Identification of RegIV as a novel GLI1 target gene in human pancreatic cancer.
Texto
Imagem
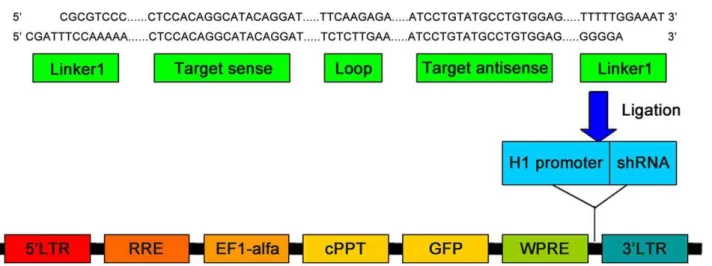
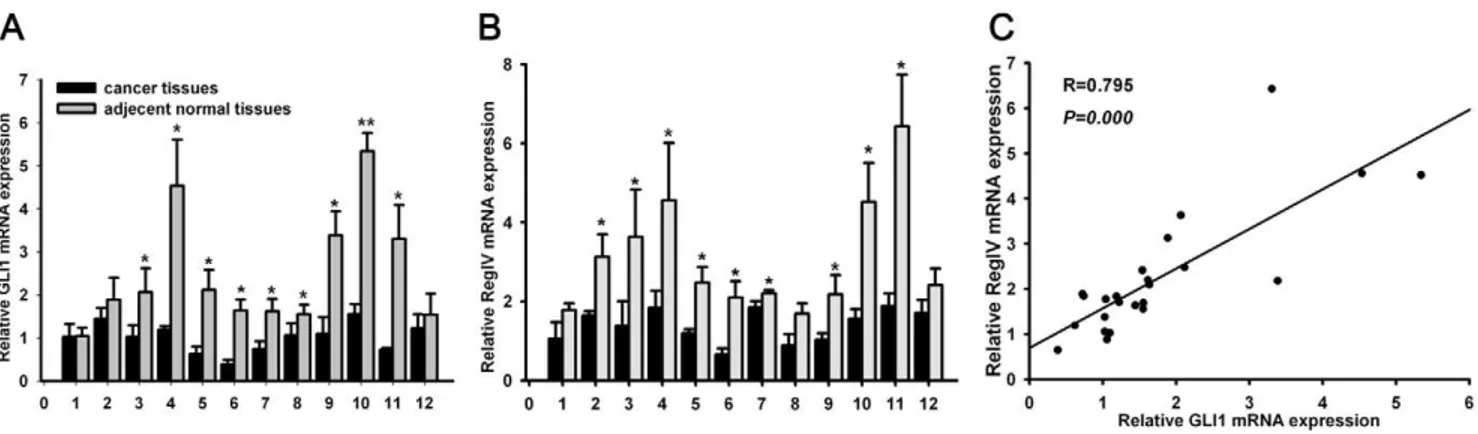
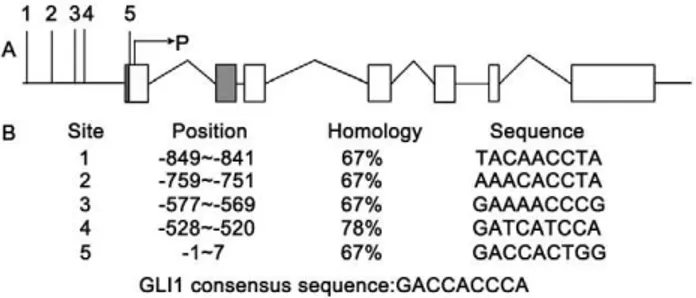
Documentos relacionados
Conforme os dados demonstrados no Gráfico I, o corpus da pesquisa que deu origem a este artigo confirmou uma tendência da toponímia brasileira como um todo, ou seja, o predomínio
Comparing the sequence of the hexon gene fragment amplified by our set of primers with sequences of other human adenoviruses present in the Gene- Bank, we conclude that this region
Polymorphism in the promoter region of the mannose- binding lectin gene among human T-cell lymphotropic virus infected subjects. Sexual transmission of human
)GF R were identified as the regulated target gene in previous studies ,. )t is possible that the discrepancies in the functions of miR‐ in different types of cancer
De acordo com a problemática referida, são considerados dois objectivos para o presente estudo: clarificar diferentes conceitos e concepções decorrentes da evolução do discurso
Multiple sequence alignments of the ribC gene demonstrated that these two cats from the north-eastern region of Brazil were infected with the same strain recently
The TFSEARCH and MAPPER platforms were used to predict the effects that the two novel polymorphisms in the promoter of the UGT1A9 gene would have on the tran- scription factor
In the present study, the conformation- sensitive gel electrophoresis (CSGE) method was applied to the screening of the Sp1-PBS- and TIS-containing portion (~0.5 kb) of the
