UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
DEPARTAMENTO DE GENÉTICA E BIOLOGIA CELULAR
DANIEL CHAVES DE LIMA
RESPOSTA DE Chromobacterium violaceum CULTIVADA
EM ALTA CONCENTRAÇÃO DE FERRO
RESPOSTA DE CHROMOBACTERIUM VIOLACEUM
CULTIVADA EM ALTA CONCENTRAÇÃO DE FERRO
Dissertação apresentada ao programa de pós-graduação em Bioquímica da Universidade Federal do Rio Grande do Norte como requisito parcial para obtenção do título de Mestre em Bioquímica. Orientador: Silvia Regina Batistuzzo de Medeiros
Como sempre, dedico este trabalho aos meus pais pela excelente educação que me deram, pelo apoio sempre presente e por serem os exemplos pelos
Mais um ciclo se fechando e muitas pessoas foram responsáveis por
este desfecho.
Agradeço, primeiramente, aos meus pais Evanildo e Vanice. Obrigado, por apoiar toda e qualquer decisão que tomo, pelos conselhos experientes e
advertências que tornam minha vida longe de vocês um pouco mais fácil. Amo vocês.
À Mariana por, simplesmente, ser você mesma e continuar comigo. Sempre. Obrigado pela companhia, pelo cuidado, pela preocupação, enfim, por
tudo o que você faz. Je t’aime.
A toda minha família (irmã, tios e tias, primos e primas, avós e avô e sogra), que apesar de estarem distantes, estão sempre torcendo pelo meu
sucesso. Obrigado vovó Alice, pelo exemplo de bondade.
A família Chromo (Raí, Vivi e Jana) pelas contribuições que permitiram a conclusão desse trabalho e, principalmente, Fábio. Mais que um tutor ou um
coorientador, um amigo divertido que me aconselha dentro e fora da vida acadêmica. Espero que nossa parceria continue sempre.
Ao time da faculdade: Ysa, Diego, Manel, Cris e, especialmente, Priscilla, João e Ricardo. Vocês são como irmãos e irmãs e tenho certeza que
sempre poderei contar com o apoio de vocês. Saibam que também podem
contar comigo.
A todos os amigos do LBMG/LAMA pelas ótimas experiências. Muitas
À Prof.ª Silvia Batistuzzo por continuar comigo e acreditar na minha
capacidade e vontade.
A todo o Programa de Pós-Graduação em Bioquímica, especialmente dona Margarita (desculpa por incomodar tanto) e aos professores Hugo e João
Paulo, pelas boas conversas.
À Deus. Apesar de não ser muito religioso, sei que o Senhor cuida de
mim e dos que estão ao meu redor.
Nada faz correr mais o tempo e abrevia mais o caminho do que um pensamento capaz de absorver em si mesmo todas as faculdades mentais daquele que pensa. A vida exterior assemelha-se então a um sono de que esse pensamento é o sonho. Graças a ele, o tempo perde a medida, o espaço perde a distância. Partimos de um lugar e chegamos a outro, e isso é tudo. Do intervalo percorrido, nada permanece em nossa lembrança, a não ser um nevoeiro difuso, no qual se diluem mil imagens confusas de árvores, montanhas e paisagens.
A Chromobacterium violaceum é uma β-proteobactéria Gram-negativa encontrada amplamente em regiões tropicais e subtropicais, cujo genoma foi sequenciado em 2003 mostrando grande versatilidade metabólica e potencial biotecnológico e farmacêutico. Dada à grande quantidade de ORFs relacionadas com o metabolismo de ferro descritas no genoma da C.
violaceum, a importância deste metal para diversos processos biológicos e
devido à carência de dados a respeito das conseqüências do excesso do ferro em organismos de vida livre, é importante estudar o mecanismo de resposta desta bactéria em meio repleto com ferro. Trabalhos anteriores mostram que a
C. violaceum apresenta resistência a grandes concentrações desse metal,
embora ainda não tenha sido descrito qual mecanismo é utilizado para esta sobrevivência. Assim, visando elucidar a resposta da C. violaceum cultivada em alta concentração de ferro e esperando-se obter genes candidatos para serem utilizados em processos de biorremediação, o presente trabalho utilizou a abordagem de proteômica em shotgun e Biologia de Sistemas para avaliar a resposta de C. violaceum cultivada na presença e ausência de 9 mM de ferro. A análise identificou 531 proteínas, sendo 71 expressas exclusivamente pela bactéria cultivada na presença do metal e 100 apenas na condição controle. O aumento na expressão de proteínas relacionadas com o ciclo do TCA possivelmente representa uma reprogramação metabólica na bactéria causada pela grande concentração de ferro no meio. Além disso, foi observado um aumento no ensaio de atividade das enzimas Superóxido dismutase e Catalase, bem como no ensaio de Atividade Antioxidante Total, sugerindo que o metal está induzindo um quadro de estresse oxidativo em C. violaceum, que passou a aumentar os níveis de violaceína e enzimas antioxidantes para melhor se adaptar à condição emergente. Também faz parte da resposta adaptativa a alteração na expressão de proteínas relacionados a transporte, inclusive de ferro, e com a resposta quimiotáxica, o que levaria a bactéria a mudar a orientação de sua locomoção afastando-se do metal. Os dados de Biologia de Sistemas também sugerem uma reprogramação metabólica com mecanismos coordenados por gargalos proteicos envolvidos com transcrição (GreA), metabolismo energético (Rpe e TpiA) e metilação (AhcY).
The Chromobacterium violaceum is a β-proteobacterium Gram-negative widely found in tropical and subtropical regions, whose genome was sequenced in 2003 showing great metabolic versatility and biotechnological and pharmaceutical potential. Given the large number of ORFs related to iron metabolism described in the genome of C. violaceum, the importance of this metal for various biological processes and due to lack of data about the consequences of excess of iron in free-living organisms, it is important to study the response mechanism of this bacterium in a culture filled with iron. Previous work showed that C. violaceum is resistant to high concentrations of this metal, but has not yet been described the mechanism which is used to this survival. Thus, to elucidate the response of C. violaceum cultured in high concentrations of iron and expecting to obtain candidate genes for use in bioremediation processes, this study used a shotgun proteomics approach and systems biology to assess the response of C. violaceum grown in the presence and absence of 9 mM of iron. The analysis identified 531 proteins, being 71 exclusively expressed by the bacteria grown in the presence of the metal and 100 just in the control condition. The increase in expression of proteins related to the TCA cycle possibly represents a metabolic reprogramming of the bacteria caused by high concentration of iron in the medium. Moreover, we observed an increase in the activity assay of superoxide dismutase and catalase as well as in Total Antioxidant Activity assay, suggesting that the metal is inducing oxidative stress in C. violaceum that increases the levels of violacein and antioxidant enzymes to better adapt to the emerging conditions. Are also part of the adaptive response changes in expression of proteins related to transport, including iron, as well as an increased expression of proteins related to chemotaxis response, which would lead the bacteria to change the direction of its movement away from the metal. Systems Biology results, also suggest a metabolic reprogramming with mechanisms coordinated by bottleneck proteins involved in transcription (GreA), energy metabolism (Rpe and TpiA) and methylation (AhcY).
FIGURA 1 Estrutura química da violaceína. Adaptado de: Durán et al. (2007)...17
FIGURA 2 Biossíntese da violaceína. Adaptado de: Ryan et al. (2008)...19
FIGURA 3 Reação de Fenton. Adaptado de Touati, (2000)...23 FIGURA 4. Reação espontânea na qual ocorre a formação do íon ferroso. Adaptado de Harrison e Arosio (1996)...23 FIGURA 5. Reação de Haber-Weiss. Adaptado de Harrison e Arosio (1996)..23 FIGURA 6. Esquema da reação bioquímica envolvida no ensaio para detecção de superóxido dismutase. Fonte: Manual do kit Superoxido Dismutase Assay (706002)...32 FIGURA 7 Curva de crescimento de Chromobacterium violaceum. Cada ponto representa a média de triplicatas biológicas...37 FIGURA 8 Perfil antioxidante em Chromobacterium violaceum, após exposição ao ferro. A) Ensaio de atividade enzimática da enzima catalase. B) Atividade enzimática da enzima Superóxido Dismutase. C) Atividade antioxidante total do extrato protéico total de C. violaceum. Cada coluna representa o valor médio de
três experimentos independentes. * p <
lado...42 FIGURA 12 Medidas de centralidade (Rede Fe-) plotadas em um gráfico Betweenness X Node Degree. Os nós com altos valores em ambos os índices (Bottlenecks) e que possuem grande probabilidade de interligarem processos, estão destacados na figura, com seus respectivos nomes ao lado...42 FIGURA 13 A) Rede de interação de proteínas do grupo Fe+. O input das proteínas foram os dados obtidos a partir das analises de proteômica. Os bottlenecks estão destacados em diferentes cores (ver texto). B e C , representam os clusters obtidos a partir do plugin MCODE...44 FIGURA 14 A) Rede de interação de proteínas do grupo Fe-. O input das proteínas foram os dados obtidos a partir das analises de proteômica. Os
bottlenecks estão destacados em diferentes cores (ver texto). B, C, D, e E
representam os clusters obtidos a partir do plugin MCODE...45 Figura 15 Integração do metabolismo energético em C. violaceum. O aumento da expressão das enzimas com ferro em seu centro ativo está levando a uma maior ativação do ciclo do TCA e do ciclo do Glioxilato. Entretanto, a segunda etapa da glicólise está sendo inibida, enquanto a primeira etapa está mais ativada. Consequentemente está havendo um acúmulo de Gliceraldeído 3-fosfato, que pode estar sendo utilizado no mecanismo de adaptação de C.
violaceum...52
enzimas antioxidantes podem estar sendo sensibilizados pelo ferro livre, o que inibe a produção dessas enzimas, como a tioredoxina; E) Glutationas Transferases, juntamente com a violaceína, podem estar antagonizando o estresse oxidativo causado pelo ferro; F) Para balancear a grande quantidade de ferro no citoplasma, está ocorrendo a diminuição da expressão de proteínas transportadores responsáveis pela entrada do metal, enquanto que outras relacionadas com a saída do ferro estão sendo produzidas em maior quantidade; G) Aumento da resposta quimotáxica induzida pelo ferro. O metal interage com proteínas quimiotáticas aceptoras de metil (MCPs) que transmitem o sinal para o sistema quimiossensório, induzindo a uma mudança na rotação flagelar, o que causa o afastamento da bactéria do ambiente inóspito; H) As setas vermelhas indicam os processos coordenados pelas proteínas bottlenecks (ver discussão)...66 Figura 1 (Apêndice) SDS-PAGE das proteínas totais extraídas de C. violaceum. M = Marcador de peso molecular. 1-3 representam os extratos protéicos totais de C. violaceum cultivada sem adição de ferro; 4-6 representam os extratos protéicos totais de C. violaceum cultivada com adição de 9mM de ferro. As grades mostram os fragmentos que foram excisados para posterior digestão in
1. Introdução ... 16
1.1. Chromobacterium violaceum ... 16
1.2. O ferro e sua importância biológica para os microrganismos ... 21
1.3. Proteômica, Biologia de Sistemas e a Era Pós-genômica ... 24
2. Objetivos ... 29
2.1. Objetivo geral ... 29
2.2. Objetivos específicos ... 29
3. Metodologia ... 30
3.1. Condições de cultivo, curva de crescimento e tratamento com FeSO4 ... 30
3.2. Extração de proteínas ... 30
3.3. Avaliação da atividade antioxidante de C. violaceum ... 31
3.4. SDS-PAGE ... 32
3.5. Digestão in gel e extração dos peptídeos ... 33
3.6. Análise de espectrometria de massas e análise in silico ... 33
3.7. Anotação das proteínas pelo Gene Ontology (GO) ... 35
3.8. Análises de Bioinformática ... 35
3.9. Análises de Biologia de Sistemas ... 35
4. Resultados ... 37
4.1. Crescimento relativo de C. violaceum ... 37
4.2. Perfil antioxidante de C. violaceum ... 37
4.3. Análise de LC-MS/MS ... 38
4.4. Análises de biologia de sistemas ... 41
5. Discussão ... 48
5.1. Metabolismo energético ... 48
5.2. O papel da violaceína ... 52
5.3. Quimiotaxia ... 54
5.4. Proteínas Relacionadas a Transporte ... 55
5.5. Proteínas relacionadas ao estresse ... 56
5.6. Proteínas Hipotéticas ... 58
5.7. Ferric uptake regulator... 59
5.9. Biologia de Sistemas ... 60
6. Conclusões ... 68
7. Considerações finais e perspectivas ... 69
8. Referências ... 70
1. Introdução
1.1. Chromobacterium violaceum
Primeiramente descrita no final do século XIX por Boisbaudran (1882),
Chromobacterium violaceum é uma β-proteobactéria Gram-negativa encontrada amplamente em regiões tropicais e subtropicais. Trata-se de um bacilo móvel, anaeróbio facultativo pertencente à ordem Neisseriales e à família Neisseriaceae (GARRITY e HOLT, 2001). O interesse por esta bactéria vem desde o final da década de 1970 em que diversos estudos foram realizados visando às propriedades terapêuticas de seus produtos metabólicos (CALDAS, 1978; DURAN, 1989; HUNGRIA, 2004). No Brasil, além da Região Amazônica, esta bactéria é encontrada em outros dois ecossistemas: no Cerrado e na Mata Atlântica, de forma que os três ecossistemas representam juntos quase 50% da área de toda região Neotropical (LIMA-BITTENCOURT et
al., 2007).
Com quase quatro décadas de pesquisas realizadas sobre C. violaceum, foi observado que esta bactéria possui grande potencial biotecnológico, com aplicabilidade em várias áreas. Duran e Menk (2001) descreveram, em uma completa revisão, enzimas e metabólitos encontrados nessa bactéria, conferindo a ela inúmeras propriedades industriais e farmacológicas. Por exemplo, a bactéria produz potencializadores de antibióticos como os glicopeptídeos beta-lactâmicos SQ28,504 e SQ28,546 que agem em bactérias Gram-negativas (COOPER et al., 1984; COOPER et al., 1985), além de aerocianidina, um antibiótico que atua tanto em bactérias Gram-negativas quanto em Gram-positivas (PARKER et al., 1988). C. violaceum também produz substâncias que são intermediários do analgésico alfamenina B (ZII, 1984; OHUCHI et al., 1997), o depsipeptídeo antitumoral FR901228, que apresentou citotoxicidade contra várias linhagens de células tumorais como câncer de pulmão, câncer de mama, câncer de cólon e leucemia (NAKAJIMA et
al., 1998). Entre as aplicabilidades industriais podemos citar a síntese de
possuem aplicações médicas e na indústria (KOLIBACHUK et al., 1999; STEINBÜCHEL et al., 1993).
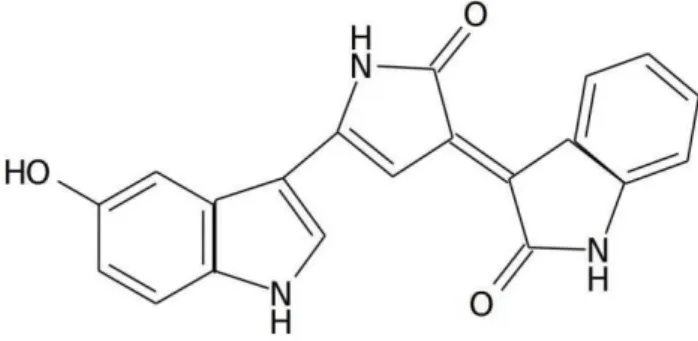
Entretanto, a característica mais notável da C. violaceum é a produção de um pigmento de cor púrpura escuro o qual lhe confere esse nome, a violaceína. Trata-se de um metabólito secundário com massa molecular de 343,3 daltons e que se apresenta insolúvel em água, pouco solúvel em etanol, moderadamente solúvel em dioxano e acetona e apresenta-se totalmente solúvel em DMSO, metanol e acetato de etila (DURAN et al., 2007). Esta substância se decompõe acima de 270ºC e possui três picos de absorbância: 258, 372 e 575 nm (RETTORI e DURÁN, 1998). Além da C. violaceum, outras bactérias produzem esse pigmento, dentre elas a Chromobacterium lividum, a bactéria psicrotrófica RT 102 e a Alteromonas luteociolacea (SHIRATA et al., 1997; NAKAMURA et al., 2003; LAATSCH e THOMSON, 1984). A violaceína teve sua estrutura química primeiramente estudada no fim da década de 1920 (REILLY e PYNE, 1927), e foi isolada pela primeira vez apenas em 1944 (STRONG, 1944). Apesar disso, foi apenas em 1958, através de estudos de degradação e síntese que o composto teve sua estrutura deduzida (BELLATINE et al., 1958), sendo confirmada depois através de análise de espectroscopia (LAATSCH e THOMSON, 1984) (Figura 1).
Figura 1. Estrutura química da violaceína. Adaptado de: Durán et al. (2007).
Diversas funções biológicas foram sugeridas para esse pigmento, entre elas a de que a violaceína seria um pigmento respiratório, uma vez que foi observado um aumento na atividade respiratória quando um extrato de violaceína foi adicionado a uma suspensão de células não-pigmentadas de C.
envolvido na regulação da produção do triptofano, visto que este aminoácido é tóxico para a bactéria em altas concentrações (DEMOSS, 1967). Apesar disso, nenhum desses papéis são aceitos, visto que foi observado que C. violaceum não necessita do pigmento para sobreviver e crescer (SIVENDRA e LO, 1975; DURAN e FALJONI-ALARIO, 1980). Hoje, acredita-se que a violaceína está envolvida na proteção contra radiação, uma vez que a bactéria é amplamente encontrada exposta em águas e solos de regiões tropicais e subtropicais (VASCONCELLOS e CRECZYNSKI-PASA, 2004). Apesar disso, esta hipótese ainda não foi investigada, de forma que, até hoje, ainda não existe nenhum dado experimential que sugira, com segurança, o papel biológico deste pigmento.
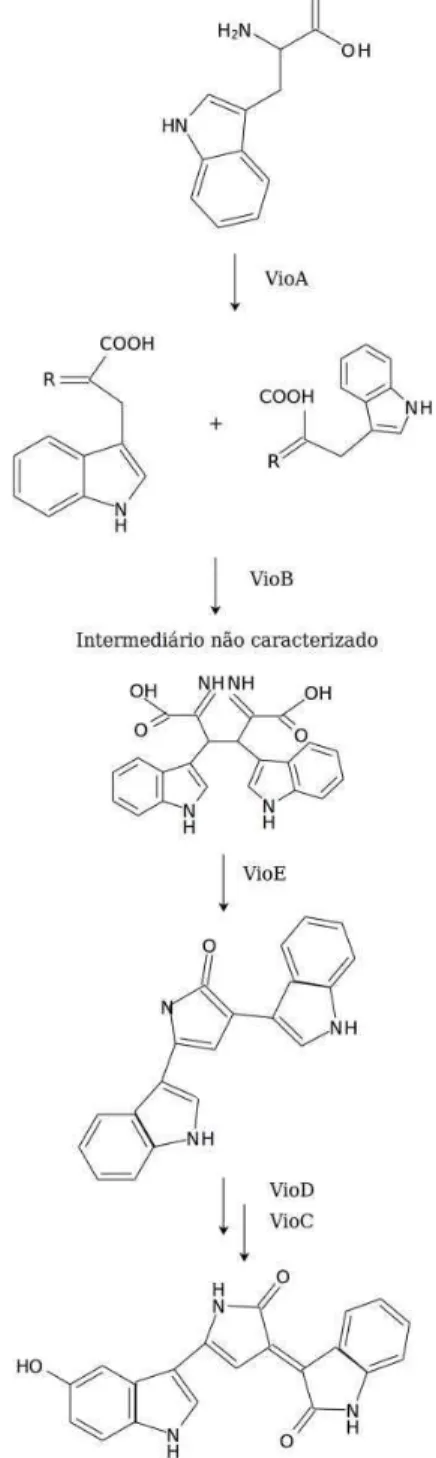
Figura 2. Biossíntese da violaceína. Adaptado de: Ryan et al. (2008).
A violaceína possui grandes atributos farmacológicos como toxicidade contra vários tipos celulares, entre eles leveduras (PEREIRA et al., 2005) e fibroblastos (MELO et al., 2000). Foi visto também que este pigmento apresenta atividade antitumoral com indução de apoptose em células de leucemia mieloide em humanos (MELO et al., 2003; FERREIRA et al., 2004) bem como ação citotóxica em outras linhagens de células tumorais (DURAN et
bactérias Gram-negativas e, principalmente, contra Gram-positivas como a
Mycobacterium tuberculosis (SOUZA et al., 1999; DURAN, et al., 1983; DURAN et al., 1990). A violaceína também foi relacionada com ação antifúngica
(BARRETO et al., 2008), antileishmaniose (LEON et al., 2001), tripanossomicida (DURÁN, 1994) e foi visto que esse metabólito também apresenta atividade antiviral, com a inibição de células infectadas por poliovírus e vírus da herpe (MAY et al., 1991).
Devido ao seu grande valor biotecnológico, o genoma da
Chromobacterium violaceum foi sequenciado pela Rede Nacional de
Sequenciamento Nucleotídico. A linhagem sequenciada foi a ATCC 12472, isolada de águas da cidade de Mentekab, Malásia (HUNGRIA et al., 2005). Esta linhagem foi escolhida por ser a linhagem tipo (SNEATH, 1984) e por ser utilizada em vários estudos no Brasil. O genoma completo de C. violaceum consiste de um único cromossomo circular de 4.751.080 pares de base e uma média de G + C de 64,83%. A bactéria possui 4.431 quadros de leitura aberta (ORFs) e constatou-se que cerca de 40% dessas ORFs não possuem uma função identificada, sendo classificados como hipotéticos ou hipotéticos conservados (VASCONCELOS et al., 2003). Comparando o genoma de C.
violaceum com outros organismos foi vista uma maior semelhança com Ralstonia solanacearum (17,4%), Neisseria meningitidis (9,75%) e
Pseudomonas aeruginosa (9,61%). O aparato genético dessa bactéria permite
espécies reativas de oxigênio (HUNGRIA et al., 2004). Foi visto que C.
violaceum possui genes relacionados com várias vias de reparo de DNA,
incluindo as vias: reparo por reversão direta do dano, reparo por excisão de base, reparo por excisão de nucleotídeo, reparo mismatch, reparo recombinacional e resposta SOS, bem como a presença de genes de reparo duplicados, o que sugere uma defesa mais elaborada contra danos no DNA (DUARTE et al., 2004). C. violaceum possui ainda, inúmeros genes com ação biotecnológica como, por exemplo, biorremediação, visto que foram encontrados operons envolvidos na resistência ao arsênico (AZEVEDO et al.,
2008) e ao cianeto, genes capazes de reduzir compostos halógenos a formas
menos tóxicas e outros genes que podem ser usados na redução de impactos provocados por metal (ouro, por exemplo) em áreas de garimpo, além do controle de pragas na agricultura (CAREPO et al., 2004).
Além disso, foi observada a presença de um grande número de genes em C. violaceum relacionados com o metabolismo de ferro. Entre estes, encontram-se 35 genes envolvidos no transporte e armazenamento desse metal, indicando a sua grande relevância para a bactéria (VASCONCELOS et
al., 2003).
1.2. O ferro e sua importância biológica para os microrganismos
elétrons torna-o um dos principais mediadores das reações de oxido-redução de forma que, sozinho ou incorporado em agregados de ferro-enxofre ou grupos heme, o ferro é indispensável para uma variedade de processos celulares incluindo respiração, ciclo do ácido tricarboxílico, inibição de estresse oxidativo, bem como a síntese de aminoácidos, desoxiribonucleotídeos, lipídeos e esteróis (HASS et al., 2008).
O ferro também é um componente muito importante relacionado ao processo de patogenicidade de microrganismos, uma vez que o processo de multiplicação de bactérias dentro do hospedeiro requer, entre outros nutrientes, uma quantidade mínima de ferro, o que torna este metal um fator limitante para organismos patogênicos (MIETZNER e MORSE, 1994). Dessa forma, todos os microrganismos precisam de ferro, porém a disponibilidade deste é limitada na maioria dos ambientes aeróbios, uma vez que é encontrado somente em complexos minerais insolúveis ou ligados a proteínas com grupos prostéticos de ferro pertencentes a mamíferos ou a plantas (HUNGRIA et al., 2004). Por isso, muitos organismos desenvolveram maneiras eficientes de obter ferro sob condições ambientais desfavorecidas. Um importante mecanismo são os
clusters gênicos que codificam enzimas e receptores que reconhecem
componentes quelantes de ferro com baixo peso molecular, chamados de sideróforos. Esses sideróforos são secretados no ambiente onde se ligam ao ferro no estado férrico com alta afinidade em um mecanismo crucial para aumentar a competitividade e adaptação em diferentes condições ambientais (LANKFORD, 1973; CROSA, 1989).
transporte de ferro, mostrando que esta bactéria está bem adaptada a ambientes contendo este metal.
Embora o ferro em excesso não possua ação genotóxica uma vez que não é capaz de reagir com o DNA, esse metal pode promover a reação de Fenton a qual produz espécies reativas de oxigênio e estes sim têm atividade genotóxica e mutagênica (AROSIO e HARRISON, 1996). Nessa reação, o íon ferroso (Fe2+) não complexado reage com o peróxido de hidrogênio, que também não possui efeito direto sobre o DNA. Durante a reação (Figura 3), ocorre a oxidação do íon ferroso e a decomposição do peróxido de hidrogênio, levando à produção de radicais hidroxilas que podem causar danos a qualquer macromolécula biológica (TOUATI, 2000).
H2O2 + Fe2+ Fe3+ + OH● + OH
-Figura 3. Reação de Fenton. Adaptado de Touati, (2000).
Outra reação envolvendo o ferro e que pode levar, indiretamente, à formação de espécies reativas de oxigênio é a Reação de Haber-Weiss (HABER e WEISS, 1934). Esta é a soma de duas reações: a Reação de Fenton e outra reação espontânea na qual o superóxido reduz o íon férrico (Fe3+), de forma que há a produção do íon ferroso mais oxigênio (Figura 4). Por sua vez, o Fe2+ pode reagir com o peróxido de hidrogênio, produzindo radicais hidroxila que têm potencial de reagir com o DNA e outras biomoléculas como proteínas e lipídeos (Figura 5).
O2●- + Fe3+ Fe2+ + O2
Figura 4. Reação espontânea na qual ocorre a formação do íon ferroso. Adaptado de Harrison
e Arosio (1996).
O2●- + H2O2 + O2 + OH- + OH●
Dessa forma, quando o ambiente está com um excesso de ferro, pode ocorrer uma desregulação no metabolismo desse metal, levando a um aumento da concentração intracelular do íon ferroso na bactéria, que por sua vez pode provocar as reações supra citadas, levando à produção de radicais hidroxila que poderão reagir diretamente com o material genético levando a uma instabilidade genômica.
1.3. Proteômica, Biologia de Sistemas e a Era Pós-genômica
Com os avanços da tecnologia, cresce, a cada dia, o número de organismos que tiveram seu material genético totalmente sequenciado. Em junho de 2012, segundo o NCBI (http://www.ncbi.nlm.nih.gov), os genomas de mais de 12.400 procariontes já estavam disponíveis nos bancos de dados e esse valor cresce, paralelamente, com os avanços tecnológicos (YATES et al., 2009). Por outro lado, pouco progresso vem sendo feito em acessar toda essa informação visto que o conhecimento de todos os genes de um determinado organismo não é suficiente para entender sua fisiologia. Dessa forma, uma nova área de estudo surge na era pós-genômica: a Genômica Funcional. Esse campo utiliza abordagens e técnicas que estudam além da função do gene, sua interação e relação com metabólitos, mRNA, proteínas entre outros (SAITO e MATSUDA, 2010). Um dos alvos da Genômica Funcional é o trascriptoma, o conjunto de todos os genes expressos por um organismo em uma determinada condição. A Trascriptômica é a área responsável por esse estudo e utiliza, principalmente, técnicas de DNA Microarray para determinar a regulação transcricional dos genes através de seus mRNAs (SÁNCHEZ-PLA et al., 2012).
O estudo da proteômica inclui duas áreas distintas de pesquisa: a expressão das proteínas e as suas interações (BLACKSTOCK e WEIR, 1999). O principal objetivo de se estudar a expressão de proteínas é estabelecer mapas quantitativos e qualitativos de expressão de proteínas em condições específicas como diferentes estímulos ambientais e estados de doença, por exemplo (TOMANEK, 2011). Esta abordagem é especialmente útil em estudos durante desenvolvimento de drogas ou estudos toxicológicos em que é de interesse traçar todo o perfil do proteoma em resposta a alguma perturbação fisiológica e identificar proteínas biomarcadoras cujos níveis de expressão possam ser usados para diagnosticar doenças ou avaliar a eficiência de drogas (BAHOU, 2012; HAN et al., 2012). Os dados gerados por esta abordagem da proteômica permitem atingir objetivos como identificar as proteínas envolvidas em rotas metabólicas relacionadas aos diferentes processos celulares; identificar novos alvos farmacológicos e marcadores biológicos, relacionados ao processo de estabelecimento e progressão de doenças; a identificação de moléculas bioativas a partir de extratos biológicos naturais, levando ao desenvolvimento de novos fármacos; caracterização das respostas celulares a determinadas drogas, doenças e mudanças ambientais (SILVA et al., 2007). Já a abordagem de interação de proteínas cuida da caracterização específica de interações proteína-proteína e da formação de complexos proteicos (SAITO e MATSUDA, 2010). A identificação de interações entre proteínas facilita o estudo de processos específicos, vias metabólicas e leva ao desenvolvimento de mapas de interação para vários tipos de células e condições, aumentando nosso entendimento das redes intracelulares (SAITO e MATSUDA, 2010; BLACKSTOCK e WEIR, 1999).
coordenadamente, permitem a sobrevivência de um organismo a uma determinada condição (ALBERT, 2005). Estas interações, por sua vez, podem ser representadas graficamente por meio de redes de interações. Neste diagrama, os componentes (DNA, RNA, Proteína, entre outros) são designados como “nós” que se comunicam com os outros componentes por meio de “conectores”, que por sua vez podem especificar um ponto de início e de fim ou podem não especificar nenhuma direção (ALBERT, 2005).
Apesar de os diagramas de interações terem a sua importância, também é necessário compreender o sentido, o fluxo, o padrão ou qualquer outra informação contida nestas interações (KITANO, 2002). Assim, alguns índices estatísticos auxiliam as análises de Biologia de Sistemas. Por exemplo, os nós de uma determinada rede podem ser caracterizados pela quantidade de conexões que faz com outros nós adjacentes, o que é conhecido como Node
Degree (ALBERT, 2005). Nós com altos valores de Node Degree são referidos
como “hubs” e estes, caso sejam removidos do organismo (por meio de mutações, por exemplo), podem ser letais dada a grande quantidade de processos que interliga (ARMSTRONG et al., 2006). Outro conceito importante em Biologia de Sistemas é o de Betweenness. Originalmente introduzido por Freeman (1977) como uma nova medida de centralidade para redes sociais,
Betweenness refere-se, de maneira simplista, à menor distância existente entre
um par de nós. A maioria dos “caminhos” mais curtos dentro de uma rede passa por nós com altos valores de betweenness, de forma que estes nós tornam-se pontos centrais na comunicação entre processos biológicos (YU et
al., 2007). Por fim, um dos conceitos mais importantes dentro da Biologia de
Todos esses parâmetros mencionados acima fornecem informações importantes que tornam as análises de Biologia de Sistemas uma poderosa ferramenta para o estudo de complexos biológicos, permitindo uma visão global e dinâmica dos dados fornecidos a partir de técnicas de alto rendimento, além de identificar os principais componentes regulatórios de um dado mecanismo fisiológico.
Dessa forma, vários estudos utilizando abordagens da genômica funcional vêm sendo realizados para investigar as conseqüências tanto do excesso quanto da carência de ferro em microrganismos (CRESTANI et al., 2012; MIYAMOTO et al., 2009; LAFRENTZ et al., 2009; WINTERS et al., 2008; EBANKS et al., 2004). Estes trabalhos utilizam, em sua maioria, abordagem
bottom-up de proteômica que traz resultados similares do efeito de
desequilíbrios na homeostasia do ferro, principalmente para organismos patogênicos. Em contrapartida, poucos trabalhos são realizados investigando as conseqüências do excesso de ferro em microrganismos de vida livre. Estudos prévios em nosso laboratório mostram que a C. violaceum apresenta ampla capacidade de resistência e tolerância a metais, inclusive ao ferro, quando comparadas com E. coli (LEAL, comunicação pessoal; BRITO, comunicação pessoal). De acordo com Leal, C. violaceum apresenta capacidade de sobrevivência prolongada em concentrações de até 8 mM de ferro, o que é uma concentração superior a que maioria de outros microrganismos suportam. Além disso, dados do genoma mostram que C.
violaceum possui 50 ORFs relacionadas ao metabolismo de ferro, o que, em
ensaio foi realizado para verificar a ocorrência de estresse oxidativo, ou identificar qual padrão protéico está sendo utilizado como resposta da bactéria a esta concentração de ferro.
Assim, utilizando a Proteômica em shotgun e a Biologia de Sistemas como ferramentas, procuramos entender a resposta da Chromoabcterium
violaceum cultivada em 9 mM de ferro, contribuindo para o entendimento da
2. Objetivos:
2.1. Objetivo geral
Elucidar a resposta de Chromobacterium violaceum cultivada em alta concentração de ferro.
2.2. Objetivos específicos
• Verificar o padrão de crescimento de C. violaceum cultivada em 9 mM de ferro;
• Mensurar a atividade antioxidante da bactéria por meio da quantificação das enzimas catalase e superóxido dismutase, bem como analisar a atividade antioxidante total da bactéria na presença e ausência do ferro; • Identificar o padrão de expressão proteica de C. violaceum cultivada na
presença e ausência de ferro;
• Identificar, por meio de Biologia de Sistemas, os principais bottlenecks responsáveis pela resposta da bactéria ao ferro;
3. Metodologia
3.1. Condições de cultivo, curva de crescimento e tratamento com FeSO4
Colônias isoladas de Chromobacterium violaceum cepa ATCC 12472 foram inoculadas previamente em meio Luria Bertani (LB) líquido por 16-18 horas a 28 ºC e sob agitação de 200 rpm.
O tratamento foi realizado em Erlenmeyer contendo o cultivo prévio de bactéria e meio LB líquido (1:10) em um volume final de 100 mL. A solução de FeSO4 foi previamente filtrada em filtros Acrodisk GSF GHP 0,2 µm PALL e
adicionada em uma concentração final de 9 mM. O controle negativo consistiu apenas de C. violaceum mais meio de cultura. A duração do tratamento foi de 4 horas sob as mesmas condições mencionadas acima. O controle negativo e a condição experimental do tratamento foram realizados em triplicata biológica.
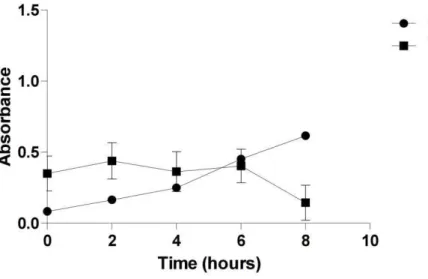
Para comparar o padrão de crescimento da bactéria no meio com e sem adição do metal, foi realizada uma curva de crescimento. Dessa maneira, o tratamento supracitado foi realizado e a densidade óptica (OD) com absorbância em 720 nm foi verificada nos seguintes tempos nas duas condições: 0h, 2h, 4h, 6h e 8h. Todos os experimentos foram realizados em triplicata biológica.
3.2. Extração de proteínas
3.3. Avaliação da atividade antioxidante de C. violaceum
Com intuito de avaliar se o tratamento com ferro altera o nível de estresse oxidativo em Chromobacterium violaceum, a atividade das enzimas antioxidantes catalase (CAT) e superóxido dismutase (SOD) foi mensurada no extrato protéico total na 4ª hora de crescimento utilizando os kits: Catalase Assay Kit 707002 e Superoxide Dismutase Assay Kit 706002 (Cayman Chemical, Ann Arbor, MI) de acordo com as recomendações do fabricante. A concentração protéica das amostras foi determinada pelo método de Bradford e utilizada para normalizar a atividade enzimática detectada.
O kit utilizado para catalase (Catalase Assay Kit) fundamenta-se na função peroxidásica da catalase para a determinação da atividade enzimática. O método é baseado na reação da enzima com metanol, na presença de uma concentração ótima de H2O2. O metanol que serve de substrato para atividade peroxidásica da catalase não é utilizado por outras peroxidases. O formaldeído produzido é medido espectrofotometricamente utilizando-se o 4-amino-3-hidrazino-5-mercapto-1,2,4-triazol (Purpald) como cromógeno. O Purpald reage especificamente com aldeídos e o produto dessa reação, ao sofrer oxidação, origina um produto de coloração roxa. A atividade é mensurada lendo-se a absorbância em 540 nm.
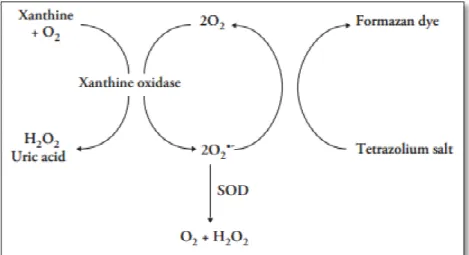
Para a dosagem dos níveis de SOD o kit utiliza um sal tetrazólio para a detecção de O2• gerados pela xantina oxidase e hipoxantina, sendo uma
unidade de SOD definida como a quantidade de enzima necessária para dismutar 50% do O2•. O ensaio avalia todos os três tipos de SOD (Cu/Zn, Mn e
Figura 6. Esquema da reaçã
dismutase. Fonte:
O ensaio de Ativid
Antioxidant Assay kit da
ensaio é a formação do r peróxido de hidrogên etilbenzotiazolina-6-ácido cromógeno de cor verde
Os antioxidantes dependentemente da co de agentes antioxidante TroloxTM, um análogo d padrão.
3.4. SDS-PAGE
Os extratos protéic (1976) e 20 µg de prot poliacrilamida sob condiç utilizado foi o Precision gel foi corado usando o C
ação bioquímica envolvida no ensaio para detecção
te: Manual do kit Superoxido Dismutase Assay (706
vidade Antioxidante Total (AAT) foi realizad da Sigma-Aldrich (CS0790). O princípio u o radical mioglobina ferril a partir da metm
ênio, o qual oxida o ABTS (2, ido sulfônico), produzindo o radical catiônic e detectável espectrofotometricamente a 4 tes suprimem a produção do radi concentração de forma que quanto maior ntes, menor a intensidade colorimétric da Vitamina E, foi utilizado na prepara
téicos totais foram quantificados pelo métod roteínas de cada amostra foram resolvid
dições desnaturantes (SDS-PAGE) a 12% n Plus ProteinsTM WesternCTM Standard d o Coomassie Coloidal da Sigma-Aldrich.
ão de superóxido
06002).
zado utilizando o o utilizado neste tmioglobina e do (2,2’-azino-bis(3-nico ABTS·+, um a 405 nm.
dical catiônico ior a quantidade trica emitida. O ração da curva
3.5. Digestão in gel e extração dos peptídeos
Após o final da eletroforese, cada lane foi excisado em nove fragmentos conforme a densidade de proteínas. A digestão dos fragmentos foi realizada de acordo com a adaptação do protocolo de Shevchenko e colaboradores (1996). Para remover o corante e o SDS, os fragmentos foram lavados três vezes em solução de acetonitrila (ACN) 50% e bicarbonato de amônio 10 mM. Em seguida, os géis foram desidratados em ACN a 100%, reduzidos com ditiotreitol (DTT) 10 mM à temperatura ambiente e alquilados com iodacetamida (IAA) 50 mM em ambiente escuro. Os fragmentos foram lavados mais uma vez com bicarbonato de amônio 100 mM. Novamente os fragmentos foram desidratados com ACN 100% e reidratados com bicarbonato de amônio 100 mM. Após mais uma desidratação com ACN a 100%, os géis foram hidratados em solução de tripsina (Trypsin Gold, Mass Spectrometry Grade da Promega [v5280]), preparada de acordo as instruções do fabricante. Foram aplicados de 35-50 µL de tripsina em banho de gelo. Em seguida, foi adicionado bicarbonato de amônio 50 mM, o suficiente para cobrir os géis que foram incubados a 37ºC por 16-18 horas.
Para extrair os peptídeos, foram adicionados de 10-30 µL de ácido fórmico 5% sobre os géis. Após incubação de 10 minutos em temperatura ambiente, o sobrenadante contendo parte dos peptídeos foi transferido para outro tubo. Em seguida, foi adicionada a segunda solução de extração (ácido fórmico 5% e ACN 50%) em um volume suficiente para cobrir os géis que foram incubados por 10 minutos, em temperatura ambiente. O sobrenadante mais uma vez foi transferido para o tubo previamente separado, contendo os peptídeos já extraídos. A extração de peptídeos feita com a segunda solução foi realizada mais uma vez. Finalmente, a solução contendo os peptídeos digeridos foi concentrada em Concentrator Plus da Eppendorf.
3.6. Análise de espectrometria de massas e análise in silico
BEH130-C18 (100µm × 100 mm) em um fluxo de 600 nL/min. O gradiente foi de 2-98% de ACN em ácido fórmico de 0,1% durante 45 minutos. O modo de operação do instrumento foi o “top three”. Neste modo, um espectro MS é adquirido seguido pelo MS/MS dos três mais intensos picos detectados. Os espectros foram adquiridos utilizando o programa MassLynx v.4.1 e os arquivos de extensão raw foram convertidos em formato de lista de picos (mgf) utilizando o Mascot Distiller 2.2.1.0, 2008, (Matrix Science Ltda) contra o banco de dados de proteínas não-redundantes de Chromobacterium violaceum ATCC 12472 baixados do NCBI em 24 de Janeiro de 2012. A busca utilizou como modificação fixa a carbamidometilação, oxidação de metionina como modificação variável, uma clivagem perdida pela tripsina e tolerância de erro de massa de 0.1 Da, tanto para o íon precursor ou para o fragmento. Apenas peptídeos com score significativo (p < 0,05) calculado pelo Mascot foram analisados. Após a análise inicial do Mascot, os resultados obtidos de todas as três replicatas de cada condição foram concatenados utilizando o modo merge do programa.
3.7. Anotação das proteínas pelo Gene Ontology (GO)
As categorias funcionais das proteínas foram obtidas com o programa Blast2GO (http://www.blast2go.org) (CONESA et al., 2005). Nessa análise, a ferramenta escolhida usa as sequências FASTA das proteínas que são alinhadas usando-se a ferramenta BLAST contra os bancos de dados do NCBI. Todos os parâmetros utilizados estavam no modo default. Assim, com base em similaridade de sequências, o programa realizou a anotação das proteínas nos três domínios do Gene Ontology: Função Molecular (F), Processo Biológico (P) e Componente Celular (C). Gráficos de Pizza foram gerados pelo Blast2GO representando as anotações do GO.
3.8. Análises de Bioinformática
As comparações das sequências protéicas foram realizadas por meio da ferramenta BLAST (ALTSCHUL et al., 1990). As proteínas hipotéticas e/ou hipotéticas conservadas detectadas no trabalho foram reanotadas, a fim de verificarmos se estas continuavam com funções indefinidas. Aquelas que continuaram sem função estabelecida mesmo após o alinhamento, tiveram seus domínios funcionais identificados no Conserved Domain Database (CDD) (MARCHLER-BAUER et al., 2011).
3.9. Análises de Biologia de Sistemas
experimentais, um índice de baixa confidência (0.150) foi utilizado para gerar as redes, permitindo um maior número de interações entre os nós. Todos os métodos de predição foram ativos: Neighborhood, Gene Fusion,
Co-occurrence, Co-expression, Experiments, Databases e Textmining.
As redes montadas foram exportadas e subsequentemente analisadas no Cytoscape 2.8.2 (SMOOTH et al,. 2010). Para selecionar as subredes de proteínas mais relevantes dentro do interatoma, o plugin do Cytoscape MCODE (Molecular Complex Detection) v. 1.32 foi utilizado (BADER e HOGUE, 2003). Os parâmetros da análise do MCODE foram: inclusão de loops; degree cutoff 2; opção haircut ativada (o que leva a deleção do cluster de nós com uma única conexão); opção de fluff ativada; node score cutoff de 0.2; K-core 2 e maximum
depth de 100. Apenas os clusters gerados com índice acima ou igual a 2.5
foram usados.
Os cálculos de centralidade das redes foram computados a partir das redes e topologias locais. Os nós Bottlenecks das redes foram identificados por meio do gráfico Betweenness X Node Degree com valores gerados pelo
plugin CestiScaPe 1.21. Betweenness indica a extensão em que um nó
específico está entre todos os outros nós dentro de uma rede e, em geral, mostra a influência de um nó sobre a propagação da informação dentro da rede (NEWMAN, 2005; FELTES et al., 2011). Node Degree corresponde à quantidade de conexões que um nó específico faz com outros nós adjacentes. Altos índices de Node degree são denominados “hubs” (YU et al., 2007). Dessa forma, um nó específico com altos valores de Node Degree e de Betweenness representa um Bottleneck, ou seja, uma proteína que interliga muitos processos biológicos (FELTES et al., 2011)
4. Resultados
4.1. Crescimento relativo de C. violaceum
Com o objetivo de avaliar a interferência do ferro no crescimento de C.
violaceum, foi realizada uma curva de crescimento com e sem adição desse
metal (Figura 7). Foi observada uma evidente diferença no crescimento a partir de 4 horas de cultivo, indicando que o ferro está inibindo o crescimento populacional da bactéria impedindo-a de alcançar a fase log de crescimento.
Figura 7 Curva de crescimento de Chromobacterium violaceum. Cada ponto representa a média de triplicatas biológicas.
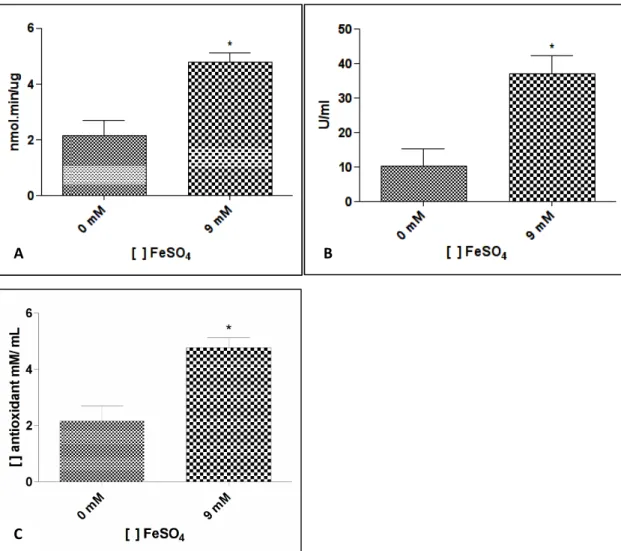
4.2. Perfil antioxidante de C. violaceum
Visando verificar se a concentração de ferro estabelecida para a proteômica gerava um estresse oxidativo, as atividades enzimáticas da catalase e superoxido dismutase foram mensuradas, assim como foi realizada uma avaliação da atividade antioxidante total do extrato protéico de C.
violaceum.
Figura 8. Perfil antioxidante em Chromobacterium violaceum, após exposição ao ferro. A) Ensaio de atividade enzimática da enzima catalase. B) Atividade enzimática da enzima Superóxido Dismutase. C) Atividade antioxidante total do extrato protéico total de C. violaceum. Cada coluna representa o valor médio de três experimentos independentes. * p < 0,05.
4.3. Análise de LC-MS/MS
As proteínas totais foram resolvidas em SDS-PAGE seguida de digestão
in gel (Apêndice, Figura 1) com posterior identificação por espectrometria de
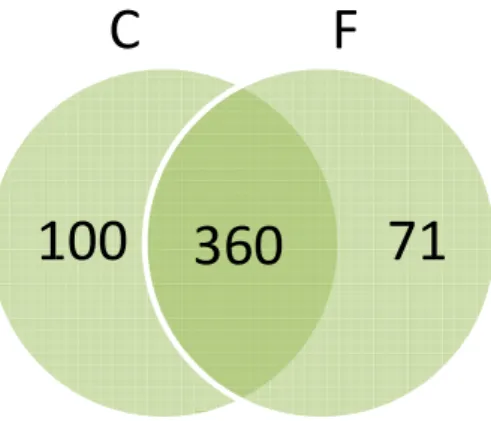
massas. A análise feita no programa Scaffold identificou um total de 531 proteínas (Figura 9). Desse total, 100 proteínas são exclusivas da condição controle e 71 exclusivas da condição tratada com ferro. Das 360 proteínas encontradas em ambas as condições, 61 aumentaram a sua expressão enquanto que 124 diminuíram a expressão na presença de ferro.
A
C
Figura 9 Diagrama de Ven condição. C
A anotação das pr pela ferramenta Blast2G representação de proteín diante, para melhorar apareceram apenas no t aumentaram a express mencionadas como prote e as que diminuíram a ex Entre as proteínas destacou foi Transporte relação às proteínas do g de proteínas foram os p biossíntese de aminoácid
10
enn mostrando a quantidade de proteínas detecta C = Controle negativo; F = Tratamento com ferro.
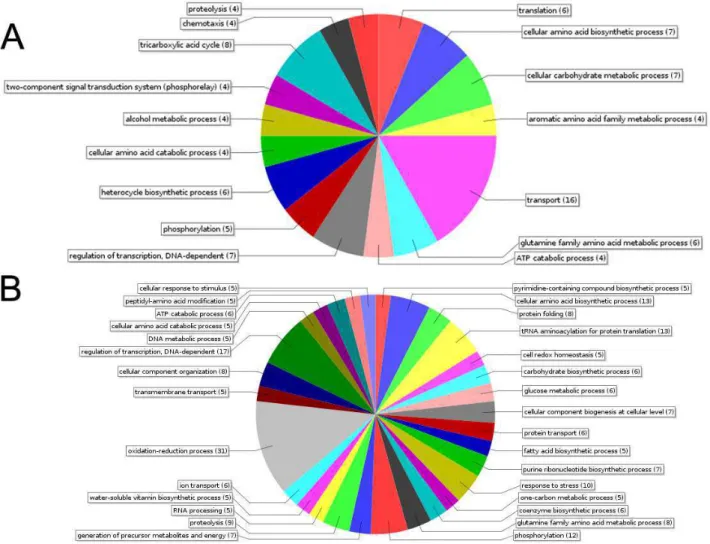
proteínas nas categorias do Gene Ontolog t2GO. Após a anotação, as categoria eínas foram também fornecidas pelo softw r a interpretação dos resultados, as p o tratamento com o ferro (71), juntamente
ssão após a exposição com o meta oteínas da condição Fe+. As exclusivas do expressão (124) serão as da condição
Fe-as da condição Fe+, o processo biológico rte e Ciclo do Ácido Tricarboxílico (Figu o grupo Fe-, os processos biológicos com
processos de oxirredução, regulação da cidos (Figura 10B).
100
360
71
C
F
tadas em cada
logy foi realizada rias com maior tware. Daqui por proteínas que nte com as que tal (61), serão do controle (100)
-.
Figura 10 Distribuição dos principais processos biológicos (P) detectados na análise de proteômica. A) Classificação das proteínas que aumentaram a expressão ou surgiram após o tratamento. B) Classificação das proteínas que diminuíram a expressão ou desapareceram após o tratamento.
Dentre as proteínas da condição Fe+ que merecem destaque, encontram-se proteínas do metabolismo energético, principalmente enzimas pertencentes ao ciclo do Ácido Tricarboxílico (TCA) (Apêndice, Tabela 1). Muitas enzimas dessa via, além de proteínas exclusivas do ciclo do glioxilato tiveram a sua expressão aumentada ou surgiram após o tratamento. De maneira contrária, proteínas da glicólise deixaram de ser expressas, sugerindo que o ferro está induzindo a uma reprogramação metabólica em C. violaceum.
da condição Fe-, o que poderia ser um indício de algum mecanismo provocado pelo ferro estar levando à repressão dessas enzimas (Apêndice, Tabelas 2 e 3).
O grupo de proteínas com maior representação nas condições Fe+ e Fe-foi o das proteínas hipotéticas (Apêndice, Tabela 4). Após a identificação inicial das proteínas pelo Mascot (22 ORFs hipotéticas em Fe+ e 28 em Fe-), foi realizada uma reanotação das proteínas hipotéticas por meio de alinhamento com BLAST, a fim de se verificar quais destas ainda continuavam com função desconhecida. Mesmo após a reanotação, um número considerável de proteínas ainda continuou como hipotéticas (22 na condição Fe+ e 26 na Fe-). Apenas as proteínas hipotéticas que tiveram a expressão aumentada e/ou que surgiram após o tratamento com o ferro serão discutidas nesse trabalho.
4.4. Análises de biologia de sistemas
intercelulares. Apesar da análise do MCODE ter separado os interatomas maiores em sub-redes, muitos nós são compartilhados entre os clusters.
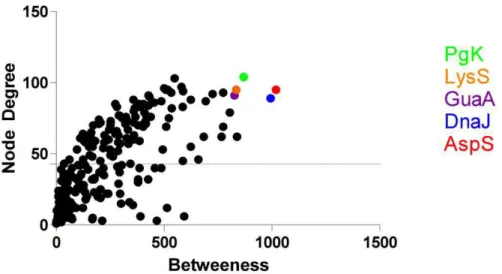
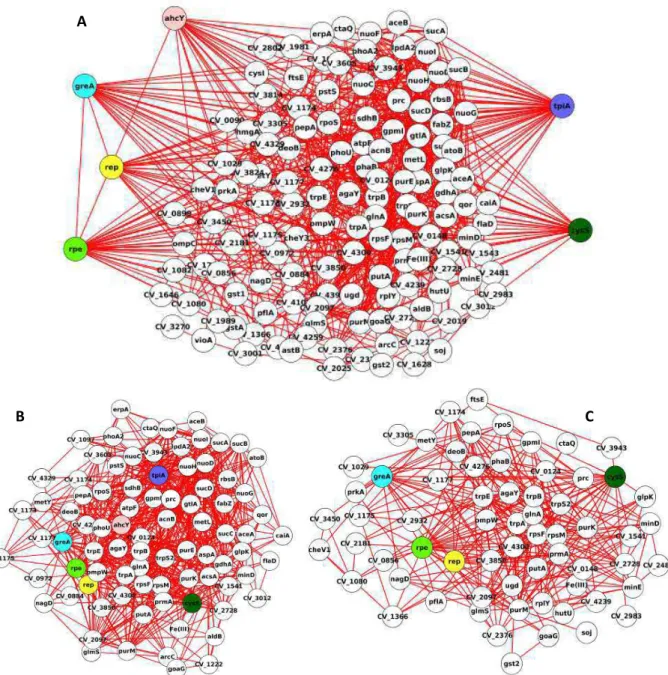
Os índices de centralidade produzidos pelo plugin CestiScaPe permitiram a identificação dos nós centrais no controle da comunicação entre processos biológicos, os bottlenecks (Figuras 11 e 12). Os bottlenecks de cada rede estão destacados nas Figuras 13 e 14, onde também é possível observar os nós e conectores das redes principais e dos clusters.
Figura 11 Medidas de centralidade (Rede Fe+) plotadas em um gráfico Betweenness X Node
Degree. Os nós com altos valores em ambos os índices (Bottlenecks) e que possuem grande
probabilidade de interligarem processos, estão destacados na figura, com seus respectivos
nomes ao lado.
Figura 12 Medidas de centralidade (Rede Fe-) plotadas em um gráfico Betweenness X Node
probabilidade de interligarem processos, estão destacados na figura, com seus respectivos
nomes ao lado.
Figura 13. A) Rede de interação de proteínas do grupo Fe+. O input das proteínas foram os
dados obtidos a partir das analises de proteômica. Os bottlenecks estão destacados em
diferentes cores (ver texto). B e C , representam os clusters obtidos a partir do plugin MCODE.
A
A
C
Figura 14A) Rede de interação de proteínas do grupo Fe-. O input das proteínas foram os
dados obtidos a partir das analises de proteômica. Os bottlenecks estão destacados em
diferentes cores (ver texto). B, C, D, e E representam os clusters obtidos a partir do plugin
MCODE.
5. Discussão
O presente trabalho é um dos primeiros estudos de proteômica realizados em Chromobacterium violaceum. O único trabalho, até o momento, a fazer uma análise protéica em larga escala de C. violaceum foi o de Baraúna e colaboradores (2011) que investigaram os efeitos do cianeto sobre a bactéria. Entretanto, a técnica utilizada por eles foi o 2D-SDS-PAGE que é de baixa resolução e poucas proteínas foram identificadas. A abordagem shotgun permite a identificação de um grande número de proteínas, o que facilita o entendimento das respostas fisiológicas (YATES et al., 2009). Os processos biológicos que foram mais representados nas categorias do Gene Ontology serão discutidos de maneira independente a fim de se entender a sua relação com a resposta adaptativa da bactéria frente ao ferro.
5.1. Metabolismo energético
As principais vias metabólicas que tiveram suas enzimas mais, ou exclusivamente, expressas na condição Fe+, foram aquelas relacionadas ao metabolismo energético, sugerindo uma reprogramação metabólica em resposta ao acúmulo do ferro.
para evitar o acúmulo de ferro livre no citoplasma. Também relacionado ao ciclo do TCA, as proteínas citrato sintase (GtlaA), α-cetoglutarato desidrogenase (SucA) e Succinil-CoA sintetase (SucC) também aumentaram a sua expressão em decorrência do ferro, sugerindo que não apenas as proteínas da via com o metal em sua composição aumentam mas também aquelas que não o possuem. Assim, provavelmente devido ao aumento na produção de intermediários do ciclo do TCA produzidos pelas ferro-enzimas, todo este ciclo está mais ativo, em consequência de regulação alostérica causada pelos intermediários, que são substratos para etapas seguintes da via.
O ciclo do Glioxilato é uma via metabólica na qual algumas de suas enzimas fazem parte do ciclo do ácido tricarboxílico. Apesar disso, este ciclo não acontece em mamíferos devido a ausência de duas proteínas exclusivas dessa via, a Isocitrato Liase (AceA) e a Malato sintase (AceB), sendo encontrado apenas em alguns invertebrados, plantas e microrganismos (ENSIGN, 2006). Pieper e colaboradores (2010) ao estudarem os efeitos da carência de ferro na bactéria patogênica Yersinia pestis observaram uma diminuição na expressão de enzimas tanto da via do ciclo do TCA quanto do Glioxiltato. Em C. violaceum foi observado o aparecimento da malato sintase e um aumento na expressão da isocitrato liase na condição Fe+. Esta via é um desvio do ciclo do TCA, sendo responsável pela síntese de succinato a partir de duas moléculas de Acetil-CoA. Além disso, permite a assimilação de carbono a partir de compostos C2 como o acetato o que pode levar a gliconeogênese ou outros processos biosintéticos (ENSIGN, 2006; DUNN et
al., 2009). AceA e AceB, juntamente com outras enzimas do ciclo do Glioxilato,
metabólica, poderia também estar relacionado com o sucesso da patogenicidade da bactéria, visto que o cultivo com o ferro está reprimindo seu crescimento como é possível observar no gráfico de crescimento (Figura 7).
Ainda relacionado ao metabolismo energético, foi observado o surgimento de uma subunidade da enzima NADH desidrogenase (NuoD). Esta enzima faz parte do complexo NADH: ubiquinona oxirredutase, responsável pela oxidação do NADH, contribuindo para o gradiente de prótons da cadeia respiratória. Como as outras enzimas mencionadas acima, a NuoD possui em seu centro catalítico um núcleo de ferro e enxofre, o que justificaria o aumento em sua expressão na condição Fe+.
A glicólise é a principal via catabólica da glicose. A degradação da glicose até o piruvato faz parte do metabolismo energético central da maioria dos sistemas biológicos, com grande conservação de rotas entres os domínios
Eukaryota e Bacteria, divergindo em algumas minúcias em Archaea
G3F, enquanto que a oxidação deste intermediário até o piruvato e, consequentemente, até o Acetil-CoA (etapa catalisada pela Piruvato desidrogenase – não detectada em níveis significantes) está diminuindo. Dessa forma, o ciclo do TCA estaria perdendo a principal via que o alimenta. Assim, a
C. violaceum necessitaria de vias anapleróticas para alimentar este ciclo, o que
pode ser observado pelo aumento na expressão de enzimas como a Acetil-CoA sintetase (AcsA), Glutamato desidrogenase (GdhA) e Aspartato Amônia-Liase (AspA) que produz Acetil-CoA, α-cetoglutarato e fumarato, respectivamente. A não-utilização do gliceraldeído-3-fosfato no restante da glicólise pode estar relacionada com o direcionamento deste composto para alguma outra função, provavelmente relacionada com a adaptação da bactéria a condição de estresse. O aparecimento da enzima Glicerol kinase (GlpK), que produz G3F a partir de glicerol e ATP, na condição Fe+ é mais um indício de que C.
violaceum necessita aumentar os níveis dessa molécula para realizar alguma
outra função que não o metabolismo energético usual. As proteínas envolvidas no metabolismo de lipídios foi outro grupo de proteínas que, apesar de pouco representadas, foram encontradas na análise do perfil protéico. A Acil-CoA desidrogenase (CaiA), enzima chave no catabolismo de ácidos graxos, apareceu em C. violaceum na condição Fe+, indicando um aumento da beta-oxidação, processo no qual há uma grande produção de acetil-CoA, o que seria mais uma via anaplerótica que estaria alimentando o ciclo do TCA. Em contrapartida, foi observada uma diminuição de enzimas relacionadas com a biossíntese de lipídios. A acetil-CoA carboxilase catalisa a etapa inicial na formação de malonil-CoA, reação irreversível no anabolismo de ácidos graxos. Quatro subunidades desta enzima (AccA-D) tiveram a sua expressão diminuída ou desapareceram na condição Fe+. Esses resultados mostram que C.
violaceum está mobilizando sua reserva lipídica, provavelmente para alimentar
Figura 15 Integração do meta das enzimas com ferro em seu e do ciclo do Glioxilato. Entret primeira etapa está mais Gliceraldeído 3-fosfato, que p violaceum.
5.2. O papel da violaceín
A violaceína é um que está associada com
al., 2006; LOPES et al
etabolismo energético em C. violaceum. O aumen seu centro ativo está levando a uma maior ativaçã retanto, a segunda etapa da glicólise está sendo in s ativada. Consequentemente está havendo u e pode estar sendo utilizado no mecanismo de
eína
um metabólito secundário produzido pela om diversas propriedades farmacológicas
al., 2009; DURÁN et al., 2010). No ent
ento da expressão ção do ciclo do TCA inibida, enquanto a um acúmulo de e adaptação de C.
fisiológico do pigmento no contexto bacteriano ainda não foi completamente elucidado.
As etapas e os genes necessários para a biossíntese da violaceina (e outros intermediários com grupo indol) foram revisados recentemente por Hoshino (2011). Cinco genes são responsáveis por produzir a violaceína (VioA-E) além de etapas não enzimáticas de autooxidação (Hoshino, 2011). Em nosso trabalho, um desses genes teve a sua expressão aumentada (VioE) após o tratamento com o ferro, enquanto que o VioA foi detectado exclusivamente nesta condição, embora o VioA também esteja sendo expresso no grupo controle, visto que esta enzima é essencial na biossíntese da violaceína e o pigmento também estava sendo produzido na ausência de ferro. Genes relacionados com a biossíntese do aminoácido triptofano também foram representados na condição Fe+ (TrpA e TrpE), sugerindo um aumento nas vias que convergem para a produção do pigmento violaceína. Esse resultado sugere que a violaceína pode ter um papel na sobrevivência de C. violaceum frente às condições de estresse gerado pelo excesso de ferro.
De fato, um papel antioxidante foi proposto para a violaceína por Konzen e colaboradores (2006). Nesse trabalho foi possível observar que o pigmento se incorpora à membrana plasmática, o que protegeria a bactéria contra peroxidação lipídica. Além disso, foi observado que a violaceína é capaz de varrer (scavenge) diversas espécies reativas de oxigênio, incluindo o radical hidroxila. Este pode ser produzido pela Reação de Fenton e constitui uma das espécies reativas de oxigênio mais danosas às biomoléculas (TOUATI, 2000; CORNELIS et al., 2011). Assim, podemos sugerir que o tratamento de ferro está induzindo a formação de radicais hidroxil via Reação de Fenton e que a C.
violaceum está aumentando a produção da violaceína para se proteger do
5.3. Quimiotaxia
O sistema quimiossensório é utilizado por bactérias patogênicas e de vida livre para perceber condições ambientais que podem ou não ser nocivas para o microrganismo (PORTER et al., 2011; LERTSETHTAKARN et al., 2011). O sinal ambiental é captado e transmitido no meio intracelular, controlando a movimentação flagelar o que permite as bactérias mudarem para ambientes com condições adequadas ao crescimento (PORTER et al., 2011).
C. violaceum possui um grande aparato protéico relacionado com o
mecanismo de quimiotaxia já descrito em outros microrganismos (Pereira e colaboradores, 2004). As proteínas quimiotáxicas aceptoras de metil (MCP) são responsáveis por captar o sinal externo que em seguida é transmitido para proteínas de quimiotaxia intracelulares.
Em nosso trabalho, foram detectadas cinco proteínas relacionadas com a resposta quimiotática (CV_0899, CV_1082, CheV1, CheY3 e CheA) bem como uma proteína flagelar (FlaD) que aumentaram e/ou surgiram na condição Fe+, indicando que o ferro no meio está agindo como um repelente para a bactéria que, dessa maneira, utiliza a movimentação flagelar para buscar ambientes menos hostis. As MCPs são responsáveis por receber o sinal externo que é transmitido por proteínas acopladoras do sistema quimiossensório. As proteínas acopladoras podem ser as CheW e/ou CheV. Esta última está também associada com a capacidade de adaptação da quimiotaxia que permite os microrganismos reiniciar os sinais do ambiente, impedindo a saturação do sistema (ALEXANDER et al., 2010). A proteína CheA, quando fosforilada, doa o seu grupo fosfato para CheY que se difunde para o motor flagelar, modificando a rotação do flagelo o que é responsável pela mudança direcional da bactéria (PORTER et al., 2011). A proteína CheY tem um papel crucial nessa resposta visto que a sua interação com o flagelo controla a frequência com que a bactéria muda de direção enquanto nada (WADHAMS et al., 2004).
gradiente eletroquímico de H+ ou Na+ entre a membrana citoplasmática (SOWA e BERRY, 2008). De fato, está havendo um acúmulo de prótons no espaço inter-membranas, visto que as subunidades de ATP-sintase estão deixando de ser expressas enquanto que a cadeia transportadora de elétrons está continuamente sendo alimentada pelo ciclo do TCA. Maurer e colaboradores (2005) mostram que em condições de ácidas, ocorre uma diminuição na expressão de ATP-sintase como mecanismo para evitar que o acúmulo de prótons no citoplasma acidifique, ainda mais, o meio, de forma que esta diminuição na ATP-sintase também pode ter um papel direto na regulação do pH. A NuoD também pode estar contribuindo para esse gradiente de prótons. Dessa forma, o gradiente gerado pode estar sendo utilizado para a movimentação flagelar orientada pela resposta quimiotáxica.
5.4. Proteínas Relacionadas a Transporte
Juntamente com as proteínas hipotéticas e as do metabolismo geral, o grupo com maior representação foi o das proteínas relacionadas ao transporte, principalmente as do sistema ABC. O seqüenciamento do genoma de C.
violaceum mostrou a presença de uma grande quantidade de genes que
codificam proteínas transportadoras de ferro, além de ORFs hipotéticas que possuem domínios possivelmente relacionados com o transporte desse metal (VASCONCELOS et al., 2003). Grangeiro e colaboradores (2004) sugerem que a grande quantidade de genes de transporte em C. violaceum provavelmente está relacionada com o fato de esse microrganismo habitar um grande número de ambientes, o que facilitaria a sua adaptação a diversas condições ambientais.
presentes na condição Fe+, embora não possamos garantir a sua interação com os transportadores ABC.
A Ferric binding protein (FbpA) é uma proteína periplasmática que serve de domínio auxiliar do transportador ABC FbpABC e tem um papel crucial na entrada de ferro em microrganismos patogênicos (SIBURT et al., 2012; WEAVER et al., 2010). Esta proteína deixou de ser expressa em C. violaceum após a exposição ao ferro. Devido à grande quantidade desse metal no meio, a inibição da expressão de proteínas relacionadas com a captação de ferro é uma estratégia eficiente na sobrevivência do organismo, associada com a incorporação de Fe livre a metaloproteínas.
As outras proteínas de transporte detectadas estão associadas com outros tipos de substâncias como aminoácidos e peptídeos (CV_2481, CV_2983, CV_4053, CV_4329) e carboidratos (CV_1197), o que seria esperado em um quadro de reprogramação metabólica na qual as concentrações necessárias de cada nutriente seriam modificadas.
5.5. Proteínas relacionadas ao estresse
Como já mencionado, o ferro é capaz de reagir com o peróxido de hidrogênio, produzindo o radical hidroxila, uma espécie reativa de oxigênio capaz de danificar biomoléculas (ATACK e KELLY, 2009). Dessa forma, o tratamento com ferro em C. violaceum pode estar induzindo o estresse oxidativo via Reação de Fenton. Esse quadro foi comprovado quando verificamos a atividade antioxidante total (AAT) em C. violaceum que foi significativamente maior no extrato protéico Fe+ em comparação com o Fe- (Figura 8C). Assim, podemos inferir que o excesso de ferro no meio está levando a uma maior produção de ROS e a bactéria, por sua vez, está aumentando as suas defesas antioxidantes, o que pode ser observado quando comparamos a AAT dos dois extratos protéicos.
oxidativo (ALSCHER et al., 2002). De fato, houve um aumento significativo da atividade de SOD no extrato protéico da condição Fe+ quando comparado com o controle negativo (Figura 8B). Outra evidência que reforça a existência de estresse oxidativo, foi o aumento observado na expressão da Sodb2 na condição Fe+, embora sem significância estatística. Apesar disso, esse pequeno aumento na expressão, provavelmente foi suficiente para elevar a atividade de SOD nos níveis observados no ensaio.
Dessa forma, como consequência do aumento da atividade de SOD, estaria ocorrendo um aumento na produção de peróxido de hidrogênio, que é um produto da dismutação do superóxido. Assim, é de se esperar uma elevação na atividade da enzima catalase (KatE), que de fato, foi observado na condição Fe+. Apesar disso, os níveis da enzima diminuíram significativamente na condição experimental, o que nos leva a questionar o que está causando o aumento da atividade observado. Provavelmente, alguma enzima similar à catalase está sendo responsável pelo aumento da atividade de detoxificação, visto que o ensaio utilizado mensura a decomposição do peróxido de hidrogênio. Além disso, Atack e Kelly (2009) ao revisarem as conseqüências de estresse oxidativo para a bactéria Gram-negativa Campylobacter jejuni afirmaram que a catalase, em muitos organismos, não é o principal meio de remover o peróxido de hidrogênio. Outras peroxidases, bem como o piruvato, estão associadas com a capacidade de reduzir esta ROS até água (MALLET et
al., 2005). Foi visto que reguladores transcricionais estão relacionados com a