4 1,0 - - - - 4 1,0
São Joaquim 1 0,25 - - - - 1 0,3
Varjota 19 4,8 1 0,25 - - 20 5,0
TOTAL 374 95,8 11 2,8 5 1,3 390 100
No primeiro inquérito sorológico nacional no município de Caicó foi observada prevalência de 5,0%, e quando comparada a 2,8% demonstrada neste trabalho, revelou a redução de 56%. Mesmo assim, ainda deve ser considerada elevada, uma vez que há mais de duas décadas existe programa de controle ao vetor nessa mesorregião. Sabe-se que o controle da doença de Chagas no Nordeste é difícil, pelo baixo poder aquisitivo da maioria dos indivíduos, pois é nessa região onde se encontram os mais elevados índices de habitações humanas de baixa qualidade adequadas ao abrigo de triatomíneos e, ainda pelo discreto investimento em inseticidas pela descentralização do serviço de saúde e de vigilância sanitária, e no RN, especialmente, pela dificuldade em se combater as espécies T. brasiliensis e ao T. pseudomaculata no domicílio e no peridomicílio (DIAS & COURA, 1997; DIAS et
al., 2000). Vários autores destacaram que as habitações humanas de baixa qualidade é um dos
fatores de riscos mais relevante em se adquirir a infecção pelo T. cruzi, uma vez que nessas moradias a temperatura e umidade são semelhantes ao do ambiente silvestre, facilitando a adaptação dos triatomíneos. Geralmente nessas moradias ocorre superlotação de indivíduos representando risco para a transmissão vetorial, com elevada concentração de alimento disponível para os triatomíneos em um único local (HOYOS et al., 2007). BORGES-PEREIRA et al. (2008) relataram que a maioria das moradias construídas em áreas rurais, com o desmatamento, parece ser mais favorável à adaptação de triatomíneos, que a usam como habitat e os moradores como fonte de alimentação.
Outros estudos também revelaram prevalências da infecção pelo T. cruzi diferentes das obtidas no inquérito nacional da doença de Chagas realizado no período de 1975 e 1980, destacando a necessidade de atualizar as investigações. No Piauí, estudando 36.399 indivíduos foi constatada uma reatividade de 1,9%, mostrando uma diminuição da infecção quando comparada ao inquérito nacional, que a prevalência foi de 4,04% (BORGES-PEREIRA et al., 2006). No Estado do Ceará, em um inquérito com 541 indivíduos a prevalência foi de 3,1% mostrando um aumento quando comparada a anterior obtida no inquérito nacional de 0,84% (BORGES et al., 2008).
doença de Chaga crônica podem estar relacionados com a diversidade e/ou complexidade genética do T. cruzi que está infectando o hospedeiro, podendo não ser detectado pelos antígenos utilizados no método de diagnóstico em uso, ou resposta imune do indivíduo infectado que determina redução na parasitemia, dificultando a positividade da sorologia (CAMARGO, 1992). Outra possibilidade seria a ocorrência de reação falso-positiva, no caso de soros de pacientes portadores de outras parasitoses tais como, a infecção por T. rangeli, leishnanioses visceral e tegumentar, sífilis, toxoplasmose, hepatite, lupus eritematoso sistêmico e mononucleose (ARAUJO, 1986).
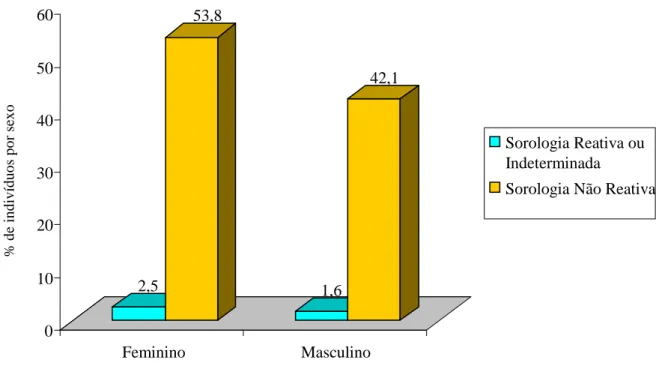
Com a finalidade de traçar um perfil epidemiológico da população dessas comunidades, para avaliar os determinantes da infecção pelo T. cruzi, foram observadas as seguintes características epidemiológicas: gênero, idade, grau de instrução, ocorrência de transfusão de sangue e conhecimento do triatomíneo (ANEXO 5). A maioria dos indivíduos é do gênero feminino, sendo que 2,5% (10/390) deles com sorologia reativa ou indeterminada e 53,8% (210/390) com sorologia não reativa. Comparando-se os dois grupos (reativa ou indeterminada e não reativa) pelo teste do Qui-quadrado em nível de significância de 5% não houve diferença significativa quanto ao gênero no presente trabalho (p= 0,318) (GRAF. 2).
Em áreas rurais, nas quais a transmissão vetorial ocorre ou ocorreu no intradomicílio, foi identificada elevada soroprevalência da infecção pelo T. cruzi no gênero feminino (BORGES-PEREIRA et al., 2008). Isto tem sido atribuído a um período mais longo de permanência das mulheres no interior dos domicílios, indicando que a transmissão vetorial do
T. cruzi tenha ocorrido principalmente no interior das casas (BORGES-PEREIRA et al.,
GRÁFICO 2 - Percentual de indivíduos com sorologia reativa ou indeterminada e não reativa por gênero das diferentes comunidades rurais do município de Caicó, RN.
A idade dos indivíduos com sorologia reativa ou indeterminada variou de 22 a 82 anos, média de 51,7 anos e desvio padrão de 19,8, e naqueles com sorologia não reativa a variação foi de 6 a 89 anos, média de 39,8 anos e desvio padrão de 15,6 (TAB. 4). Comparando os dois grupos de indivíduos pelo teste estatístico T-student em nível de significância de 5% obtivemos diferença significativa (p=0,018). Esta diferença significativa quanto à idade permitiu uma correlação direta entre a incidência da infecção e o tempo de vida do indivíduo, demonstrando o tempo mais longo de exposição desses indivíduos a uma possível contaminação com o T. cruzi. Esse achado também foi observado nos estudos realizados em diversas áreas de Minas Gerais, como, Iguatama e Virgem da Lapa (COURA et al., 1985; BORGES-PEREIRA & COURA, 1986). Em Serra Azul, centro oeste de Minas Gerais, a prevalência da infecção chagásica foi 2,1% em 675 indivíduos avaliados mostrando predominância em indivíduos com faixa etária mais avançada, sendo a média de idade de 67 anos (SILVA et al., 2010). A presença do T. cruzi apenas em indivíduos com idade mais avançada sugere que o controle da transmissão vetorial do parasito nessa área foi bem sucedido. Vale destacar que no presente trabalho não foi detectado nenhum caso de sorologia reativa em crianças e adolescentes (TAB. 4), o que corrobora com os dados do inquérito
2,5 53,8 1,6 42,1 0 10 20 30 40 50 60 Feminino Masculino
Sorologia Reativa ou Indeterminada
Sorologia Não Reativa
sorológico realizado em escolares de 7 a 14 anos de idade (SILVEIRA & VINHAES, 1998). Contudo, as medidas de controle aos triatomíneos devem ser contínuas para evitar contato dos moradores da área rural com os transmissores do agente etiológico no peri e intradomicílio.
TABELA 4
Reatividade sorológica para anticorpos anti-T. cruzi por faixa etária dos 390 moradores das comunidades rurais no município de Caicó, RN
Faixa
Etária
(anos)
Sorologia Não
Reativa
Sorologia
Reativa
Sorologia
Indeterminada
Total
Nº % Nº % Nº % Nº %
0-10 15 3,8 - - - - 15 3,8
11-20 61 16,6 - - - - 61 15,6
21-30 60 15,4 1 0,25 - - 61 15,7
31-40 70 18,0 4 1,0 - - 74 19,0
41-50 60 15,4 2 0,50 1 0,3 63 16,2
51-60 40 10,3 3 0,75 - - 43 11,0
> 60 68 17,4 1 0,25 4 1,0 73 18,7
TOTAL 374 95,9 11 2,8 5 1,3 390 100
N=número de amostras; %=percentual
TABELA 5
Reatividade sorológica para anticorpos anti-T. cruzi por escolaridade dos 390 moradores das diferentes comunidades rurais do município de Caicó, RN
Grau de
Escolaridade
Sorologia Não
Reativa
Sorologia
Reativa
Sorologia
Indeterminada
Total
Nº % Nº % Nº % Nº %
Analfabetos 62 15,9 3 0,75 3 0,8 68 17,5
Fundamental
Incompleto 198 50,9 6 1,5 2 0,5 206 52,9
Fundamental
Completo 19 4,9 - - - - 19 4,9
Médio Incompleto 49 12,6 - - - - 49 12,6
Médio Completo 36 9,2 2 0,5 - - 38 9,7
Superior
Incompleto 1 0,2 - - - - 1 0,2
Superior Completo 9 2,2 - - - - 9 2,2
TOTAL 374 95,9 11 2,8 5 1,3 390 100
N= número de amostras; %= percentual.
migração cria novos dados epidemiológicos e problemas de saúde pública para os países que receberam os imigrantes infectados. Esses problemas incluem riscos de transfusão e transmissão congênita, bem como a necessidade de cuidados médicos para pacientes chagásicos e controles adicionais sobre os bancos de sangue em países com pouca experiência sobre o assunto (COURA & BORGES-PEREIRA, 2010).
Considerando esses dados verificamos que a infecção pelo T. cruzi no Estado do RN, principalmente no município de Caicó foi mais baixa que àquela estimada previamente na literatura para esse município (CAMARGO et al., 1984). Apesar de a transmissão vetorial parecer sob controle ainda há necessidade de um programa epidemiológico contínuo de controle ao transmissor no intra e peridomicílio pela vigilância sanitária o que propiciaria uma melhor qualidade de vida aos moradores dessa mesorregião.
4.2 Avaliação da variabilidade intra-específica das populações do T.
cruzi pela técnica de RAPD
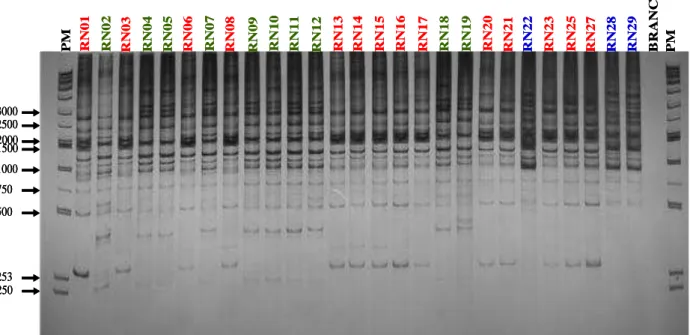
Os perfis de RAPD foram determinados em 27 isolados com os iniciadores aleatórios (L15996, M13-40 e F-λgt11). Os produtos amplificados detectaram semelhanças entre os isolados pertencentes ao mesmo grupo genético do parasito e diferenças entre isolados de grupos genéticos distintos, gerando produtos de DNA de tamanho e intensidade variáveis. Devido à superposição de bandas superior a 3000 pares de base (pb), a análise foi realizada com as bandas que variaram entre 250-3000pb para todos os isolados. A Fig. 2 mostra o gel representativo usando o iniciador M13-40 com elevado compartilhamento de bandas em todos os isolados e entre aqueles representados por uma mesma cor. O compartilhamento de bandas entre todos os isolados foi de 73,7% considerando a média obtida com os três iniciadores e variou de 66% para o iniciador L15996, 75% para o M13-40 e 80% para o gt11-F.
3000 2500 2000 1500 1000 750 253 250 500 RN 04 RN 12 RN 23 B RA NC O PM RN 01 RN 02 RN 03 RN 05 RN 06 RN 07 RN 08 RN 09 RN 10 RN 11 RN 13 RN 14 RN 15 RN 16 RN 17 RN 18 RN 19 RN 20 RN 21 RN 22 RN 25 RN 27 RN 28 RN 29 PM 3000 3000 2500 2500 2000 2000 1500 1500 1000 1000 750 750 253 253 250 250 500 500 RN 04 RN 12 RN 23 B RA NC O PM RN 01 RN 02 RN 03 RN 05 RN 06 RN 07 RN 08 RN 09 RN 10 RN 11 RN 13 RN 14 RN 15 RN 16 RN 17 RN 18 RN 19 RN 20 RN 21 RN 22 RN 25 RN 27 RN 28 RN 29 PM
FIGURA 2 - Gel poliacrilamida a 5% representativo dos perfis de RAPD do T. cruzi, obtidos de indivíduos e triatomíneos naturalmente infectados, com o iniciador M13-40. Nas canaletas das extremidades, o marcador de peso molecular (PM), e na penúltima canaleta, controle negativo da PCR (Branco). As cores representam grupos de isolados com elevado compartilhamento de bandas entre eles.
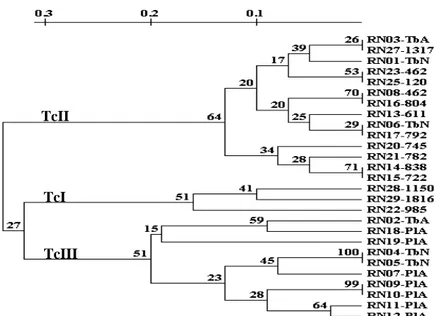
A análise da distância genética obtida com o programa Treecon correspondente aos isolados do T. cruzi utilizando o iniciador M13-40, evidenciou separação desses isolados em três braços principais (FIG. 3). Esses dados demonstraram que os isolados pertencentes ao mesmo grupo apresentam perfil genético ainda mais similar. O primeiro braço separou amostras obtidas de triatomíneos e de humanos oriundas de quatro municípios distintos (Caicó, Caraúbas, Serra Negra do Norte e Governador Dix-Sept Rosado), destacando que os isolados obtidos de hospedeiros e áreas geográficas diferentes foram geneticamente bem correlacionados. O segundo braço do fenograma reuniu amostras isoladas apenas de seres humanos procedentes dos municípios de Caraúbas, Angicos e Caicó. O terceiro braço separou os demais isolados obtidos de triatomíneos de duas espécies diferentes (T. brasiliensis e P.
FIGURA 3 - Fenograma correspondente aos perfis de RAPD de isolados do T. cruzi obtidos de indivíduos e triatomíneos naturalmente infectados, construído pela UPGMA, usando matriz de distância genética obtida com o iniciador M13-40. As percentagens correspondem aos valores bootstrap (percentagens de vezes em que o agrupamento se manteve durante 1000 re-amostragens).
Os dados obtidos com o programa DNA POP para o iniciador M13-40 (ANEXO 7) corroboram com o agrupamento dos isolados obtidos com o programa Treecon, pois a similaridade genotípica foi evidenciada entre os isolados RN14 e RN15, obtidos de indivíduos procedentes do município de Caraúbas localizado na mesorregião oeste, com 100% de compartilhamento de bandas e pertencentes a um mesmo braço no fenograma. É interessante destacar que esse elevado compartilhamento de bandas também ocorre entre os isolados obtidos de diferentes hospedeiros procedentes de mesorregiões distintas, como é o caso das amostras RN01 e RN16, isoladas de T. brasiliensis e de indivíduo com sorologia reativa procedentes de Caicó (mesorregião central) e Caraúbas (mesorregião oeste) com 84% de bandas compartilhadas. No segundo braço o elevado compartilhamento de bandas pode ser destacado entre os isolados RN28 e RN29 com 95% de homogeneidade. Essas amostras foram obtidas de humanos em diferentes municípios (Angicos e Caicó) e são geneticamente bem correlacionadas. No terceiro braço, a homogeneidade das amostras foi evidenciada principalmente pelas amostras RN02 e RN05 isoladas do T. brasiliensis e procedentes de Serra Negra do Norte (mesorregião central) que compartilham 73% de bandas. O padrão elevado de compartilhamento de bandas (74%) entre os isolados RN02 e RN18 obtidos de diferentes espécies de triatomíneos (T. brasiliensis e P. lutzi) capturadas nos municípios Serra Negra do Norte e Caraúbas, localizados em mesorregiões distintas também pode ser
TcII
TcI
observado nesse braço. A comparação entre os isolados pertencentes a distintos braços do fenograma mostrou uma baixa similaridade. Como exemplo, os isolados RN14 e RN19 apenas com 58% de compartilhamento de bandas (FIG. 3). Esses dados demonstraram claramente que todos os isolados são geneticamente bem correlacionados, com similaridade genética ainda mais elevada entre isolados pertencentes ao mesmo grupo genético do T. cruzi e correspondente ao mesmo braço do fenograma.
O programa DNA POP mostra ainda que para esse iniciador o compartilhamento de bandas entre os isolados obtidos de humanos foi de 78,4% e entre os obtidos de triatomíneos foi de 79,3%, sendo 72,8% de compartilhamento entre isolados obtidos do T. brasiliensis e 85,3% entre isolados do P. lutzi.
A análise de isolados correspondente as distintas populações do T. cruzi e com similaridade genotípica tem sido relatado anteriormente por diferentes autores. Zalloum et al. (2005) verificaram elevado compartilhamento de bandas entre os isolados do T. cruzi dos grupos TcI e TcII obtidos de Didelphis marsupialis, triatomíneos e indivíduos infectados no Estado do Paraná. Esses autores sugeriram a presença de uma população do parasito TcI com perfil genético similar circulando entre vetores e reservatórios silvestres e a população do TcII entre humanos. Nesta mesma área, a análise de isolados do T. cruzi obtidos de reservatórios silvestres e de triatomíneos por eletroforese de isoenzimas demonstrou variabilidade clonal limitada sugerindo uma origem ancestral recente dessas populações do T. cruzi (SOCCOL et
al., 2002). As amostras de pacientes chagásicos crônicos provenientes de diferentes áreas
geográficas do Brasil e analisadas por RAPD demonstraram 71% de bandas compartilhadas entre todos os isolados, indicando claramente a presença de um grupo bastante correlacionado (GOMES et al., 1998). No entanto, na Amazônia, cepas obtidas de indivíduos infectados por
via oral e na fase aguda, de primatas e do Rhodnius sp analisadas por RAPD e outros marcadores moleculares, demonstraram similaridade entre as amostras, possivelmente atribuída à mesma linhagem do T. cruzi. Evidenciando que essas populações são transmitidas pelo mesmo vetor na região amazônica e, circulam em ciclos de transmissão independentes determinadas pelos ecótopos específicos de seus hospedeiros vertebrados e invertebrados (MARCILI et al., 2009).
Neste trabalho, a caracterização das populações do T. cruzi por RAPD revelou três grupos distintos do parasito, geneticamente correlacionados entre si. Os resultados sugerem que nas comunidades rurais dos municípios circulam três populações do parasito, uma entre triatomíneos e humanos (braço 1 FIG.3), outra entre os humanos (braço 2 FIG. 3) e, ainda uma terceira circulando apenas entre triatomíneos (braço 3 FIG. 3). Uma mesma população do
Luna-Marin et al. (2009) em 16 isolados do parasito no ciclo doméstico de transmissão na Colômbia. Os isolados foram separados em um grupo de amostras obtidas de triatomíneos e de humanos, e também em um segundo grupo de isolados apenas de triatomíneos, havendo heterogeneidade entre as amostras de grupos diferentes, sugerindo a presença de um determinado clone do T. cruzi circulando entre triatomíneos e humanos (LUNA-MARIN et
al., 2009). A população do parasito que circula apenas entre triatomíneos poder ser
caracterizada como mais heterogênea, devido à necessidade desses triatomíneos se alimentarem em hospedeiros de diferentes espécies, muitas vezes já infectados pelo T. cruzi, propiciando a formação de populações heterogêneas do parasito no hospedeiro invertebrado. Os ciclos naturais de transmissão do T. cruzi são abundantes e complexos, onde uma grande diversidade de mamíferos silvestres e vetores invertebrados são infectados por populações distintas do parasito (COURA et al., 2002). A população que se apresenta circulando entre humanos pode estar correlacionada com a teoria do filtro biológico (MACEDO & PENA, 1998). Essa teoria relata que o organismo humano age como um filtro biológico selecionando populações específicas do parasito, principalmente pela ação do sistema imune. Evidências experimentais têm demonstrado que cepas isoladas de pacientes chagásicos crônicos mostram baixa variabilidade genética do que aquelas obtidas de vetores e hospedeiros silvestres (GOMES et al., 1998; OLIVEIRA et al., 1998).
Os dados do presente trabalho, não demonstraram haver correlação entre a distância geográfica e a variabilidade intraespecífica das amostras do parasito. Esses achados também foram descritos em populações silvestres do T. cruzi obtidas de áreas geográficas distintas da América Central e do Sul com perfis genéticos homogêneos (CARRASCO et al., 1996). As cepas do T. cruzi obtidas de gambás, triatomíneos e indivíduos infectados no Estado do Paraná correspondentes a dois grupos diferentes T. cruzi I e T. cruzi II mostraram homologia entre os isolados, mas não houve correlação com a localização geográfica (ZALLOUM et al., 2005). Utilizando o RAPD, 56 cepas obtidas de triatomíneos, humanos e mamíferos silvestres provenientes de oito Estados do México mostraram homogeneidade em 97,6% do isolados correspondentes ao grupo T. cruzi I independente da localização geográfica (BOSSENO et