Correspondence to: Bela Balint, Military Medical Academy, Institute of Transfusiology, Crnotravska 17, 11 040 Belgrade. E-mail:
be-O R I G I N A L A R T I C L E UDC: 615.387:641.437
Microprocessor–controlled vs. “dump–freezing”platelet and
lymphocyte cryopreservation: a quantitative and qualitative
comparative study
Mikroprocesorski kontrolisana kriokonzervacija trombocita
i limfocita nasuprot neprogramiranom zamrzavanju:
kvantitativna i kvalitativna komparativna studija
Bela Balint*/†, Dušan Vucetić*, Biljana Drašković‡, Danilo Vojvodić‡, Goran Brajušković║, Miodrag Čolić‡, Miroljub Trkuljić*
Military Medical Academy, *Institute of Transfusiology, ‡Institute for Medical Research,
║Institute of Pathology, Belgrade;†
Institute for Medical Research,Belgrade
Abstract
Background/Aim. Thermodynamical and cryobiological pa-rameters responsible for cell damages during cryopreservation (cryoinjuries) have not yet been completely explained. Thus, freezing procedures should be revised, exactly optimized to obtain an enhanced structural and functional recovery of fro-zen–thawed cells. The aim of this study was to compare mic-roprocessor–controlled (controlled–rate) with the compensati-on of the released fusicompensati-on heat and “dump–freezing” (unccompensati-on- (uncon-trolled–rate) of the platelet and lymphocyte cryopreservation efficacy. Methods. Platelet quantitative recovery (post–thaw vs. unfrozen cell count), viability (using hypotonic shock res-ponse – HSR), morphological score (PMS), ultrastructural (electron microscopy) properties and expression of different surface antigens were investigated. In lymphocyte setting, cell recovery and viability (using trypan blue exclusion test) as well as functionality (by plant mitogens) were determined. Control-led–rate freezing and uncontrolControl-led–rate cryopreservation were combined with 6% (platelets) and 10% (lymphocytes) dimethyl sulfoxide (DMSO). Results. Platelet recovery and functionality were superior in the controlled–rate system. The majority of surface antigen expression was reduced in both freezing groups vs. unfrozen cells, but GP140/CD62p was significantly higher in controlled–rate vs. uncontrolled–rate setting. Controlled– rate freezing resulted with better lymphocyte recovery and viability (trypan blue–negative cell percentage). In mitogen–in-duced lymphocyte proliferative response no significant inter-group difference (controlled–rate vs. uncontrolled–rate) were found. Conclusion. The data obtained in this study showned the dependence of cell response on the cryopreservation type. Controlled–rate freezing provided a superior platelet quantitative and functional recovery. Lymphocyte recovery and viability were better in the controlled–rate group, although only a minor inter-group difference for cell proliferative response was obtained.
Key words:
cryopreservation; blood platelets; lymphocytes; dimethyl sulfoxide; cell survival.
Abstrakt
Uvod/Cilj. Termodinamski i kriobiološki parametri, odgovorni za oštećenje ćelija do kojih dolazi tokom kriokonzervacije (krio-oštećenja), još uvek nisu sasvim razjašnjeni. Stoga, postupke zamrz-vanja treba tako optimizirati da se poboljša i strukturni i funkcionalni oporavak zamrznuto-razmrznutnih ćelija. Cilj ove studije bio je upo-ređivanje efikasnosti kriokonzervacije trombocita i limfocita prime-nom programirane kriokonzervacije (mikroprocesorom kontrolisana brzina hlađenja), uz kompenzaciju oslobođene toplote fuzije, i nep-rogramirane kriokonzervacije (bez kompjuterske kontrole brzine hlađenja). Metode. Istraživali smo kvantitativni oporavak trombo-cita (broj ćelija posle odmrzavanja nasuporot broju ćelija pre zamrza-vanja), vijabilnost (koristeći reakciju trombocita na hipotoni šok – HAR), morfološki skor trombocita (PMS), ultrastrukturne (elektro-mikroskopski) karakteristike i ekspresiju različitih površinskih antige-na. Kod limfocita odredili smo njihov kvantitativni oporavak i vija-bilnost (primenom testa isključivanja tripan plavog), kao i funkcio-nalnost (stimulacijom pomoću mitogena). Programirana i neprogra-mirana kriokonzervacija su kombinovane sa 6% (trombociti) i 10% (limfociti) dimetil-sulfidom (DMSO). Rezultati. Oporavak trombo-cita bio je bolji pri uslovima kontrolisane brzine. Većina ekspresije površinskih antigena snižena je bila u obe ispitivane grupe u odnosu na nezamrznute ćelije, dok je ekspresija GP140/CD62p bila znatno veća pri uslovima programirane u odnosu na neprogramiranu krio-konzervaciju. Programirana kriokonzervacija je rezultovala boljim oporavkom i vijabilnošću limfocita (procenat tripan plavo – negativ-nih ćelija). Nismo utvrdili značajnu razliku među grupama (progra-mirana vs neprogra(progra-mirana kriokonzervacija) u proliferativnoj reakciji limfocita prilikom ispitivanja reakcije limfocita nakon indukcije mito-genom. Zaključak. Podaci dobijeni u ovoj studiji pokazali su zavis-nost oporavka ćelija od vrste kriokonzervacije. Zamrzavanje pri kontrolisanoj brzini omogućilo je bolji kvantitativni i funkcionalni oporavak trombocita. Oporavak i vijabilnost limfocita bili su bolji u grupi sa programiranom kriokonzervacijom, mada je konstatovana mala razlika u proliferativnoj reakciji ćelija među grupama.
Ključne reči:
Introduction
Current increasing use of different cell–mediated thera-peutic approaches has resulted in increasing needs for both blood cells and operating procedures in order to minimize cell damages during their collection, processing and storage in liquid or frozen state. Freezing of isolated cells by various cryopreservation systems has been investigated for many years as a method of long–term storage at cryogenic (sub-zero) temperatures. Current freezing techniques are devel-oped from the initial report of the cryopreservation of bovine sperm 1. Cryothermal damages (cryoinjuries) may be the re-sult of extensive cell dehydration (“solution effect”) or intra-cellular ice crystallization (“mechanical damage”) and they result in cell destruction 2–8. Different freezing procedures (modifications of the Polge–Smith–Parkers method) are in routine use for hematopoietic stem cell and progenitor, as well as mature blood cell cryopreservation. The ability of human stem cells to survive a freezing–thawing procedure results in marrow repopulation and hematopoietic reconsti-tution after transplantation 9–21. Nowadays numerous freezing techniques are used in the practice of platelet 22–31 and lym-phocyte 32–36 cryopreservation. However, the optimal cryo-preservation strategy with the greatest quantitative and qualitative cell recovery is still unsolved.
In this study, the influence of the different cryopreserva-tion approaches on the platelet and lymphocyte quantitative and functional recover was compared. It is proposed that the use of controlled–rate vs. uncontrolled–rate (“dump–freezing”) procedure will provide a superior cell recovery. Cryopreserva-tion was carried out using our own controlled–rate systems designated for lymphocyte and platelet cryopreservation, as well as by a standard "dump–freezing" technique under opti-mized dymethyl sulfoxide (DMSO) conditions.
Methods
Platelet and lymphocyte collection and testing
The study included 70 whole blood units collected into CPD SAGM plastic bags (Terumo, Japan), designated for platelet (n = 35) or lymphocyte (n = 35) harvest. The primary buffy–coat (BC–I) was obtained by centrifugation (3 890 × g for 10 min) (Hettich, Germany) and processing of whole blood (450±45 ml) collected from donors in steady–state hematopoiesis into a quadruple bag system (Terumo, Japan). Secondary buffy–coat (BC–II) and buffy– coat derived platelet concentrates (BC–PC) was prepared by centrifugation (377 × g for 5 min) and processing from BC–I 12, 28. Lymphocytes were isolated from BC–II by cen-trifugation (320 × g for 35 min) using Ficoll–Paque (Phar-macia Fine Chemicals, Sweden) as a gradient agent. BC– PC samples (2 ml) were dropped by an automatic micropi-pette into thermo–stable plastic tubes for testing (control group) or cryopreservation. Lymphocytes resuspended in RPMI 1640 culture medium (Serva Feinbiochemica, USA) were taken in the same quantities (2 ml) into plastic tubes for in vitro investigations (control group) or cryopreserva-tion 28, 31, 34.
Platelet count was determined by flow cytometry (Technicon, USA). Platelet morphology was examined by a phase–contrast microscope (Polyvar, Austria) 31. The PMS was calculated according to the numerical values (bal-loons = 0; dendrites = 1; spheres = 2; discs = 4) of different platelet shapes 28, 29. The platelet viability was assessed ac-cording to their hypotonic shock response (HSR) as de-scribed earlier 31, 32. Platelet ultrastructure was examined us-ing an electron microscope (Philips, The Netherlands). Platelet surface antigens (GPIb/CD42b, GPIIb–IIIa/CD41, GP140/CD62p, GP53/CD63, and GPIX/CD42a) were ex-amined using specific monoclonal antibodies (CLB, The Netherlands), while afterward cells were analyzed by an Ep-ics XL (Coulter, USA) 25.
Lymphocyte count in the unfrozen (control) and fro-zen/thawed samples was quantified manually by a Neubauer chamber. Cell viability was investigated by the trypan blue exclusion test. Based on the quantity and viability of thawed lymphocytes, post–cryopreservation total trypan blue–nega-tive lymphocyte recovery (TBNCR) was determined. To as-sess functional statement of cryopreserved lymphocytes 0.5 × 106 cells were cultivated with different concentration of plant mitogens, phytohemaglutinin (PHA) (Sigma, USA) and con-canavalin–A (Con–A) (Sigma, USA), in complete RPMI 1640 culture medium in 96–well microplates (Flow, Irvine, UK). Within the last 18 h cells were pulsed with 3H–thymidine (1 μCi/well) (Amersham, UK). Radioactivity was determined on a beta scintillation counter (Beckman, Germany). Data are expressed as counts per minute (CPM) ± SE and the stimula-tion index (SI) 33–37.
Cell cryopreservation
Cell samples (2 ml) were placed into plastic tubes for cryopreservation. Then, an equal volume (2 ml) of DMSO (Sigma, USA), diluted in autologous plasma (platelet setting) or RPMI 1640 culture medium (lymphocyte setting) was added to cell suspensions. DMSO in 6% (for platelets) and 10% final concentration (for lymphocytes) with both con-trolled–rate and unconcon-trolled–rate freezing were combined. Thus, four cryopreservation protocols: platelet and lympho-cyte controlled–rate (Con−Plt and Con−Ly) and uncon-trolled–rate (Unc−Plt and Unc−Lymphocyte) were investi-gated.
min; IV = -1 °C/min, for 35 min and V = -5 °C, for 5 min. Dur-ing the phase transition period (liquid to solid state) from – 5 °C to –15 °C (platelet setting) and from 0 °C to –10 °C (lymphocyte setting) an intensified (double over) cooling rate (2 °C/min) was used due to the compensation of the re-leased fusion heat 10, 28.
After the completion of controlled–rate freezing proce-dures, cells were placed into a mechanical freezer (Eurospi-tal, Italy) at the temperature of -90±5 °C till thawing (one to tree months) and the structural and functional investigations that followed. The frozen cell samples were thawed rapidly in a water bath at 37±3 °C.
Statistical analysis
The results were expressed as a mean value ± standard deviation (SD) or mean value ± standard error (SE). Prolif-erative response of lymphocytes after controlled–rate vs. un-controlled–rate freezing was compared by the General Linear Model method and Paired Samples t–test. The results ob-tained before and after freezing of platelets were compared by Student's t–test, using the Original PC Program. The dif-ferences were considered as significant at p < 0.05.
Results
The results related to platelet and lymphocyte recovery and viability obtained in this study are presented in Table 1.
The intergroup (controlled–rate vs. uncontrolled–rate) differences showed that there was better platelet and
lym-phocyte quantitative recovery (p < 0.05) in the controlled– rate groups. Uncontrolled–rate freezing caused higher platelet HSR reduction with the significant intergroup differences (p < 0.01). Regarding cell viability, there were no significant differences neither between the cryopreserved vs. control group, nor between lymphocytes frozen by both freezing methods. On the contrary, the TBNCR values for lymphocytes were significantly (p < 0.05) higher in the controlled–rate setting.
The PMS values were rapidly reduced after cryopreser-vation, regardless the applied procedure. For controlled–rate cryopreserved platelets, PMS was 81.8±2.8 %, while for those frozen by the uncontrolled–rate technique PMS was 75.7±3.9 % (Table 2).
The analysis of PMS recovery confirmed that the inter-group differences were significant (p < 0.05) in favor of the controlled–rate freezing.
The results of platelet surface marker investigations be-fore and after cryopreservation are presented in Table 3.
GPIb/CD42b expression was reduced, in both freezing groups as compared to the control one, with no significant intergroup (controlled–rate vs. uncontrolled–rate) differ-ences. On the contrary, noticable intergroup differences (p < 0.05) for GP140/CD62p expression were observed, sug-gesting that controlled–rate freezing provided lower platelet activation.
GP53/CD63 showed an increased expression as com-pared to the control group, but with no significant intergroup
Table 1 The recovery and viability of the frozen/thawed cells
P l a t e l e t s L y m p h o c y t e s Pltrecovery PltVB Lyrecovery LyVB LyTBNCR
Con−Plt* 91.0±5.5† 68.0±23.2‡ / / /
Con−Ly* / / 89.7±5.2† 91.6±1.3 82.5±4.5†
Unc−Plt* 86.0±6.5† 51.6±17.5‡ / / /
Unc−Ly* / / 71.2±16.1† 91.9±7.8 65.5±6.4†
Data are expressed as mean ± SD;
*Protocols investigated: Con−Plt and Con−Ly = controlled–rate freezing with 6% and 10% DMSO; Unc−Plt and Unc−Ly = uncontrolled–rate freezing with 6% and 10% DMSO;
Pltrecovery =cryopreserved/unfrozen (control) platelet count;
PltVB =calculated accordingly of the hypotonic shock response – HSR. Lyrecovery =cryopreserved/unfrozen (control) lymphocyte count; LyVB = lymphocyte viability (determined by trypan blue exclusion test); TBNCR = post–cryopreservation total trypan blue–negative lymphocyte recovery; †
Intergroup (controlled–rate vs. uncontrolled–rate) differences (p < 0.05); ‡
Intergroup (controlled–rate vs. uncontrolled–rate) differences (p < 0.01).
Table 2 Plateletsmorphological score values
Absolute PMS Relative PMS[ %] Control group 356.5±10.3 100 Con–Plt* 291.7±7.4†‡ 81.8±2.8†‡ Unc–Plt 269.9 ± 10.3† 75.7±3.9† Data are expressed as mean ± SD;
*Protocols investigated: Con−Plt = controlled–rate freezing with 6% DMSO; Unc−Plt = uncontrolled–rate freezing with 6% DMSO; †
Cryopreserved vs. unfrozen (control) (p < 0.01);
differences. GPIIb–IIIa/CD41 values were lower than in the control, but also without the significant intergroup differ-ences.
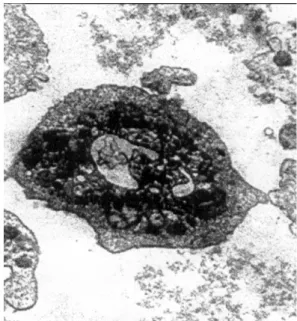
Figure 1a shows the normal unfrozen discoid platelet (control) with the typical ultrastructural properties. The elec-tron microscopy appearance of platelets cryopreserved by controlled–rate and uncontrolled–rate freezing are also pre-sented in Figure 1b–e.
Fig. 1a – The ultrastructural (electron microscopy) prop-erties of cryopreserved platelets
Unfrozen platelet.
Besides discoid and spheric shapes, there were several dendritic platelets, in the controlled-rate freezing group (Fig-ure 1b and 1c). The discoid platelets had maintained intact
microtubules and an open canalicular system with a minimal damage of the membrane. Platelets frozen by the uncon-trolled–rate procedure (Figures 1d and 1e) showed the
mem-brane damages and altered ultrastructural properties (den-dritic or balloon shapes). In addition, these platelets regularly had an unclear edge with numerous large pseudopodes.
Table 3 Surface antigen expression of cryopreserved platelets
CD markers Control Con−Plt* Unc−Plt*
GPIb/CD42b [%] 97.22 ± 2.6 83.2 ± 3.6† 86.08 ± 4.4†
GPIIb–IIIa/CD41 [%] 98.45 ± 0.6 97.47 ± 1.3 96.16 ± 0.8 ‡
GP140/CD62p [%] 8.88 ± 3.8 28.38 ± 5.2† 34.91 ± 8.3†‡
GP53/CD63 [%] 0.79 ± 0.2 1.96 ± 0.9† 2.08 ± 1.2†
*Protocols investigated: Con−Plt = controlled–rate freezing with 6% DMSO; Unc−Plt = uncontrolled–rate freezing with 6% DMSO; †
Cryopreserved vs. unfrozen (control) (p < 0.05);
‡Intergroup (controlled–rate vs. uncontrolled–rate) differences (p < 0.05).
Fig. 1c – The ultrastructural (electron microscopy) properties of cryopreserved platelets
Controlled–rate cryopreserved platelets.
Fig. 1d – The ultrastructural (electron microscopy) properties of cryopreserved platelets
Uncontrolled–rate cryopreserved platelets.
Fig. 1b – The ultrastructural (electron microscopy) properties of cryopreserved platelets
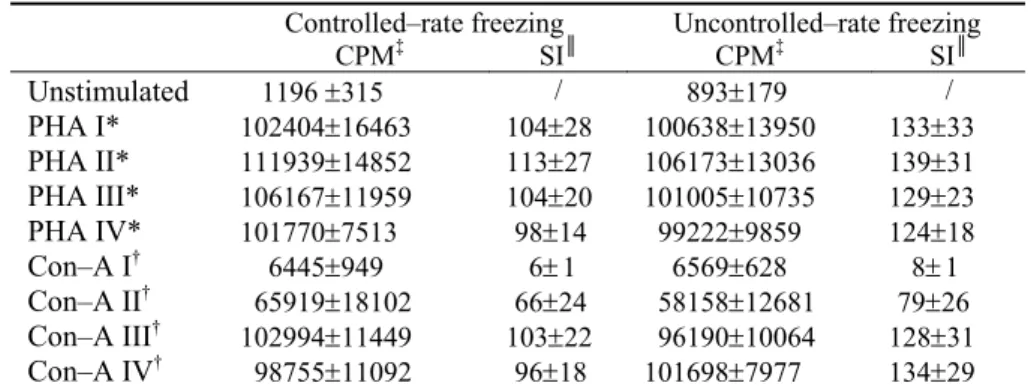
Finally, Table 4 shows the mitogen (PHA and Con–A) induced lymphocyte proliferative response in controlled–rate and uncontrolled–rate setting.
The analysis of the obtained results of the lymphocyte proliferative response demonstrated that the cryopreservation resulted in no significant intergroup differences, regardless the used freezing procedure.
Discussion
Our earlier cryoinvestigations 10, 12, 28 pointed out the im-portance of the kinetics of the programmed cooling for the to-tal nucleated cell (TNC), lymphocyte and platelet cryopreser-vation. In this study, we confirmed that the use of the con-trolled–rate method had significantly influenced the protection of quantity and functionality of frozen/thawed platelets. The platelet postthaw recovery was exactly 91.0±5.5%, which was better than in other studies (62 to 86%) reported 22, 23, 37, 38.
The results obtained in this research showed that the controlled–rate freezing yielded the improved platelet
viabil-ity, as projected across HSR. Although in this study the lower HSR values were observed for both freezing proce-dures (compared to the control one), the 32% reduction in the controlled–rate setting is in accordance with other re-ports. However, HSR value depends on some additional pa-rameters, such as cell platelet age, cell membrane integrity, type of platelet shapes, etc 23, 31, 39. Finally, we believe that HSR of cryopreserved platelets is not always a result of the unsuccessful response of all cells, but could rather represent an unequal response of different platelet shapes.
The examination of PMS also confirmed clearly that controlled–rate freezing was more efficient than the untrolled–rate technique. Better PMS values obtained in the con-trolled–rate setting were primarily due to a greater proportion of platelets of disc shapes (around 70% in comparison with the control). These data generally agree with those reported in the literature on PMS values after freezing–thawing procedure. It is generally accepted that the high–quality in vivo survival of cells can be expected after transfusion if 50% or more of platelets keep their disc shape after cryopreservation 39–41.
The investigation of ultrastructural properties of platelets confirmed the advantage of the controlled–rate freezing, since it revealed that the discoid platelets cryopreserved by this protocol had kept intact microtubules and an open canalicular system, with the minimal membrane changes. The spheric shaped platelets showed the lower electron density of cyto-plasm, gradually changed the interior structure comparing to other shapes, which was manifested by the marginal location of granules and other cytoplasmatic organelles. This finding is in accordance with our previous platelet cryoinvestigation 28. On the contrary, the samples cryopreserved by the uncon-trolled–rate technique showed an abundance of the damaged platelets and cell detritus, along with the lower percentage of platelets with a partially protected membrane integrity. The occurrence of the regular dendritic and balloon shapes, as well as rare spheric platelets indicated reduced morphological properties and the subsequent functionality of thawed cells.
Fig. 1e – The ultrastructural (electron microscopy) properties of cryopreserved platelets
Uncontrolled–rate cryopreserved platelets.
Table 4 Mitogen–induced lymphocyte proliferative response
Controlled–rate freezing Uncontrolled–rate freezing
CPM‡ SI║ CPM‡ SI║
Unstimulated 1196 ±315 / 893±179 /
PHA I* 102404±16463 104±28 100638±13950 133±33
PHA II* 111939±14852 113±27 106173±13036 139±31
PHA III* 106167±11959 104±20 101005±10735 129±23
PHA IV* 101770±7513 98±14 99222±9859 124±18
Con–A I† 6445±949 6±1 6569±628 8±1
Con–A II†
65919±18102 66±24 58158±12681 79±26
Con–A III† 102994±11449 103±22 96190±10064 128±31
Con–A IV†
98755±11092 96±18 101698±7977 134±29
Data are expressed as mean ± SE; (four lymphocyte samples per each group); ‡
CPM = counts per minute; ║SI = stimulation index;
*PHA concentrations used: I = 62.5 μg/ml; II = 31.3 μg/ml; III = 15.6 μg/ml; IV = 7.8 μg/ml;
†
In the present study, GPIIb–IIIa/CD41 platelet surface marker was investigated. In addition, platelet activation was assessed by measuring the expression of GPIb/CD42b, GP53/CD63 and GP140/CD62p (P–selectin). The last one is expressed on the cell surface after the release from α –gran-ules.
It is generally accepted that the cold–induced post– transfusion platelet clearance with following compromised clinical effectiveness should be mediated by GPIb/CD42b reorganization into “clusters” 42–47. Our preliminary cryoin-vestigation (29) of cold–induced GPIb/CD42b alterations was in agreement with these studies, particularly when un-controlled–rate freezing was performed. We proposed that the reduction of the GPIb/CD42b expression in the present study (from 11.1 to 14.0%) might be partially caused by the GPIbα/CD42b “relocation”. However, these values are markedly better than those reported by Lozano (reduction > 50%) 44. The mentioned “relocation” could probably make surface antigens less accessible to monoclonal antibodies, resulting in the obtained decrease of GPIb/CD42b expres-sion the of cryopreserved vs. the control platelets. How-ever, Hoffmeister et al. (personal communication) did not find a reduced GPIbα/CD42b expression following cooling using the monoclonal antibody for platelets, so they con-cluded that clustering did not lead to “pseudo hyporeactiv-ity”. In this context, GPIb/CD42b expression decrease should be mediated by other mechanisms such as proteoly-sis by plasmin, thrombin, neutrophil elastase or the activa-tion of the cell “death machinery”, etc 29, 46. The results obtained in our study are comparable with those published by Barnard et al., who also found a reduced GPIb/CD42b expression after cryopreservation 46. They proposed that decreased antigen expression could be associated with GPIb/CD42b proteolysis, mediated by calcium–activated protease, or calpain.
It is generally accepted that the cold–induced GP140/CD62p expression less than 40% could be an ade-quate threshold, because higher expression values of GP140/CD62p have been shown to result in platelet in vivo recovery of less than 50% 48). As shown in Table 3, freezing promoted a raise in the number of platelets expressing GP140/CD62p antigen. The percentage of platelets express-ing GP140/CD62p in the controlled–rate group was signifi-cantly lower than in the uncontrolled–rate sample. The ele-vated GP140/CD62p values for cryopreserved cells pointed to the platelet activation during the freezing/thawing proce-dure.
The expression of GP140/CD62p was associated with the clearance of platelets by the reticuloendothelial system 48. More than 50% of platelets stored at 22 °C expressed GP140/CD62p on their surface after 5 days, which indicated that the cryopre-served cells had the superior retention of intact granules, which correlated with their in vivo functionality 49. With respect to GP53/CD63 antigen (activation marker of the externalization of the contents from lysosomes), a considerable increase in the expression of this antigen after freezing–thawing process was observed. However, this increase in the GP53/CD63 ex-pression was not as high as for GP140/CD62p antigen. This
finding might reflect the more intensive activator stimulus necessary for the release of lysosomes compared to that of
α–granules 50.
In this study, the GPIIb–IIIa/CD41 (the main platelet aggregation protein) examination showed the decrease in cryopreserved platelets in both groups. However, the reduc-tion of GPIIb–IIIa/CD41 was not significant after con-trolled–rate freezing vs. the control group (opposite to un-controlled–rate freezing), which is in agreement with data from the literature 51, 52. GPIV/CD36 is an accelerator of the interaction between platelets and collagen and it is a media-tor of the platelet adhesion to subendothelial surfaces 53. Cryopreserved platelets in the present investigation obtained a significant reduction of the expression of GPIV/CD36 as compared to unfrozen platelets, but without intergroup dif-ferences. These data are in accordance with data from the lit-erature 44.
For a successful cryopreservation not only the use of appropriate freezing strategy, but also the selection of a cryoprotectant in the optimized concentration is required. For platelet and lymphocyte freezing, DMSO and/or hy-droxyethyl starch (HES) dissolved in a cell–specific “freezing medium” is commonly used, but with different concentrations of cryoprotectant 54, 55. In the present study, 6% DMSO in autologous plasma (platelet setting), and 10% DMSO in RPMI 1640 (lymphocyte setting) were used. Our earlier cryoinvestigation showed that the TNC and the committed progenitor recovery was better when the lower (5%) DMSO and controlled–rate freezing were used. In contrast, pluripotent Makrow Repopulating Ability (MRA) cells could only be well cryopreserved in the presence of 10% DMSO 10. These data implied a different “cryobi-ological request” of MRA cells in comparison to the com-mitted progenitors or TNCs. However, as recently pre-sented, the postthaw cell quality is better when rHu–DNase, membrane stabilizers or bioantioxidants are added to the Conventional Freezing Medium (CFM) 56–58. Accordingly, it is possible that the use of these additives is more success-ful than the reduction of DMSO concentration. We also showed that the differences in the cell recovery and func-tionality rates were not related to the changes in the total number of frozen/thawed cells 10, 11.
Conclusion
In conclusion, the results presented here point undoubt-edly out the controlled–rate system (with the compensated fusion heat) which resulted in a superior platelet recovery of the tested parameters. A simultaneous analysis of platelet surface antigens and cell micro–integrity markers could pre-dict certain in vivo properties of the cooled, especially
cryo-preserved platelets, however these postulates should be con-firmed. The lymphocyte recovery and TBNCR values were obviously lower in the uncontrolled–rate setting. The ob-tained high–level postthaw cell viability can explain their similar proliferative response regardless the used freezing. Further researches in various blood–derived cell settings are recommended to define the optimal cell–specific cryopreser-vation strategy with the best cell recovery.
R E F E R E N C E S
1. Polge C, Smith AU, Parkes AS. Revival of spermatozoa after vit-rification and dehydratition at low temperatures. Nature 1949; 164: 666.
2. Barnes DW, Loutit JF. The radiation recovery factor: preserva-tion by the Polge-Smith-Parkes technique. J Natl Cancer Inst 1955; 15(4): 901–5.
3. Lovelock JE, Bishop MW. Prevention of freezing damage to living cells by dimethyl sulphoxide. Nature 1959; 183(4672): 1394–5. 4. Mollison PL, Sloviter HA. Successful transfusion of previously
frozen human red cells. Lancet 1951; 2(19): 862–4.
5. Rowe AW, Rinfret AP. Controlled rate freezing of bone mar-row. Blood 1962; 20: 636.
6. Rowe AW. Biochemical aspects of cryoprotective agents in freezing and thawing. Cryobiology 1966; 3(1): 12–8.
7. Mazur P. Theoretical and experimental effects of cooling and warming velocity on the survival of frozen and thawed cells. Cryobiology 1966; 2(4): 181–92.
8. Meryman HT. Cryoprotective agents. Cryobiology 1971; 8(2): 173–83.
9. Gorin NC. Collection, manipulation and freezing of haemopoi-etic stem cells. Clin Haematol 1986; 15(1): 19–48.
10. Balint B, Ivanović Z, Petakov M, Taseski J, Jovčić G, Stojanović N, et al. The cryopreservation protocol optimal for progenitor re-covery is not optimal for preservation of marrow repopulating ability. Bone Marrow Transplant 1999; 23(6): 613–9.
11. Balint B, Taseski J. Cryopreservation of haematopoietic stem and progenitor cells. Makedon Med Pregl 2000; 54: 80–4. 12. Balint B, Ivanović Z, Petakov M, Taseski J, Vojvodić D, Jovčić G, et
al. Evaluation of cryopreserved murine and human hemato-poietic stem and progenitor cells designated for transplanta-tion. Vojnosanit Pregl 1999; 56(6): 577–85.
13. Balint B. Stem cells – unselected or selected, unfrozen or cryo-preserved: marrow repopulation capacity and plasticity poten-tial in experimental and clinical settings. Makedon Med Pregl 2004; 58(Suppl 63): 22–4.
14. Balint B. Coexistent cryopreservation strategies: microproces-sor-restricted vs. uncontrolled-rate freezing of the "blood-derived" progenitors/cells. Blood Banking Transf Med 2004; 2(2): 62–71.
15. Balint B; The National Apheresis Group. Apheresis in donor and therapeutic settings: recruitments vs. possibilities – a mul-ticenter study. Transfus Apher Sci 2005; 33(2): 181–9. 16. Balint B. Stem and progenitor cell harvesting, extracorporeal
"graft engineering" and clinical use – initial expansion vs. cur-rent dillemas. Clin Appl Immunol. 2006. In press.
17. Takaue Y, Abe T, Kawano Y, Suzue T, Saito S, Hirao A, et al. Com-parative analysis of engraftment after cryopreservation of periph-eral blood stem cell autografts by controlled- versus uncontrolled-rate methods. Bone Marrow Transplant 1994; 13(6): 801–4. 18. Perez-Oteyza J, Bornstein R, Corral M, Hermosa V, Alegre A,
Torra-badella M, et al. Controlled-rate versus uncontrolled-rate cryo-preservation of peripheral blood progenitor cells: a prospective multicenter study. Group for Cryobiology and Biology of
Bone Marrow Transplantation (CBTMO), Spain. Haema-tologica 1998; 83(11): 1001–5.
19. Montanari M, Capelli D, Poloni A, Massidda D, Brunori M, Spitaleri L, et al. Long-term hematologic reconstitution after autolo-gous peripheral blood progenitor cell transplantation: a com-parison between controlled-rate freezing and uncontrolled-rate freezing at 80 degrees C. Transfusion 2003; 43(1): 42–9. 20. Stiff PJ, Koester AR, Weidner MK, Dvorak K, Fisher RI.
Autolo-gous bone marrow transplantation using unfractionated cells cryopreserved in dimethylsulfoxide and hydroxyethyl starch without controlled-rate freezing. Blood 1987; 70(4): 974–8. 21. Buchanan SS, Gross SA, Acker JP, Toner M, Carpenter JF, Pyatt
DW. Cryopreservation of stem cells using trehalose: evaluation of the method using a human hematopoietic cell line. Stem Cells Dev 2004; 13(3): 295–305.
22. Kim BK, Baldini MG. Preservation of viable platelets by freez-ing. Effect of plastic containers. Proc Soc Exp Biol Med 1973; 142(1): 345–50.
23. Liu JH, Ouyang XL, Lu LC, Gao D. Thermometry of intracel-lular ice crystal formation in cryopreserved platelets. Zhong-guo Shi Yan Xue Ye Xue Za Zhi 2002; 10(6): 574–6. (Chinese) 24. Angelini A, Dragani A, Berardi A, Iacone A, Fioritoni G, Torlantano G. Evaluation of four different methods for platelet freezing. In vitro and in vivo studies. Vox Sang 1992; 62(3): 146–51. 25. Vadhan-Raj S, Kavanagh JJ, Freedman RS, Folloder J, Currie LM,
Bueso-Ramos C, et al. Safety and efficacy of transfusions of autologous cryopreserved platelets derived from recombinant human thrombopoietin to support chemotherapy-associated severe thrombocytopenia: a randomised cross-over study. Lancet 2002; 359(9324): 2145–52.
26. Landi EP, Roveri EG, Ozelo MC, Annichino-Bizzacchi JM, Origa AF, de Carvalho Reis AR, et al. Effects of high platelet concen-tration in collecting and freezing dry platelets concentrates. Transfus Apher Sci 2004; 30(3): 205–12.
27. Arnaud F, Kapnik E, Meryman HT. Use of hollow fiber mem-brane filtration for the removal of DMSO from platelet con-centrates. Platelets 2003; 14(3): 131–7.
28. Balint B, Vučetić D, Trajković-Lakić Z, Petakov M, Bugarski D, Brajušković G, et al. Quantitative, functional, morphological and ultrastructural recovery of platelets as predictor for cryopre-servation. Haematologia (Budap) 2002; 32(4): 363–75. 29. Balint B, Vučetić D, Vojvodić D, Petakov M, Brajušković G, Ivanović Z,
et al. Cell recovery, cryothermal micro-damages and surface an-tigen expression as predictors for cold-induced GPIbα /CD42b-cluster mediated platelet clearance after controlled-rate vs. un-controlled-rate cryopreservation. Blood Banking Transf Med 2004; 2(1): 22–6.
30. Balint B, Paunović D, Vučetić D, Vojvodić D, Petakov M, Trkuljić M, et al. Controlled-rate vs. uncontrolled-rate freezing as pre-dictors for platelet cryopreservation efficacy. Transfusion 2006; 46: In press.
32. Vučetić D, Balint B, Trajković-Lakić Z, Mandić-Radić S, Brajušković G, Miković D, et al. Cryopreserved vs. liquid-state stored plate-lets – quantitative and qualitative comparative stud. Blood Banking Transfus Med 2003; 1: 81–8.
33. Miniscalco B, D'Angelo A, Cagnasso A. Effect of storage and cryopreservation on the lymphocyte responses to polyclonal mitogens in cattle. Vet Res Commun 2003; 27 Suppl 1: 775–8. 34. Costantini A, Mancini S, Giuliodoro S, Butini L, Regnery CM,
Sil-vestri G, et al. Effects of cryopreservation on lymphocyte im-munophenotype and function. J Immunol Methods 2003; 278(1–2): 145–55.
35. Petakov M, Balint B, Bugarski D, Jovčić G, Stojanović N, Vojvodić D, et al. Donor leukocyte infusion--the effect of mutual reactivity of donor's and recipient's peripheral blood mononuclear cells on hematopoietic progenitor cells growth. Vojnosanit Pregl 2000; 57(5): 89–93.
36. Sohn SK, Jung JT, Kim DH, Lee NY, Seo KW, Chae YS, et al. Prophylactic growth factor-primed donor lymphocyte infu-sion using cells reserved at the time of transplantation after allogeneic peripheral blood stem cell transplantation in pa-tients with high-risk hematologic malignancies. Cancer 2002; 94(1): 18–24.
37. Knight SC, Farrant J. Comparing stimulation of lymphocytes in different samples: separate effects of numbers of responding cells and their capacity to respond. J Immunol Methods 1978; 22(1–2): 63–71.
38. Balduini CL, Mazzucco M, Sinigaglia F, Grignani G, Bertolino G, Noris P, et al. Cryopreservation of human platelets using di-methyl sulfoxide and glycerol-glucose: effects on "in vitro" platelet function. Haematologica 1993; 78(2): 101–4.
39. Vadhan-Raj S, Currie LM, Bueso-Ramos C, Livesey SA, Connor J. Enhanced retention of in vitro functional activity of platelets from recombinant human thrombopoietin-treated patients following long-term cryopreservation with a platelet-preserving solution (ThromboSol) and 2% DMSO. Br J Hae-matol 1999; 104(2): 403–11.
40. Schiffer CA, Buchholz DH, Aisner J, Wolff JH, Wiernik PH. Fro-zen autologous platelets in the supportive care of patients with leukemia. Transfusion 1976; 16(4): 321–9.
41. Melaragno AJ, Carciero R, Feingold H, Talarico L, Weintraub L, Valeri CR. Cryopreservation of human platelets using 6% di-methyl sulfoxide and storage at -80 degrees C. Effects of 2 years of frozen storage at -80 degrees C and transportation in dry ice. Vox Sang 1985; 49(4): 245–58.
42. Hoffmeister KM, Felbinger TW, Falet H, Denis CV, Bergmeier W, Mayadas TN, et al. The clearance mechanism of chilled blood platelets. Cell 2003; 112(1): 87–97.
43. Hoffmeister KM, Josefsson EC, Isaac NA, Clausen H, Hartwig JH, Stossel TP. Glycosylation restores survival of chilled blood platelets. Science 2003; 301(5639): 1531–4.
44. Lozano M, Escolar G, Mazzara R, Connor J, White JG, DeLecea C, et al. Effects of the addition of second-messenger effectors to platelet concentrates separated from whole-blood donations and stored at 4 degrees C or -80 degrees C. Transfusion 2000; 40(5): 527–34.
45. Bode AP, Knupp CL, Miller DT. Effect of platelet activation in-hibitors on the loss of glycoprotein Ib during storage of plate-let concentrates. J Lab Clin Med 1990; 115(6): 669–79. 46. Barnard MR, MacGregor H, Ragno G, Pivacek LE, Khuri SF,
Mich-elson AD, ‚et al. Fresh, liquid-preserved, and cryopreserved platelets: adhesive surface receptors and membrane procoagu-lant activity. Transfusion 1999; 39(8): 880–8.
47. Rinder HM, Murphy M, Mitchell JG, Stocks J, Ault KA, Hillman RS. Progressive platelet activation with storage: evidence for shortened survival of activated platelets after transfusion. Transfusion 1991; 31(5): 409–14.
48. Hamburger SA, McEver RP. GMP-140 mediates adhesion of stimulated platelets to neutrophils. Blood 1990; 75(3): 550–4. 49. Connor J, Currie LM, Allan H, Livesey SA. Recovery of in vitro
functional activity of platelet concentrates stored at 4 degrees C and treated with second-messenger effectors. Transfusion 1996; 36(8): 691–8.
50. Holmsen H. Platelet metabolism and activation. Semin Hematol 1985; 22(3): 219–40.
51. Owens M, Werner E, Holme S, Afflerbach C. Membrane glycopro-teins in cryopreserved platelets. Vox Sang 1994; 67(1): 28–31. 52. Lozano ML, Rivera J, Corral J, Gonzalez-Conejero R, Vicente V.
Platelet cryopreservation using a reduced dimethyl sulfoxide concentration and second-messenger effectors as cryopre-serving solution. Cryobiology 1999; 39(1): 1–12.
53. Diaz-Ricart M, Tandon NN, Gomez-Ortiz G, Carretero M, Escolar G, Ordinas A, et al. Antibodies to CD36 (GPIV) inhibit platelet adhesion to subendothelial surfaces under flow conditions. Arterioscler Thromb Vasc Biol 1996; 16(7): 883–8.
54. Lakota J, Fuchsberger P. Autologous stem cell transplantation with stem cells preserved in the presence of 4.5 and 2.2% DMSO. Bone Marrow Transplant 1996; 18(1): 262–3. 55. Halle P, Tournilhac O, Knopinska-Posluszny W, Kanold J, Gembara
P, Boiret N, et al. Uncontrolled-rate freezing and storage at -80 degrees C, with only 3.5-percent DMSO in cryoprotective so-lution for 109 autologous peripheral blood progenitor cell transplantations. Transfusion 2001; 41(5): 667–73.
56. Beck C, Nguyen XD, Kluter H, Eichler H. Effect of recombinant human deoxyribonuclease on the expression of cell adhesion molecules of thawed and processed cord blood hematopoietic progenitors. Eur J Haematol 2003; 70(3): 136–42.
57. Limaye LS, Kale VP. Cryopreservation of human hematopoietic cells with membrane stabilizers and bioantioxidants as addi-tives in the conventional freezing medium. J Hematother Stem Cell Res 2001; 10(5): 709–18.
58. Dijkstra-Tiekstra MJ, de Korte D, Pietersz RN, Reesink HW, van der Meer PF, Verhoeven AJ. Comparison of various dimethylsul-phoxide-containing solutions for cryopreservation of leu-coreduced platelet concentrates. Vox Sang 2003; 85(4): 276–82. 59. Moroff G, Seetharaman S, Kurtz JW, Greco NJ, Mullen MD, Lane TA,
et al. Retention of cellular properties of PBPCs following liquid storage and cryopreservation. Transfusion 2004; 44(2): 245–52. 60. Weinberg A, Zhang L, Brown D, Erice A, Polsky B, Hirsch MS, et
al. Viability and functional activity of cryopreserved mononu-clear cells. Clin Diagn Lab Immunol 2000; 7(4): 714–6.