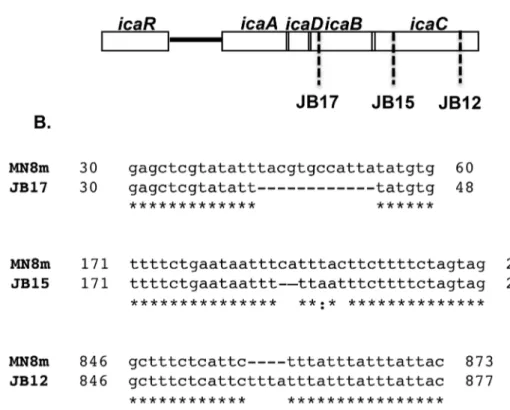
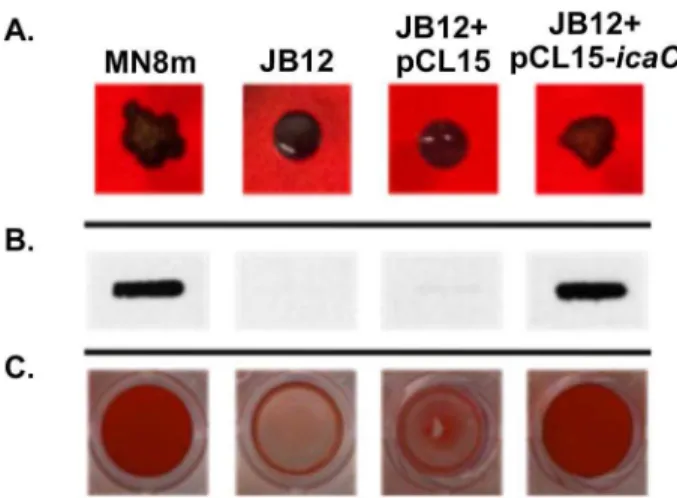
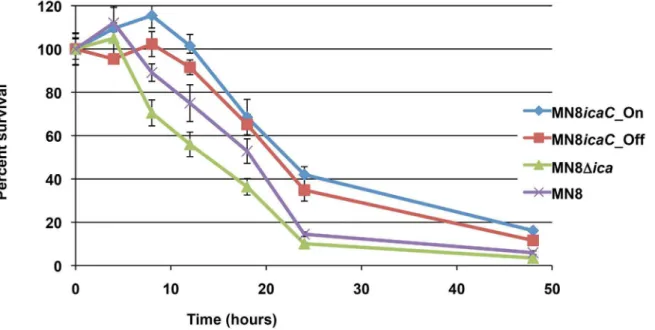
Phase variation of poly-N-acetylglucosamine expression in Staphylococcus aureus.
Texto
Imagem

Documentos relacionados
Em relação aos testes farmacológicos, não é possível afirmar que a fração n-butanólica apresenta atividade hipno-sedativa, uma vez que prolongou o tempo de sono
Taken together, these findings strongly suggest that hypoxia regulates IGF1 expression through HIF1 a , and that upregulated IGF1 induces the activation of IGF1R and increases
Taken together, these results suggest that IL-27 enhances the expression of TRAIL and TLR3 in human melanomas and inhibits their tumor growth in cooperation with poly(I:C), partly in
These results indicate that ellagic acid might be the main active antifungal compound of Lafoensia pacari and further suggest that the mode of antifungal action of these extracts
Se considerarmos o momento improvisacional como impulso-intuitivo, sensível e o estrutural como racional pois implica capacidade de repetição rigorosa, nos
Our results suggest that about 1.8% of expressed sugarcane genes be- long to the chaperone category, and our identification of a high level of chaperone expression confirms
Taken together, these results suggest that elevated expression of H19/IGF2 is caused by a specific chromatin structure that is regulated by the DNA methylation status of IGF2 DMRs
O tema da migração internacional de goianos para os Estados Unidos da América, em especial para o estado da Califórnia e para a cidade de San Francisco, tem sido tema e
