PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA ELÉTRICA
Alex Martins
SINCRONISMO ENTRE REDES NEURAIS COM TOPOLOGIA DE ACOPLAMENTO DO TIPO NEWMAN-WATTS
Orientador: Prof. Dr. Luiz Henrique Alves Monteiro
Dissertação apresentada ao Programa de Pós-Graduação em Engenharia Elétrica da Universidade Presbiteriana Mackenzie como requisito para obtenção do título de Mestre em Engenharia Elétrica
M386s Martins, Alex.
Sincronismo entre redes neurais com topologia de acoplamento
do tipo Newman-Watts / Alex Martins – 2011.
94 f. : il. ; 30 cm.
Dissertação (Mestrado em Engenharia Elétrica) –
Universidade Presbiteriana Mackenzie, São Paulo, 2011. Bibliografia: f. 92-94.
1. Autômato celular. 2. Neurodinâmica. 3. Redes complexas.
Sincronismo. I. Título.
SINCRONISMO ENTRE REDES NEURAIS COM TOPOLOGIA DE ACOPLAMENTO DO TIPO NEWMAN-WATTS
Dissertação apresentada ao Programa de Pós-Graduação em Engenharia Elétrica da Universidade Presbiteriana Mackenzie como requisito para obtenção do título de Mestre em Engenharia Elétrica
Aprovada em
BANCA EXAMINADORA
Prof. Dr. Luiz Henrique Alves Monteiro - orientador Universidade Presbiteriana Mackenzie
Profa. Dra. Gisele Akemi Oda Universidade de São Paulo
Agradeço ao Luiz, mentor1 deste trabalho; Ao Programa de Pós-graduação em Engenharia Elétrica do Mackenzie, por seu apoio.
Pode-se entender sincronismo como uma organização temporal de eventos, possível de emergir em sistemas complexos, como redes neurais. Aqui, usam-se grafo aleatório e autômato celular (AC) para representar redes neurais, a fim de investigar a ocorrência de sincronismo em tais redes. A topologia de acoplamento da rede é do tipo Newman-Watts, formada por uma grade regular com ligações aleatórias acrescentadas. Duas partes com essa estrutura são conectadas por ligações aleatórias. Resultados obtidos por simulações numéricas com esse modelo indicam diversidade de comportamento oscilatório: há casos em que as duas partes oscilam em períodos iguais, múltiplos e submúltiplos; e casos sem oscilação. Investigaram-se as relações entre comportamento oscilatório e a atividade máxima, o tempo para se alcançar essa atividade, o comprimento do caminho mínimo médio, o tamanho da rede, a porcentagem de ligações aleatórias adicionadas, e as regras de transição de estado do AC. Comportamento síncrono foi encontrado em mais de 75% das 28.000 simulações realizadas. A dinâmica do sistema é mais influenciada por variações no número de passos de tempo em que a célula permanece disparando do que por alterações no tamanho do reticulado ou no percentual das ligações aleatórias adicionais.
Synchronization can be understood as a temporal organization of events, able of emerging in complex systems, as neural networks. Here, random graph and cellular automaton (CA) are used to represent neural networks, in order to investigate the occurrence of synchronism in such networks. The network coupling topology is of Newman-Watts type, formed by regular lattice with additional random connections. Two parts with this structure are connected by random links. Results obtained from numerical simulations with this model indicate variety of oscillatory behavior: there are cases in which both parts oscillate with equal, multiple and submultiple periods; and cases without oscillation. Investigations were performed concerning the relation among oscillatory behavior and maximum activity, the time to reach such an activity, the minimum average path length, size of the network, the percentage of random connections added and the rules of the CA state transition. Synchronous behavior was found in more than 75% of 28000 simulations accomplished. The system dynamics is influenced more by variations on the number of time steps in which a cell remains firing than by alterations on the lattice size or on the percentage of the randomly added links.
Lista de Figuras
Figura 3.1 - Autômato de duas partes, baseado em grafo regular (ligações em preto) com m=10 e n=20, com ligações aleatórias intrapartes (ligações em azul) e entrepartes (ligações em verde). ...29 Figura 4.1 – Topologia da rede 7, que possui 5% de ligações adicionais (N=200). Ligações entre células da mesma parte em azul; entre células de partes diferentes em verde. Os números 1, 2 e 3 identificam as células estimuladas que produziram diferentes comportamentos...48 Figura 4.2– Topologia da rede 19, que possui 5% de ligações adicionais. Ligações entre células da mesma parte em azul; entre células de partes diferentes em verde. Os números 8,9 e 10 identificam as células estimuladas que produziram diferentes comportamentos. ...50
Lista de Gráficos
Gráfico 4.32 – Participação dos tercis dos caminhos mínimos médios das redes nos tipos de oscilação, parte 2. . À esquerda redes com 5%, ao centro redes com 10% e à direita redes com 15% de ligações aleatórias adicionadas (N=200). ...72 Gráfico 4.33 – Comparação dos tipos de oscilação nas diferentes razões entre o período da parte 1 e o da entrada. À esquerda, redes com 5%; ao centro, redes com 10%; e à direita, redes com 15% de ligações aleatórias adicionadas (N=200). ...73 Gráfico 4.34 – Participação dos tercis da atividade máxima da parte 1 nos tipos de oscilação. À esquerda, redes com 5%; ao centro, redes com 10%; e à direita, redes com 15% de ligações aleatórias adicionadas (N=200). ...74 Gráfico 4.35 – Participação dos tercis da atividade máxima da parte 2 nos tipos de oscilação. À esquerda, redes com 5%; ao centro, redes com 10%; e à direita, redes com 15% de ligações aleatórias adicionadas (N=200). ...75 Gráfico 4.36 – Participação dos tercis dos tempos para atingir a atividade máxima nos tipos de oscilação, para a parte 1. À esquerda, redes com 5%; ao centro, redes com 10%; e à direita, redes com 15% de ligações aleatórias adicionadas (N=200).. ...76 Gráfico 4.37 – Participação dos tercis dos tempos para atingir a atividade máxima nos tipos de oscilação, para a parte 2. À esquerda, redes com 5%; ao centro, redes com 10%; e à direita, redes com 15% de ligações aleatórias adicionadas (N=200). ...76 Gráfico. 4.38 – Número de observações em que o período de oscilação da parte 2 em relação ao período da parte 1, por período da entrada, para as regra 2 (acima) e para a regra 3 (abaixo) (N=200, q=5%). ...79 Gráfico 4.39 – Participação dos tercis da atividade máxima da parte 1 nos tipos de oscilação. À esquerda, a regra 1; ao centro, a regra 2; à direita, a regra 3 (N=200, q=5%). ...81 Gráfico 4.40 – Participação dos tercis da atividade máxima da parte 2 nos tipos de oscilação. À esquerda, a regra 1; ao centro, a regra 2; à direita, a regra 3 (N=200, q=5%). ...82 Gráfico 4.41 – Participação dos tercis dos tempos para atingir a atividade máxima nos tipos de oscilação, para a parte 1. À esquerda, a regra 1; ao centro, a regra 2; à direita, a regra 3 (N=200, q=5%). ...83 Gráfico 4.42 – Participação dos tercis dos tempos para atingir a atividade máxima nos tipos de oscilação, para a parte 2 À esquerda, a regra 1; ao centro, a regra 2; à direita, a regra 3 (N=200, q=5%). ...84
Lista de Tabelas
Sumário
1 Introdução ... 15
2 Conceitos básicos ... 18
2.1 Grafo ... 18
2.1.1 Distribuição de graus ... 18
2.1.2 Caminho mínimo médio... 19
2.1.3 Coeficiente de agregação ... 20
2.2 Autômatos celulares ... 21
2.3 Redes neurais e sincronismo ... 23
3 O modelo proposto ... 28
3.1 Topologia e vizinhança ... 28
3.2 O método de criação da rede aleatória ... 28
3.3 Estados possíveis ... 29
3.4 As regras de transição ... 29
3.5 Condições iniciais ... 32
3.6 Condições de contorno ... 32
3.7 Tipo de autômato ... 32
3.8 Definições ... 32
3.8.1 Sincronismo ... 32
3.8.2 Tipos de oscilação... 33
3.8.3 Tipos de sincronismo ... 33
3.9 Simulações e Medidas ... 34
4 Resultados ... 36
4.1 Caracterização dos reticulados ... 36
4.1.1 Caminho mínimo médio... 36
4.1.2 Comparação do caminho mínimo médio em redes de diferentes tamanhos ... 38
4.1.3 Coeficiente de agregação ... 39
4.2 O comportamento dinâmico das redes ... 40
4.2.1 Oscilações ... 42
4.2.2 Sincronizações ... 48
4.2.3 Caminho mínimo médio... 51
4.2.4 Razão entre o período da parte 1 e o período da entrada ... 53
4.2.6 Tempo para atingir a atividade máxima ... 57
4.3 Diferentes tamanhos de rede ... 59
4.3.1 Oscilações ... 59
4.3.2 Sincronizações ... 62
4.3.3 Caminho mínimo médio... 62
4.3.4 Razão entre o período da parte 1 e o período da entrada ... 64
4.3.5 Atividade máxima ... 64
4.3.6 Tempo para atingir a atividade máxima ... 66
4.4 Diferentes percentuais de ligações adicionadas ... 68
4.4.1 Oscilações ... 68
4.4.2 Sincronizações ... 71
4.4.3 Caminho mínimo médio... 72
4.4.4 Razão entre o período da parte 1 e o período da entrada ... 73
4.4.5 Atividade máxima ... 73
4.4.6 Tempo para atingir a atividade máxima ... 75
4.5 Diferentes regras de transição ... 77
4.5.1 Oscilações ... 77
4.5.2 Sincronizações ... 80
4.5.3 Razão entre o período da parte 1 e o período da entrada ... 81
4.5.4 Atividade máxima ... 81
4.5.5 Tempo para atingir a atividade máxima ... 83
5 Análise dos resultados ... 85
5.1 Oscilações ... 85
5.2 Sincronizações ... 86
5.3 Atividade máxima ... 87
5.4 Tempo para atingir a atividade máxima ... 87
6 Conclusões ... 89
1 Introdução
O cérebro humano pode ser considerado uma rede complexa, pois é composto por uma infinidade de células que se ligam através de inúmeras conexões realizadas tanto a curtas e quanto a longas distâncias. Nessa rede, a atividade numa área influencia a atividade em outras áreas; e as ligações estruturais são também ligações dinâmicas. Sincronismo é uma manifestação dessa simultaneidade de atividades, dessas influências mútuas.
Investigar a ocorrência de sincronismo é relevante, pois ele está intrinsecamente relacionado a diversos processos fundamentais para a sobrevivência de seres vivos, como cognição, percepção, locomoção e processos que envolvem integração dessas funções [1-3]. Por exemplo, trabalhos experimentais associam a sincronização de disparos neuronais no córtex visual com a segmentação e a compreensão de imagens por mamíferos [4]; e há evidência de atividades sincronizadas, com codificação em populações neuronais e correlação temporal, no reconhecimento de imagens [5]. Níveis anormais de sincronismo, entretanto, estão relacionados a desordens neurológicas, como autismo, epilepsia e esquizofrenia [6,7]. Portanto, estudos sobre sincronismo em redes neurais podem ajudar a compreender tanto o funcionamento normal quanto o patológico do cérebro.
O funcionamento de cada neurônio é repleto de detalhes e particularidades. Também existe uma complexidade associada às sinapses, governadas por fenômenos elétricos e bioquímicos. Ainda, há diversas substâncias envolvidas na transmissão da atividade entre os neurônios. Composto por aproximadamente 1011 neurônios e 1014 sinapses [2], o cérebro humano é, de fato, um sistema de difícil modelagem.
Portanto, um modelo dessa rede baseia-se, inevitavelmente, em muitas abstrações e simplificações. Em geral, os estudos teóricos lidam com uma única rede e usam autômatos celulares [7], equações diferenciais [8] ou equações de diferenças [9] para representar a atividade dos neurônios.
Investiga-se, via simulação numérica, a ocorrência de sincronismo entre duas redes neurais, sendo que uma delas recebe um estímulo periódico. Com isso, busca-se entender as relações observadas entre as frequências de oscilação de diferentes estruturas neurais acopladas, como, por exemplo, entre a retina, o núcleo geniculado lateral e o córtex visual de mamíferos [10,11]. Essas duas redes são conectadas por ligações aleatórias. A dinâmica exibida por esse sistema, composto por duas partes (duas redes), é estudado em função de variações no número de ligações adicionadas (tanto em cada parte como entre as partes), nas regras de transição dos estados, no tamanho dos reticulados. A primeira parte recebe um estímulo periódico que gera comportamento oscilatório que se propaga para a segunda parte. No modelo, as ligações entre as partes são direcionadas; isto é, somente a primeira parte influencia a segunda parte, não ocorrendo influência no sentido contrário.
As simulações numéricas executadas, considerando estímulos de diferentes períodos e diferentes parâmetros para as redes neurais, revelam que sincronismo ocorre com alta probabilidade e que outros tipos de comportamento oscilatório também aparecem. O comportamento dinâmico das redes, em reação à variação dos valores dos parâmetros, não é linear, em geral.
2 Conceitos básicos
Usam-se grafos para representar as características topológicas de um sistema composto de várias unidades. Com grafos, pode-se expressar como suas unidades (nós) se conectam e se influenciam. A dinâmica desses nós pode ser descrita por autômatos celulares, que são definidos em termos de seus estados permitidos, de suas regras de transição, da topologia da vizinhança e da condição de contorno. Esses assuntos, detalhados a seguir, têm muitos pontos em comum e são fundamentais para a construção do este modelo.
2.1 Grafo
Relações entre entidades do mundo real podem ser representadas graficamente por meio de grafos. Por exemplo, os nós de uma rede podem representar pessoas e as arestas que conectam esses nós podem indicar relações de amizade entre essas pessoas. Ou nós podem representar neurônios e, as arestas que os unem, as ligações sinápticas entre tais neurônios. Quando há um conjunto de elementos que se relacionam, é possível representar esses elementos e as relações entre eles através de um grafo, de uma rede [1,12,13].
Grafo, nome que deriva dessa possibilidade de representação gráfica, é formalmente definido como uma tripla ordenada G= (V(G), A(G), ), sendo G o grafo em questão, V(G) o conjunto de vértices ou nós, A(G) o conjunto de arestas (conexões) e uma função de ponderação das relações entre os nós. As conexões entre os nós podem ser direcionadas ou não [1,12,13]. Se não há ponderação, de modo que todas as arestas têm o mesmo peso (a mesma intensidade), o grafo é considerado não ponderado. Grafos com autoconexões (ou seja, com conexões que ligam um nó a ele mesmo) e com conexões múltiplas (ou seja, pode haver mais de uma conexão entre um mesmo par de nós) são chamados de grafos degenerados.
O modelo de rede neural utilizado neste trabalho é baseado em grafos não direcionados, não ponderados e não degenerados.
2.1.1 Distribuição de graus
graus de um grafo aleatório, que tem forma de sino, e a de uma rede livre-de-escala, que é o do tipo P(k) ∼ k−γ (com γ > 0). Essas distribuições são mostradas no Gráfico 2.1.
Gráfico 2.1– Quantidade nós com grau k em função de k para uma rede aleatória (à esquerda) e para uma rede livre-de-escala (à direita). Para se obter P(k), devem-se dividir os números do eixo vertical por N=10000, que é o número total de nós (figura retirada e adaptada de [13]).
Seja P(k) a fração de nós com k ligações. O grau médio ̅ é definido por:
̅ ∑
Observe-se que na rede aleatória do Gráfico 2.1, o grau médio está próximo do grau mais frequente; enquanto que na rede livre-de-escala, não. Nessa última, nós com grau pequeno existem em muito maior número do que nós com grau grande.
2.1.2 Caminho mínimo médio
O caminho mínimo médio l é a menor distância média entre os nós; ou seja, é, em média, o menor número de arestas que conecta qualquer par de nós da rede. É uma medida de integração da rede: quanto menor l, maior a integração.
O caminho mínimo médio é matematicamente definido por [1,13]:
̅ ∑
2.1.3 Coeficiente de agregação
Watts e Strogatz [14] propuseram uma medida para avaliar a “clusterização” em grafos. Basicamente, ela indica a proporção de vizinhos de um nó que estão conectados entre si. Assim, o coeficiente de agregação Ci do nó i é dado por [1,2]:
sendo é o número de conexões entre vizinhos do nó i e ki(ki-1)/2 o número máximo de conexões que poderia existir entre esses vizinhos. Quanto maior o número de conexões locais, maior o valor dessa medida, que indica a capacidade de processamento local.
O grafo aleatório proposto por Erdös e Rényi (ER) [1,5] é construído a partir de N nós desconectados, realizando-se conexões entre eles com uma determinada probabilidade p. A relação entre o grau médio ̅ e p é dada por [1]:
̅
Esse grafo aleatório costuma ser tomado em comparações para questões de conectividade de grafos complexos [1,13].
A topologia conhecida como small-world é um grafo aleatório, não direcionado, que se situa entre o reticulado regular e o grafo ER. Isso está ilustrado na Gráfico 2.2. De fato, Watts e Strogatz [14] construíram uma rede com algumas características do reticulado regular e com algumas do grafo aleatório ER, pois [1,14]:
como no grafo ER, um grafo small-world tem caminho médio pequeno (da ordem do logaritmo de N) e distribuição de graus com formato de sino;
como nos reticulados regulares, um grafo small-world tem coeficiente de agregação alto (da ordem de 1).
Essas características dão à rede duas propriedades:
um caminho mínimo médio “pequeno” (mais curto) facilita a integração entre diferentes partes da rede;
A coexistência dessas duas propriedades é encontrada em várias redes do mundo real, como relacionamentos de sites na internet, redes neurais naturais, redes de amigos [1,14], etc.
Gráfico 2.2 - Small-world como grafo intermediário entre o totalmente aleatório e o regular (retirado de [14]).
2.2 Autômatos celulares
A ideia de autômato celular está ligada à própria concepção dos computadores [16]. Sua capacidade de realizar processamento local e de representar processos complexos remonta aos modelos de rede de neurônios de McCulloch e Pitts e à teoria de autômato autorreprodutível de von Neumann. Investigações sobre similaridades com o funcionamento do sistema nervoso também são recorrentes [17].
Estudos de von Neumann sobre sistemas capazes de se autorreproduzirem, com motivação biológica, realizados na década de 50 do século passado, utilizaram, pela primeira vez, autômatos celulares [18]. Autômatos são particularmente adequados para representar sistemas complexos que possuem diversas partes que interagem entre si, que possuem um número de estados possíveis finitos, e que evoluem em tempo discreto.
Conway [19], analisando as possibilidades de um determinado autômato bidimensional, criou o famoso Jogo da Vida. A partir de diferentes configurações iniciais, aplicando as mesmas regras de transição, obtêm-se evoluções espaços-temporais diferentes. Essa sensibilidade a condições iniciais é comum a vários sistemas complexos.
Há dois tipos de estudos normalmente feitos com autômatos celulares. Usualmente parte-se de um determinado autômato e verificam-se suas propriedades, ou tenta-se criar um autômato para resolver um determinado problema, uma determinada tarefa computacional.
Sistemas compostos de muitos elementos com forte interação local são apropriados à modelagem por autômatos celulares. Sistemas biológicos complexos, como ecossistemas, tecidos cardíacos e redes neurais, se enquadram nessa categoria [20].
Além de sistemas biológicos, há muitos outros exemplos de estudos empregando autômatos celulares, como: crescimento de cristais, tráfego de automóveis, processamento de imagens, evolução de galáxias espirais [18, 20]. Ainda, autômatos celulares possibilitam soluções discretas aproximadas de sistemas de equações diferenciais, como o modelo de dinâmica populacional de Lotka-Volterra, o modelo epidemiológico SIR e o de difusão de Navier-Stokes [16, 21].
A construção de modelos bioinspirados com autômatos celulares é mais intuitiva do que com equações diferenciais, pois autômatos permitem a representação de diferentes topologias de acoplamento de modo natural e a elaboração de regras de evolução temporal simples com motivação biológica [22]. Experimentos numéricos realizados com autômatos celulares mostram que comportamentos complexos podem emergir de um conjunto composto por muitas interações locais simples. Esses estudos podem ajudar a entender, por exemplo, como a consciência pode emergir das ligações sinápticas entre os neurônios ou como cultura pode emergir da interação das crenças e dos costumes dos indivíduos [20].
Por isso, autômatos celulares têm sido usados para modelar fenômenos biológicos e sociais, já que é possível propor regras baseadas em situações reais que sejam facilmente programáveis num computador. Por exemplo, simulações de propagação de doenças contagiosas com autômatos celulares evidenciam a influência do tamanho das cidades na manutenção ou erradicação de tais doenças. Esses resultados são confirmados por observações experimentais [21].
reticulado bidimensional regular, com vizinhança regular de raio unitário acrescida de ligações aleatórias, com três estados possíveis para cada célula, com regras de transição deterministas, com condição de contorno aberta (ou seja, as células da borda têm menos vizinhos do que as centrais).
Antes de se especificar mais claramente o modelo usado neste trabalho, apresentam-se definições e estudos relacionados com sincronismo em redes neurais.
2.3 Redes neurais e sincronismo
Neurônios são as células do sistema nervoso responsáveis pelo processamento da informação cognitiva e sensorial. Eles produzem pulsos elétricos devido à variação da permeabilidade seletiva da membrana a íons presentes no seu interior e no meio extracelular. Esses pulsos codificam a informação recebida por outros sistemas, como músculos e órgãos sensoriais.
O cérebro humano tem aproximadamente 1011 neurônios e 1014 sinapses [2]; ou seja, é uma rede gigantesca e intensamente conectada. Essa complexidade exige uma grande simplificação dos modelos usados para estudá-lo.
Podem-se identificar, simplificadamente, três estados para o potencial de membrana de um neurônio: repouso, disparo e refratário. Esses estados são usados no este modelo. A seguir, descrevem-se alguns modelos e estudos referentes a sincronismo de redes neurais.
Sistemas complexos são normalmente formados por um grande número de componentes conectados. Esses sistemas surgem em diversos contextos, como o biológico, o social e o tecnológico. Vários estudos, com vários modelos, em várias áreas do conhecimento, procuram descobrir as condições para que os componentes de um sistema funcionem sincronamente [1]. O conhecimento atual sobre o funcionamento do cérebro mostra que a coordenação de informações processadas em diferentes áreas é necessária para a realização de diversas atividades cognitivas, sensoriais e motoras [24-26]. Assim, existe um alto paralelismo das atividades distribuídas pelas diversas áreas do cérebro, envolvendo grandes populações de neurônios. Evidências neurofisiológicas mostram que redes corticais possuem as estruturas e os mecanismos necessários para gerarem e manterem padrões neurais de sincronização seletiva [27].
Gray [5] revisou vários estudos sobre fisiologia e anatomia do sistema visual que oferecem pistas sobre os processos neuronais que suportam seu funcionamento complexo, como codificação da atividade em populações e correlação temporal. Por exemplo, a hipótese da agregação e correlação temporal afirma que as diferentes áreas envolvidas no processamento de uma imagem reforçam sua ligação pela sincronização dos disparos neuronais. A intensidade das ligações sinápticas é aumentada transientemente quando estímulos aferentes são síncronos com estímulos eferentes dos neurônios. Isto provê um mecanismo de retroalimentação positiva que amplificaria fracas correlações de atividades entre áreas distintas. Ainda, o padrão dos disparos seria outro modo de diferenciar as diferentes atividades presentes simultaneamente na rede neural [5]. Gray apontou resultados experimentais que mostram a importância da sincronização de disparos numa escala temporal de milissegundos e a dependência dessa sincronização com os estímulos.
imagem [5]. De fato, existem muitas evidências da participação de diferentes áreas corticais na identificação e na localização espacial de objetos. Para isso, é necessário ocorrer a integração de informações distribuídas [5].
Ainda sobre o sistema visual, diferentes propriedades percebidas de um mesmo objeto, como sua cor, sua velocidade e sua forma, são processadas em áreas diferentes do córtex visual. Não está ainda completamente esclarecido como essas propriedades são unificadas para a percepção de um único objeto. Entretanto, a sincronização de grupos de neurônios, em várias camadas do córtex visual, é o mecanismo proposto atualmente mais aceito [5, 28].
Em alguns modelos matemáticos sobre esse assunto, toma-se uma coluna de neurônios como a unidade básica e a existência de oscilação como condição necessária para a sincronização. Num desses modelos, usam-se as equações de Wilson-Cowan, que considera populações de neurônios excitatórios e inibitórios [28]. Esse modelo permite, inclusive, haver oscilação sem estímulos externos, indicando ser a oscilação uma propriedade intrínseca do sistema, resultado do equilíbrio entre forças excitatórias e inibitórias.
Kopell [29] estudou a ocorrência de sincronismo em diferentes frequências entre grupos distantes de neurônios. Na comunicação entre esses grupos está implícito um atraso na condução da informação, já que está se dá com velocidade finita. No estudo realizado, constatou-se que a frequência das ondas beta (da ordem de 15-30 Hz) permite sincronização para atrasos maiores, associados a maiores distâncias entre os grupos; enquanto que a frequência das ondas gama (maiores que 40 Hz) permite sincronização para atrasos menores. Para estudar detalhadamente os mecanismos de sincronismo, Kopell concentrou sua atenção em um pequeno grupo de neurônios. Seu modelo minimalista é formado por apenas quatro neurônios, dois excitatórios e dois inibitórios, conforme ilustra a Figura 3.2. As relações pontilhadas representam ligações ora presentes, ora ausentes. As unidades excitatórias (E) representam células piramidais e as unidades inibitórias (I) representam interneurônios. As linhas representam as conexões entre as células. Resultados das simulações mostram que o ritmo das oscilações síncronas é determinado principalmente pelas inibições.
Figura 2.2 – Modelo de um grupo de neurônios locais (retirado de [29]).
Um processo cognitivo que envolve várias áreas cerebrais é a percepção. A percepção envolve múltiplas etapas; etapas simples como a identificação da cor, e etapas complexas, como a consolidação de várias propriedades em objetos (como suas cores, formas, sensações de movimento). A teoria da Gestalt propõe que o sistema cognitivo cria totalidades em função de proximidade, similaridade, continuidade, fechamento e destino comum. E que a agregação sensorial é um processo dinâmico e automático, que une características distintas em objetos que são criados pela percepção. Outros estudos mostram que tamanho, textura, disparidade binocular e coincidência no tempo também geram efeitos semelhantes [5].
Essa representação e unificação de características diversas em um único objeto devem repousar sobre um substrato neural de acoplamento e de sincronização de neurônios distribuídos pelo cérebro. Esse mecanismo explicaria a rapidez, o paralelismo e a natureza combinatória da atividade de percepção.
Outro fenômeno cerebral no qual o sincronismo desempenha papel importante, mas patológico, é a epilepsia. Volk [7] usou modelos discretos para estudar a atividade epilética. Segundo ele, seus modelos podem reproduzir o padrão encontrado em eletroencefalogramas de ratos sofrendo ataques epilépticos, que se caracterizam por um aumento de amplitude de oscilação e com atividade oscilatória regular destacada. Em regime normal, observam-se oscilações aparentemente aleatórias e de menor amplitude. Esse fenômeno de maior amplitude e maior regularidade das oscilações é referido como hipersincronização.
3 O modelo proposto
“Se em tudo o mais forem idênticas, as várias explicações de um fenômeno, a mais simples é a melhor” (William de Ockham, filósofo, 1288–1348).
O modelo proposto pretende, de forma simples, reproduzir o comportamento oscilatório e sincronizado que ocorre em sistemas neurais. Seguem-se as especificações do modelo utilizado.
3.1 Topologia e vizinhança
Usa-se um reticulado bidimensional, retangular, com vizinhança de von Neumann de raio unitário; ou seja, cada célula está ligada a outras quatro, que são aquelas a norte, sul, leste e oeste. A esse reticulado regular são acrescentadas aleatoriamente ligações. A rede resultante lembra a que foi proposta por Newman e Watts [30-32] e que tem características de topologia small-world. Esse reticulado é dividido em duas partes iguais.
Diferentemente da rede small-world proposta por Watts e Strogatz [14] que elimina, aleatoriamente, ligações da rede regular, o modelo de Newman-Watts as acrescenta, também de modo aleatório. Este procedimento evita a existência de partes isoladas na rede, o que diminuiria sua capacidade de integração, como comentado na seção 4.1.
3.2 O método de criação da rede aleatória
Parte-se de um grafo regular, não direcionado, não ponderado, de tamanho m×n (m linhas e n colunas), sendo n par. Em tal grafo, as células são posicionadas e conectadas por ligações de mesmo tamanho e em ângulo reto, conforme mostra a Figura 3.1. São retiradas as ligações entre as células das colunas n/2 e (n+2)/2, de forma a se ter duas partes iguais com m linhas e n/2 colunas. Em cada parte, inclui-se aleatoriamente um determinado percentual de ligações entre células da mesma parte. São assim construídas duas partes com ligações predominantemente locais, entre células vizinhas. A essas partes são também inseridas ligações entre elas, em um determinado percentual.
acrescentado ligando as duas partes, ou seja, há 9 ligações entre as duas partes. As ligações são todas excitatórias e têm peso unitário.
Figura 3.1 - Autômato de duas partes, baseado em grafo regular (ligações em preto) com m=10 e n=20, com ligações aleatórias intrapartes (ligações em azul) e entrepartes (ligações em verde).
3.3 Estados possíveis
A cada instante, cada célula está em um de três estados: estado excitado, estado de repouso, estado refratário. Assim, os estados possíveis do neurônio são três, que são os estados básicos de um neurônio natural. E é o mesmo número de estados de um modelo SIR usado em estudos sobre propagação de doenças contagiosas [21].
3.4 As regras de transição
Procuram-se usar regras de transição que permitam gerar oscilações. Não são adequadas regras que não estimulem suficientemente a rede para que oscilações se mantenham. O Gráfico 3.1 é um exemplo de resposta da rede a uma regra hipoexcitante.
Gráfico 3.1 – Evolução do número de células excitadas em uma rede, que evolui monotonamente para repouso total.
Como se percebe, a atividade no estado inicial (10 células excitadas) vai monotonamente diminuindo, e a rede vai para estado de completo repouso após 5 passos de tempo (a regra 1, descrita a seguir, foi utilizada para gerar esse gráfico, com limiar igual a 2).
2 4 6 8 10 12 14 t
2 4 6 8 10 x
Outro conjunto de regras que não são adequadas para este estudo são as que levam a rede ao estado de completa atividade, chamadas aqui de regras hiperexcitantes. Considere-se que uma rede de tamanho 20×10 evolua segundo a seguinte regra:
se uma célula possui vizinho ativo no passo de tempo t, ela vai para o estado ativo em t+1;
se uma célula não possui vizinho ativo em t, ela vai para o estado de repouso em t+1.
Gráfico 3.2- Evolução do número de células excitadas em uma rede, que evolui para atividade total.
Nesse caso, como mostra o Gráfico 3.2, todas as células ficam constantemente estimuladas, conforme o tempo passa.
As regras utilizadas neste estudo devem ser capazes de produzir uma resposta oscilatória a um estímulo conforme mostra o Gráfico 3.3.
Gráfico 3.3- Evolução do número de células excitadas em uma rede, cuja atividade total oscila.
A atividade total oscila: ela não converge monotonamente para zero, como no caso do Gráfico 3.1 e nem converge monotonamente para atividade máxima, como no caso do Gráfico 3.2.
2 4 6 8 10 12 14 t
50 100 150 200 x
2 4 6 8 10 12 14 t
Aqui, as regras são aplicadas de modo síncrono; isto é, são aplicadas simultaneamente a todas as células, em cada passo de tempo.
Durante as simulações foram usadas 3 regras de transição de estado. A regra 1, que permite um “disparo estendido”, é assim:
uma célula em repouso no instante t que recebe um total de impulsos, vindos das células ligadas a ela, que é maior ou igual ao limiar (de valor unitário para todas as células), dispara em t+1;
uma célula que disparou em t, e que recebe um total de impulsos maior ou igual ao limiar, continua disparando em t+1 (o que é aqui chamado de “disparo estendido”); caso contrário, vai para o estado refratário;
uma célula que disparou em t e t+1 vai para o estado refratário em t+2, independentemente do estímulo recebido;
uma célula em estado refratário em t vai para o repouso em t+1. A Figura 3.1 foi gerada usando essa regra, com limiar igual a 2.
A regra 2, que permite dois disparos estendidos, é dada por:
uma célula em repouso em t que recebe um total de impulsos maior ou igual ao limiar, dispara em t+1;
uma célula que disparou em t, e que recebe um total de impulsos maior ou igual ao limiar, continua disparando em t+1; caso contrário, vai para o repouso;
uma célula em disparou em t e t+1, e que recebe um total de impulsos maior ou igual ao limiar, continua disparando em t+2; caso contrário, vai para o repouso;
uma célula que disparou em t, t+1 e t+2 vai para o estado refratário em t+3;
uma célula em estado refratário em t vai para o repouso em t+1. A regra 3, que permite três disparos estendidos, é assim:
uma célula em repouso em t que recebe um total de impulsos maior ou igual ao limiar, dispara em t+1;
uma célula que disparou em t, e que recebe um total de impulsos maior ou igual ao limiar, continua disparando em t+1; caso contrário, vai para o repouso;
uma célula em disparou em t, t+1 e t+2, e que recebe um total de impulsos maior ou igual ao limiar, continua disparando em t+3; caso contrário, vai para o repouso;
uma célula que disparou em t, t+1, t+2 e t+3, vai para o estado refratário em t+4;
uma célula em estado refratário em t vai para o repouso em t+1.
Essas regras pressupõem a existência de uma rede oculta de sinapses inibitórias que forçaria os neurônios que dispararam seguidas vezes a irem, obrigatoriamente, para uma situação de não disparo no instante seguinte. Assim, inibição, uma característica fundamental em redes neurais naturais, está embutida nas regras de transição de estados.
3.5 Condições iniciais
Nas simulações executadas, a rede está inicialmente em repouso, com apenas uma célula da primeira coluna da parte 1 ativa (essa célula a ser estimulada é uma das células da coluna mais à esquerda da parte 1, que é a coluna mais distante da parte 2). Cada célula da primeira coluna da parte 1 foi excitada, nas diferentes simulações realizadas.
3.6 Condições de contorno
A rede tem contorno aberto; isto é, as células que estão nas bordas só são influenciadas por aquelas que estão à sua volta (elas têm, de fato, menos vizinhos do que as células que não estão nas bordas).
3.7 Tipo de autômato
A rede é governada por regras deterministas. Assim, o autômato é determinista, possuindo ligações aleatoriamente acrescentadas em sua topologia.
3.8 Definições
3.8.1 Sincronismo
Com esse autômato celular com conexões aleatórias, mas preferencialmente locais, estuda-se a ocorrência de sincronismo, para diferentes estímulos e configurações da rede. Os resultados das simulações numéricas realizadas são mostrados no capítulo 4.
3.8.2 Tipos de oscilação
Os comportamentos das redes são classificados conforme a relação entre os períodos de oscilação das duas partes. Aqui, denomina-se:
igual: quando as partes oscilam com o mesmo período, o que equivale à definição de sincronismo;
múltiplo: quando a parte 2 oscila em um período que é múltiplo do período da parte 1. Por exemplo, a parte 1 oscila com período de 10 passos de tempo e a parte 2 oscila com período de 20 passos;
submúltiplo: quando a parte 2 oscila em um período que é submúltiplo do período da parte 1. Por exemplo, a parte 1 oscila com período de 20 passos e a parte 2 oscila com período de 10 passos;
menor: quando a parte 2 oscila em um período que não é nem múltiplo nem submúltiplo e é menor que o período da parte 1. Por exemplo, a parte 1 oscila com período de 9 passos e a parte 2 oscila com período de 6 passos;
maior: quando a parte 2 oscila em um período que não é nem múltiplo nem submúltiplo e é maior que o período da parte 1. Por exemplo, a parte 1 oscila com período de 6 passos e a parte 2 oscila com período de 9 passos.
3.8.3 Tipos de sincronismo
Sincronismo, como definido, significa oscilação das partes no mesmo período. Essa oscilação pode ocorrer no mesmo período da entrada ou em outro período. Os tipos de sincronismo assinalam a relação entre o período de oscilação das partes e o período de entrada. Chama-se:
igual: quando as partes oscilam com o mesmo período da entrada;
múltiplo: quando as partes oscilam em um período que é múltiplo do período da entrada. Por exemplo, as partes oscilam com período de 20 passos e o período de entrada é de 10 passos.
menor: quando as partes oscilam em um período que não é nem múltiplo nem submúltiplo do período de entrada e é menor que este. Por exemplo, as partes oscilam com período de 6 passos e o período da entrada é de 9 passos.
maior: quando as partes oscilam em um período que não é nem múltiplo nem submúltiplo do período de entrada e é maior que este. Por exemplo, as partes oscilam com período de 9 passos e o período da entrada é de 6 passos.
3.9 Simulações: Estímulos Externos, Parâmetros e Medições
Inicialmente, foram geradas 20 redes aleatórias. Uma simulação consiste de 100 passos de tempo, de modo que a cada passo a regra de transição de estados é aplicada. A rede, totalmente em repouso, tem uma célula da primeira coluna da parte 1 levada ao estado ativo no passo 1. Esse estímulo externo, forçado, gera atividade nos vizinhos, e essa atividade se propaga para parte 2. Após um determinado período de tempo (o período do estímulo), essa mesma célula é novamente levada ao estado ativo. Cada rede tem 10 células na primeira coluna (aquela mais a esquerda). Simulações são realizadas considerando cada uma das 10 células da primeira como a célula a ser estimulada. Para cada configuração utilizada, são feitas simulações com 20 períodos, sendo que os períodos variam de 1 passo a 20 passos de tempo. Ou seja, para cada rede (construída com ligações locais regulares e ligações adicionais aleatórias), são feitas 200 simulações: 10 diferentes células estimuladas vezes 20 diferentes períodos de estímulo.
Partiu-se da seguinte configuração:
Tamanho da rede (N): 20×10, o que totaliza 200 células.
Quantidade de ligações adicionais (q): 5% de ligações aleatórias adicionadas em cada parte, e esse mesmo percentual entre as partes.
Regra de transição de estados: regra 1 descrita na seção 3.4.
Foram feitas 4.000 simulações (20 redes, com 200 simulações para cada rede).
A seguir, foram geradas redes de diferentes tamanhos, para observar a influência no comportamento quando o tamanho varia. Para isso, outras duas configurações foram geradas:
redes de 60×10 (600 células);
redes de 100×10 (1000 células).
Para se analisar a influência dos percentuais de ligações aleatórias adicionadas, o mesmo procedimento foi usado: mais 20 novas redes geradas para cada configuração:
redes com 10% de ligações adicionais;
redes com 15% de ligações adicionais.
Por fim, modificou-se a regra de transição, para verificar sua influência no comportamento oscilatório. Baseadas nas redes inicialmente usadas (tamanho 20×10, 5% de ligações adicionais), foram feitas simulações com a regra 2 de transição de estado e com a regra 3 de transição de estado, descritas na seção 3.4.
Assim, foram feitas simulações com três tamanhos de rede e três percentuais de ligações adicionadas para a regra 1, e simulações com um único tamanho de rede e com único porcentual de ligação para as três regras. No total, são sete diferentes configurações testadas, totalizando 28.000 simulações executadas (4.000 simulações para cada configuração).
Em cada simulação, em cada passo, foi calculado o número de células ativas de cada uma das partes. Esse valor é a base das medidas efetuadas. Esse número é chamado atividade da rede.
O período de oscilação é calculado como o número de passos em que atividade da rede se repete. As considerações sobre oscilação, no próximo capítulo, baseiam-se no número de casos em que as partes das redes oscilam no mesmo período, em período múltiplo ou submúltiplo; ou não oscilam periodicamente, isto é, nenhuma sequência de atividade se repete no intervalo de passos simulado.
Os valores encontrados foram, em algumas análises, separados em tercis. Foram usados tercis para diferenciar três situações: os casos próximos à média, os casos menores que a média e os casos maiores que a média.
Sincronismo significa oscilação no mesmo período. Nas análises de sincronismo, verifica-se a relação entre o período de oscilação das partes e com o período do estímulo de entrada. Conta-se o número de simulações em que a rede sincronizou no mesmo período, em período múltiplo ou submúltiplo do período de entrada.
4 Resultados
Para cada tamanho (N) e para cada diferente percentual de ligações adicionais simulados (q), foram geradas 20 redes, conforme descrito no capítulo 3. A atividade de cada parte foi medida contando-se o número de células ativas em cada passo. Para cada rede e para cada período de entrada, foram feitas simulações considerando cada uma das células da coluna mais à esquerda da parte 1 (a coluna mais distante da parte 2) como aquela que recebe o estímulo.
4.1 Caracterização dos reticulados
As simulações numéricas foram realizadas com o auxílio do aplicativo Mathematica 6. A construção das redes, em virtude das ligações extras aleatórias, apresenta variabilidade intrínseca. A estrutura topológica da rede é caracterizada em termos do caminho mínimo médio e do coeficiente de agregação.
Nos resultados apresentados nesta seção, a regra de transição utilizada é a regra 1 (um disparo estendido) e as redes possuem 5% de ligações aleatórias adicionais.
4.1.1 Caminho mínimo médio
O caminho mínimo médio é uma medida da integração de um grafo, indicando quantos passos são necessários para, em média, um nó influenciar outro. Quanto menor esse caminho, mais integrado está o grafo. Para possibilitar comparações com outros tamanhos de redes, calculam-se informações percentuais, em relação ao grafo regular correspondente. Redes de 20 × 10
Tabela 4.1 – Medidas dos caminhos mínimos médios de redes de 20×10, e suas respectivas partes (10×10), em relação à rede regular correspondente (q=5%)..
caminho min médio parte1 parte2 conjunto
mínimo 69,7% 70,4% 55,4%
máximo 84,1% 76,9% 60,0%
média2 73,3% 73,5% 57,9%
desvio padrão 3,3 % 1,8 % 1,3 %
As médias das partes são muito parecidas e estão em torno de 73% do caminho mínimo médio da rede regular de 10×10. O conjunto é a união das partes 1 e 2 pelas ligações aleatórias entre as partes. Essas ligações fazem o caminho mínimo médio do conjunto diminuir para 58%, em relação ao grafo regular de 20×10.
Redes de 60 × 10
Os mesmos procedimentos foram executados para redes de 60 × 10. O caminho mínimo médio em um grafo retangular regular de 30 × 10 é 13,3; e o de uma rede de 60 × 10 é 23,3. Os resultados de 20 simulações são mostrados na Tabela 4.2.
Tabela 4.2 – Medidas dos caminhos mínimos médios de redes de 60×10, e suas respectivas partes (30×10), em relação ao grafo regular correspondente. (q=5%).
caminho mínimo médio parte1 parte2 conjunto
mínimo 47,1% 48,2% 29,9%
máximo 50,4% 54,7% 31,1%
média 49,2% 49,8% 30,3%
desvio padrão 0,73 % 1,53 % 0,32 %
As partes 1 e 2 apresentam um caminho mínimo médio de 8,5, que representa 43% do valor encontrado no grafo regular correspondente. No conjunto, essa medida vale 22% do valor do caminho mínimo médio de um reticulado regular de 20×100, que é de 40.
Redes de 100 × 10
Os mesmos procedimentos foram executados para redes de 100 × 10. O caminho mínimo médio em um grafo retangular regular de 50 × 10 é 20; e o de uma rede de 100 × 10 é 36,7. Os resultados de 20 simulações são mostrados na Tabela 4.3.
Tabela 4.3 – Medidas dos menores caminhos médios de redes de 20×100, e suas respectivas partes (10×100), em relação ao grafo regular correspondente (q=5%).
caminho mínimo médio parte1 parte2 conjunto
mínimo 36,0% 35,9% 20,8%
máximo 37,7% 37,9% 21,3%
média 36,9% 36,8% 21,0%
desvio padrão 0,50 % 0,61 % 0,16 %
As partes 1 e 2 apresentam um caminho mínimo médio de 8,5, que representa 43% do valor encontrado no grafo regular correspondente. No conjunto, essa medida vale 22% do valor do caminho mínimo médio de um reticulado regular de 20×100, que é de 40.
4.1.2 Comparação do caminho mínimo médio em redes de diferentes
tamanhos
Comparando os resultados, verifica-se o desvio padrão diminui com o aumento do tamanho do grafo, revelando que um maior número absoluto de ligações compensa eventuais peculiaridades advindas da aleatoriedade das ligações adicionadas. Também, o percentual do caminho mínimo médio, relativo à medida no grafo regular correspondente, mostra-se influenciado pelo tamanho do grafo.
Gráfico 4.1 – Caminho mínimo médio como percentual em relação ao grafo regular, em função do tamanho da rede. Escala horizontal logarítmica (q=5%)..
20 30 40 50 60 70 80
100 1000
%
gr
a
fo
regu
a
lr
tamanho da rede (Log)
O Gráfico 4.1 mostra a relação entre o caminho mínimo médio das redes geradas e o tamanho das redes. Nesse gráfico, o caminho mínimo médio das redes está representado como o percentual em relação a um grafo regular de mesmo tamanho, mesmo número de nós. Nota-se uma relação logarítmica entre o caminho mínimo médio e o tamanho da rede, o que está de acordo com outros estudos [30]. Ou seja, quanto maior a rede, a inserção de um percentual fixo de ligações aleatórias (5%, como já explicado) leva a uma maior redução no caminho mínimo médio, comparando-se com o grafo regular correspondente.
4.1.3 Coeficiente de agregação
Conforme definido anteriormente, o coeficiente de agregação avalia a porcentagem de vizinhos de um mesmo nó que estão conectados entre si. Numa rede social, isso indicaria quantos amigos de uma pessoa são amigos entre si. Como esta rede é construída a partir de retângulos, não há triângulos nelas. Assim, esse coeficiente é zero ou ligeiramente maior do que zero, dependendo das ligações aleatórias acrescentadas formarem triângulos ou não.
No entanto, se esse coeficiente fosse definido a partir da formação de retângulos, ele seria muito alto. Nessa definição, contam-se quantos vizinhos de um nó têm um vizinho em comum (na definição original, contam-se quantos vizinhos de um nó têm uma aresta em comum). O Gráfico 4.2 ilustra essa nova definição. O nó considerado, em vermelho, tem os nós amarelos como vizinhos. Os nós verdes conectam um vizinho a outros dois. Assim, esse coeficiente valeria aproximadamente 2/3.
Gráfico 4.2 - Vizinhança do nó 13, em vermelho; os nós amarelos são os vizinhos diretos; os nós verdes ligam um vizinho (amarelo) a outros 2 dos 3 vizinhos.
1 2 3 4 5
6 7 8 9 10
11 12 13 14 15
16 17 18 19 20
4.2 O comportamento dinâmico das redes
A configuração que serviu de base para as simulações, aquela a partir da qual foram geradas modificações na configuração dos parâmetros, foi uma rede de 20×10, com 5% de ligações aleatórias adicionadas em cada parte, e esse mesmo percentual entre as partes, e usando a regra 1 de transição de estados.
Em cada simulação, evolui-se a rede durante 100 passos de tempo. O período de oscilação de cada parte é calculado como o número de passos em que um padrão se repete. Foram calculados os números de passos de tempo em que as partes das redes atingem o máximo de células estimuladas, em cada uma das partes, assim como o percentual de células ativas nesse máximo.
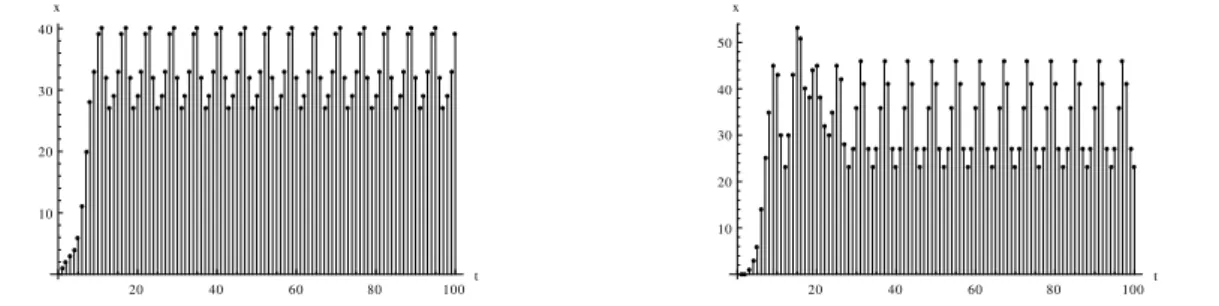
O Gráfico 4.3 mostra um exemplo dessas oscilações: a entrada tem período igual a 3, a parte 1 e a parte 2 têm período igual a 6.
Gráfico 4.3 – Total de células ativas nas parte 1 (esquerda) e na parte 2 (direita) – as partes estão sincronizadas com período de 6 passos. N=200, q=5%.
Nas 4.000 simulações (20 redes × 20 períodos × 10 células de entrada) executadas com essa configuração, observou-se grande predominância de casos das partes oscilando no mesmo período. O Gráfico 4.4 ilustra os resultados obtidos. As redes foram geradas aleatoriamente e apresentaram, para todos os períodos, no mínimo 75% de casos “igual”
(partes oscilando no mesmo período; conforme a seção 3.8.2). 20 40 60 80 100t
10 20 30 40 x
20 40 60 80 100t 10
Gráfico 4.4 – Relação entre o período de oscilação da parte 2 e o período da parte 1 (veja seção 3.8.2). N=200, q=5%.
Pode-se perceber a semelhança de comportamento quando os dados são tabulados (Tabela 4.4). Os dados estão dentro do intervalo de dois desvios padrão das médias de cada categoria de oscilação (ou muito próximos). As redes 10 e 11 têm número de casos de
“múltiplo” que ultrapassa o limite superior desse intervalo, mas muito próximos a ele: 39
casos quando o limite da faixa é 38,9. A rede 11 tem casos de “igual” ligeiramente inferior ao
limite de dois desvios padrão: 150 contra 151,6; a rede 12 tem número de casos sem oscilação ligeiramente menor: 1 contra 1,7; e a rede 14 casos de submúltiplos ligeiramente superior: 7 contra 6,1. Ou seja, a variação encontrada entre as redes está dentro do esperado para uma distribuição normal e nenhuma rede apresentou comportamento que mereça destaque.
0 20 40 60 80 100 120 140 160 180 200
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
n
o
.
ca
so
s
rede
Oscilação x Rede
igual
múltiplo
submúltiplo
Tabela 4.4 – Relação entre o período de oscilação da parte 2 e o período da parte 1, para cada rede gerada. N=200, q=5%.
contagem oscilação
rede igual múltiplo submúltiplo não total
1 167 25 0 8 200
2 192 2 0 6 200
3 177 13 5 5 200
4 179 18 0 3 200
5 182 11 0 7 200
6 174 18 1 7 200
7 177 18 1 4 200
8 184 9 0 7 200
9 181 6 6 7 200
10 156 39 0 5 200
11 150 39 0 11 200
12 196 3 0 1 200
13 174 17 2 7 200
14 178 7 7 8 200
15 171 19 5 5 200
16 184 10 0 6 200
17 170 23 0 7 200
18 179 15 1 5 200
19 176 15 1 8 200
20 153 36 0 11 200
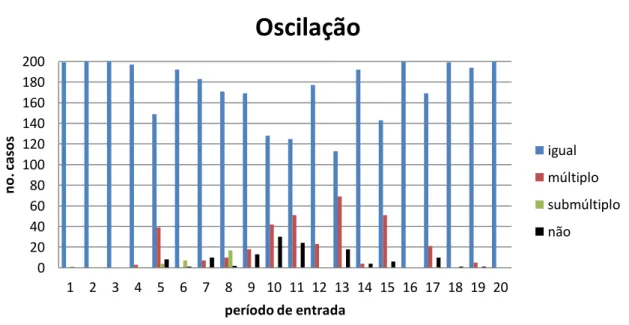
total geral 3500 343 29 128 4000
média 175,0 17,2 1,5 6,4
desvio padrão 11,7 10,9 2,3 2,4
4.2.1 Oscilações
A Tabela 4.5 mostra os resultados obtidos. A faixa verde contém os casos em que as partes oscilaram com o mesmo período. A grande maioria das simulações está contida nessa faixa. A faixa azul destaca os casos em que a parte 2 oscilou com o dobro do período da parte 1. O número desses casos é muito menor que os de mesmo período. São ainda menos frequentes os casos em que o período da parte 2 é o triplo do período da parte 1. A faixa vermelha clara realça os casos em que a parte 2 não oscilou, que representam 3% do total de observações. A grande maioria desses casos ocorreu quando a parte 1 também não oscilou. São escassos os casos em que a parte 2 oscila em período menor que a parte 1 (menos de 1%). O comportamento típico da rede apresenta as partes oscilando no mesmo período.
Gráfico 4.5 – Relação entre o período de oscilação da parte 2 e o período da parte 1, por período da entrada. N=200, q=5%.
O Gráfico 4.5 mostra a relação entre os períodos das partes, em função do período da entrada. Nos menores períodos de estímulo, assim como no período de 20 passos, só foram observados casos de igual período. Para casos de período de entrada maiores, as redes devem oscilar no mesmo período, igual ao período do estímulo de entrada, devido a período de inatividade. Nos períodos intermediários, observaram-se casos de multiplicidade, que atingiram o maior percentual dos casos no período de 13 passos – 35% dos casos. No período 10, observou-se a maior ocorrência de casos em que a parte 2 não oscilou – 23% dos casos.
0 20 40 60 80 100 120 140 160 180 200
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
n
o
.
ca
so
s
período de entrada
Oscilação
igual
múltiplo
submúltiplo
A Tabela 4.6 mostra esses mesmos dados tabulados, com informações de média e desvio padrão por categoria de oscilação. Apenas o período 13 tem número de casos de oscilações iguais das duas partes fora do intervalo de 2 desvios padrão, ou seja, o número de
casos de “igual” nesse período é um caso extremo numa distribuição normal. Também são dados desviantes os múltiplos desse mesmo período, também afastados da distribuição de dados. Já para os casos sem oscilação, apenas o período de 10 passos está afastado a mais de 2 desvios padrão da média – o período 1 está no limite.
Tabela 4.6 – Número de casos do período de oscilação da parte 2 em relação ao período da parte 1, por período. N=200, q=5%.
contagem oscilação
período igual múltiplo submúltiplo não total
1 199 0 1 0 200
2 200 0 0 0 200
3 200 0 0 0 200
4 197 3 0 0 200
5 149 39 4 8 200
6 192 0 7 1 200
7 183 7 0 10 200
8 171 10 17 2 200
9 169 18 0 13 200
10 128 42 0 30 200
11 125 51 0 24 200
12 177 23 0 0 200
13 113 69 0 18 200
14 192 4 0 4 200
15 143 51 0 6 200
16 200 0 0 0 200
17 169 21 0 10 200
18 199 0 0 1 200
19 194 5 0 1 200
20 200 0 0 0 200
total 3500 343 29 128 4000
média 175,0 17,2 1,5 6,4
desvio padrão 28,5 21,6 4,1 8,8
Os casos mais frequentes são oscilações no mesmo período, conforme ilustrado no Gráfico 4.6.
Gráfico 4.6 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). A parte 1 oscila com período 10, mesmo período da parte 2. O período da entrada é de 5 passos. N=200, q=5%.
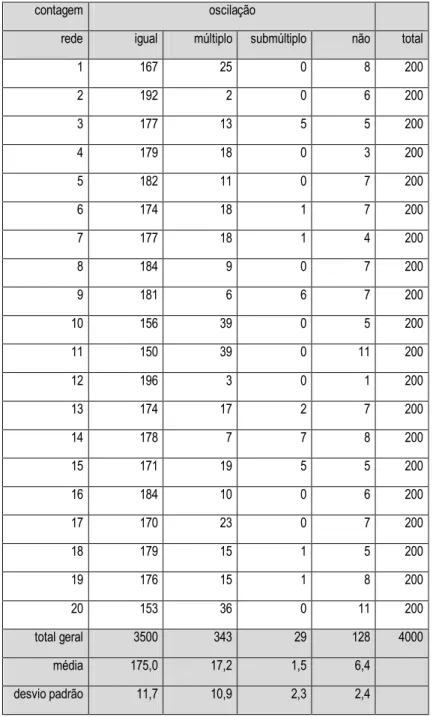
Após esses casos de sincronismo, a parte 2 oscilando com o dobro do período da parte 1 apresenta o maior número de observações. Um caso está retratado no Gráfico 4.7.
Gráfico 4.7 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). A parte 1 oscila com período 10, a parte 2 com período 20. O período da entrada é igual a 5. N=200, q=5%.
O outro grupo com número representativo de casos é o que contém casos em que a parte 2 oscila com o triplo do período da parte 3, como exemplificado no Gráfico 4.8.
Gráfico 4.8 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). A parte 1 oscila com período 10, a parte 2 com período 30. O período da entrada é igual a 5. N=200, q=5%.
Esses três casos ilustrados acima são exemplos de uma mesma rede (a rede 7), sendo estimulada com o mesmo período (período igual a 5 passos de tempo). A única diferença entre as simulações foi a célula que recebe a estimulação periódica: foram, respectivamente,
20 40 60 80 100t 10
20 30 40 x
20 40 60 80 100t 10 20 30 40 50 x
PARTE 1: Período 10 Máximo 0.41 Posição doMáximo 9 PARTE 2:Período 10 Máximo 0.56 PosiçãodoMáximo 33
20 40 60 80 100t 10
20 30 40 x
20 40 60 80 100t 10 20 30 40 50 x
PARTE 1:Período 10 Máximo 0.4 PosiçãodoMáximo 8 PARTE 2:Período 20 Máximo 0.53 PosiçãodoMáximo 31
20 40 60 80 100t 10
20 30 40 x
20 40 60 80 100t 10
20 30 40 x
de baixo pra cima, as 3 primeiras células da coluna mais à esquerda. Essa pequena diferença provocou grande mudança no comportamento da rede. Essa rede está representada na Figura 4.1, com as células de entrada numeradas.
Figura 4.1 – Topologia da rede 7, que possui 5% de ligações adicionais (N=200). Ligações entre células da mesma parte em azul; entre células de partes diferentes em verde. Os números 1, 2 e 3 identificam as células estimuladas que produziram diferentes comportamentos. N=200, q=5%.
Em apenas um caso a parte 2 oscilou com período 1, ou seja, apresentou atividade constante ao longo dos passos. Este caso está ilustrado no Gráfico 4.9.
Gráfico 4.9 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). A parte 1 oscila com período 4 e a parte 2 com período 1. O período de entrada é igual a 1. Esse foi o único caso em que a parte 2 oscilou com período 1. N=200, q=5%.
Podem-se analisar mais detalhadamente os casos em que as partes oscilam no mesmo período. Quando as partes oscilam no mesmo período elas estão sincronizadas, por definição. Pode-se investigar a relação entre o período de oscilação dos neurônios e o período de entrada (o período da estimulação forçada).
4.2.2 Sincronizações
Quando ocorrem sincronizações, isto é, quando o número de células ativas em cada parte oscila no mesmo período, tal período de oscilação pode ocorrer no mesmo período da entrada ou em outro período.
20 40 60 80 100t 10
20 30 40 50 60 x
20 40 60 80 100t 10
20 30 40 50 x
PARTE 1:Período 4 Máximo 0.59 PosiçãodoMáximo 7 PARTE 2:Período 1 Máximo 0.5 Posiçãodo Máximo 9
Gráfico 4.10 – Relação entre o período de oscilação e o período da entrada, por período de entrada. N=200, q=5%.
O Gráfico 4.10 mostra que o tipo de sincronismo varia conforme o período de entrada. Os casos de sincronização no mesmo período de entrada são maioria, seguidos dos casos em que a sincronização ocorre em período que é múltiplo do período de entrada. Nos períodos curtos, não ocorrem sincronizações no mesmo período de entrada. As redes não oscilam em período menor que 4, em caso algum. Nos períodos intermediários, ocorre variedade de comportamentos. Nos períodos maiores, observa-se maior frequência de sincronizações no período de entrada. Do Gráfico 4.11 ao Gráfico 4.13 mostram-se, respectivamente, casos de sincronismo no mesmo período da entrada, em período múltiplo e em período submúltiplo do período de entrada.
Gráfico 4.11 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). Ambas as partes estão sincronizadas com período 8, que é igual ao período de entrada. N=200, q=5%.
0 20 40 60 80 100 120 140 160 180 200
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
no
.
ca
so
s
período de entrada
Sincronismo x período de entrada
igual
múltiplo
submúltiplo
menor
20 40 60 80 100t 5 10 15 20 25 30 35 x
20 40 60 80 100t 10 20 30 40 50 x
PARTE 1: Período 8 Máximo 0.35 PosiçãodoMáximo 7 PARTE 2:Período 8 Máximo 0.53 PosiçãodoMáximo 11
20 40 60 80 100t 10 20 30 40 50 x
20 40 60 80 100t 10 20 30 40 50 x
Gráfico 4.12 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). Partes sincronizadas com período 24, que é múltiplo do período de entrada, que é igual a 8. N=200, q=5%.
Gráfico 4.13 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). Partes sincronizadas com período 4, que é submúltiplo do período de entrada, 8. N=200, q=5%.
Esses três comportamentos diferentes foram produzidos pela mesma rede, provocada com o mesmo período de estimulação. A diferença entre as simulações está nas células estimuladas, indicadas na Figura 4.2.
Figura 4.2– Topologia da rede 19, que possui 5% de ligações adicionais (N=200). Ligações entre células da mesma parte em azul; entre células de partes diferentes em verde. Os números 8,9 e 10 identificam as células estimuladas que produziram diferentes comportamentos.
Em apenas um caso, o período de sincronização não foi múltiplo ou submúltiplo do período de entrada. Nesse caso, com período de entrada 9, as partes oscilaram com período 6. Esse caso está ilustrado no Gráfico 4.14. Nenhum caso das partes oscilando em período maior, e não múltiplo do período de entrada, foi observado.
20 40 60 80 100t 35
40 45 50 x
20 40 60 80 100t 30
35 40 45 50 55 x
PARTE 1:Período 4 Máximo 0.53 Posição doMáximo 29 PARTE 2:Período 4 Máximo 0.57 Posição doMáximo 22
Gráfico 4.14 - Quantidade de células ativas (x) em função do tempo (t) para a parte 1 (gráfico à esquerda) e para a parte 2 (gráfico à direita). Ambas as partes estão sincronizadas com período 6, menor que o período da entrada, que é 9. N=200, q=5%.
A fim de tentar encontrar características das redes que pudessem ser associadas aos diferentes tipos observados de comportamento oscilatório, foram analisadas as seguintes grandezas:
Caminho mínimo médio da rede
Razão entre o período da parte 1 e o período da entrada forçada
Atividade máxima alcançada pela rede
Número de passos em que a rede atinge a atividade máxima
4.2.3 Caminho mínimo médio
As ligações adicionadas diminuem consideravelmente o caminho mínimo médio das redes. Como mostrado no início deste capítulo, essa medida reduz-se em média para 73% da correspondente para um reticulado regular. Os dados de caminho mínimo médio da parte 1 foram separados em tercis e classificados pelos tipos de oscilação.
Tabela 4.7 – Percentuais dos tipos de oscilação pelo tercil do caminho mínimo médio, para a parte 1. N=200, q=5%.
% linha tercil
oscilação 1 2 3 total
igual 35,2% 35,9% 28,9% 100,0%
múltiplo 32,4% 28,6% 39,1% 100,0%
submúltiplo 58,6% 31,0% 10,3% 100,0%
não 31,3% 28,9% 39,8% 100,0%
total 35,0% 35,0% 30,0% 100,0%
20 40 60 80 100t 10
20 30 40 x
20 40 60 80 100t 10
20 30 40 50 x
Como indica a Tabela 4.7, há uma maior participação de redes do primeiro tercil nos casos de submúltiplos do que o esperado. O Gráfico 4.15 evidencia a maior participação de redes com menor caminho mínimo médio nas oscilações submúltiplas, aquelas em que o período da parte 2 é submúltiplo do período da parte 1. Como os tercis têm aproximadamente o mesmo tamanho, seriam esperadas, a priori, alturas de aproximadamente 33% nas participações de cada tercil em cada tipo de oscilação. Pode-se também observar maior participação do terceiro tercil nos casos de múltiplos e naqueles em que a parte 2 não oscila.
Gráfico 4.15 – Participação dos tercis dos caminhos mínimos médios das redes nos tipos de oscilação. N=200, q=5%.
Análises similares foram feitas para os caminhos mínimos médios da parte 2, o que está mostrado na Tabela 4.8.
Tabela 4.8 – Percentuais dos tipos de oscilação pelos tercis do caminho mínimo médio, para a parte 2. N=200, q=5%.
% linha Tercil
oscilação 1 2 3 total
igual 33,9% 36,3% 29,8% 100,0%
múltiplo 44,0% 21,0% 35,0% 100,0%
submúltiplo 20,7% 48,3% 31,0% 100,0%
não 43,0% 34,4% 22,7% 100,0%
total 35,0% 35,0% 30,0% 100,0%
0% 33% 67% 100%
igual múltiplo submúltiplo não
pe
rc
ent
ua
l de
pa
rt
ic
ipa
çã
o
tipos de oscilação
3
2
1
Caminho mínimo médio nos tipos de oscilação