A new method for the study of essential fatty acid requirements in fish larvae
Sofia Morais* and Luı´s E. C. Conceic¸a˜o
CCMAR, Universidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal
(Received 11 July 2008 – Revised 3 September 2008 – Accepted 3 September 2008 – First published online 30 October 2008)
This study describes a methodology with potential application in the estimation of essential fatty acid (EFA) requirements of fish larvae. Senegalese sole (Solea senegalensis) larvae were fed, from 16 days after hatching (DAH), on Artemia enriched with different oils, inducing graded dietary concentrations of DHA: (1) soyabean oil, containing no measurable amounts of DHA (NDHA); (2) fish oil, inducing a medium DHA level (MDHA, 3 g DHA/100 g fatty acids); and (3) a mixture of Easy DHA Selco and Microfeed, resulting in high DHA content
(HDHA, 8 g/100 g). At 28 DAH a metabolic trial was conducted where larvae were tube fed [1-14C]DHA, in order to determine its absorption,
retention in the gut and body tissues, as well as its oxidation. At 23 DAH the HDHA treatment induced a significantly higher larval growth, while
at 32 DAH significant differences were only found between the NDHA and HDHA treatments. The absorption of tube-fed [1-14C]DHA was
extre-mely high (94 – 95 %) and independent of feeding regime. However, in larvae fed NDHA Artemia, a significantly higher amount of label was retained in the gut compartment and a concurrently lower retention was measured in the body. A significantly higher proportion of the absorbed DHA label was oxidized in larvae fed HDHA, compared to NDHA. Based on these results, we suggest that increasing dietary supply of DHA above the larval requirement level results in its increased oxidation for energy purposes and we propose potential applications of the tube feeding methodology using radiolabelled EFA in conjunction with dose – response studies.
Essential fatty acids: Fish larvae nutrition: Requirements: Metabolism
Marine fish resemble all other studied vertebrates in the fact that they need a dietary supply of PUFA, particularly DHA (22 : 6n-3), EPA (20 : 5n-3) and arachidonic acid (20 : 4n-6) for their normal growth and development(1). These are essen-tial fatty acids (EFA) for marine fish, which cannot biosynthe-size them de novo nor from shorter-chain precursors such as 18 : 3n-3 or 18 : 2n-6(1,2). Their biochemical, cellular and phys-iological functions are the same in fish as in other vertebrates, where they have a central role in maintaining the structural and functional integrity of cell membranes and as precursors of important metabolites, such as eicosanoids(1). DHA, in par-ticular, has high biological value as one of the major PUFA constituents of cell membranes and is the most abundant fatty acid (FA) in the neural and visual tissue of fish(1,3,4). Fish tissues, in general, are even richer in DHA than mamma-lian tissues and fish brain lipids are characterized by a higher degree of unsaturation and increased percentages of DHA and EPA, compared to the rat(3,4). Dietary deficiencies of DHA leading to a depletion of DHA in the phospholipids of the rod cells of the retina have been associated with a decreased ability to capture preys at low light intensity in juvenile her-ring, denoting an impaired vision and/or neural function(4). On the other hand, neural or visual malfunctioning may be one of the factors responsible for the abnormal malpig-mentation which is common in cultured flatfish larvae(2,5).
Membranes of the melanophores determining pigmentation may also be directly affected by DHA deprivation(2). Finally, dietary deficiencies of DHA have been related to poor growth, survival, abnormal behaviour and higher susceptibility to stress and disease (i.e. reduced immunity) of several fish larvae species(6 – 8).
The need to provide adequate dietary levels of DHA for early developing fish with unusually high growth rates (10 – 100 % per d) and large developmental demands(9) is therefore unquestionably one of the highest priorities in larval nutrition, which is reflected in the large number of studies focusing on the determination of DHA requirements of different fish species(1,2). Dose – response approaches are commonly used in mammalian and juvenile and adult fish nutrition studies but have been very difficult to adapt to marine fish larvae, given that they are initially fed on live prey with a variable and extremely difficult to control nutri-tional composition. However, the development of new pro-ducts and live food enrichment technologies(10) has recently enabled the use of dose – response designs to study the EFA requirements of marine fish larvae(8,11,12). Studies examining the biological response of fish larvae to key indicators of larval performance such as survival, growth, pigmentation and stress resistance, to feeding live prey (mostly rotifers and Artemia) enriched with graded concentrations of the
* Corresponding author: Dr Sofia Morais, Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, UK, fax þ 44 1786 472133, email sofia.morais@stir.ac.uk
Abbreviations:DAH, days after hatching; EFA, essential fatty acids; FA, fatty acids; HDHA, high DHA; IAA, indispensable amino acids; MDHA, medium DHA;
NDHA, no dietary DHA.
qThe Authors 2008
British
Journal
of
Nutrition
https://www.cambridge.org/core . Universidade do Algarve , on 12 Feb 2019 at 13:38:40, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms
.
EFA of interest or with a gradient of EFA ratios (e.g. DHA:EPA or EPA:arachidonic acid) will likely become more common. However, these parameters might not be the most sensitive to determine an exact EFA requirement.
In mammals, in order to determine an indispensable amino acid (IAA) requirement, the indicator amino acid concept is commonly employed, resulting in very accurate estimates(13). This method is based on the idea that when one IAA is deficient for protein synthesis, then all other IAA (including a labelled indicator) will be oxidized. However, when increas-ing amounts of the limitincreas-ing IAA are included in the diet, the oxidation of the indicator IAA will decrease, until no further change is observed in the indicator oxidation, signifying that the requirement for the limiting IAA is met(13). EFA, on the other hand, and DHA in particular, given their prominent structural role, are mostly retained in body tissues and are a poor substrate for the energy-generating b-oxidation system(2). Therefore, in the present study it was hypothesized that an increase in the dietary supply of DHA above larval requirements would result in its increased catabolism or oxi-dation, enabling in this way an accurate and quick estimation of DHA dietary requirements. In order to test this assumption, Senegalese sole (Solea senegalensis Kaup, 1858) larvae were fed Artemia enriched with different oil emulsions resulting in three different DHA concentrations. The effects on growth were analysed and a complementary metabolic trial was additionally performed, enabling the determination of not only the total absorption and tissue retention of14C-labelled DHA that was tube fed but also its metabolic fate (i.e. how much was oxidized for the production of energy).
Materials and methods
Larval rearing and experimental diets
Larvae were obtained from IPIMAR Aquaculture Research Centre (Olha˜o, Portugal) at 14 days after hatching (DAH), with an average wet weight of 3·73 (SD 0·17) mg (n 5,
twenty pooled larvae). Until this age they had been fed Artemia enriched in a mixture of microalgae (Nannochloropsis sp. and Isochrysis sp.). At this time the larvae had already settled and were transferred to a recirculation system of nine 3-litre flat bottom trays, which were stocked with 200 larvae each. During two acclimation days all trays were fed Artemia enriched for 12 h, at 250 nauplii/ml, with a mixture of Easy DHA Selco (INVE Aquaculture NV, Dendermonde, Belgium; two doses, at 0 and 9 h, according to manufacturer’s instruc-tions) and Microfeed (EWOS, Bathgate, UK; 0·4 g/l in the same two doses). From 16 DAH onwards the larvae were fed one of three experimental treatments, in triplicate trays: (1) Artemia enriched on a soyabean oil emulsion, containing no dietary DHA (NDHA); (2) Artemia enriched on a fish oil emulsion, a medium DHA regime (MDHA); and (3) Artemia enriched on a mixture of Easy DHA Selco and Microfeed, containing a high DHA level (HDHA). The HDHA Artemia was enriched in the conditions described earlier, while the enrichments using soyabean oil and fish oil emulsions were conducted at a density of 150 nauplii/ml, during 16 h, with 0·6 g/l being added in two doses (0·4 g/l at 0 h and 0·2 g/l at 8 h). A single batch of enriched Artemia from each treatment was produced and was kept frozen for the duration of the
experiment. Larvae were fed frozen enriched Artemia, after thawing in seawater, in excess to satiation, three times daily. Photoperiod was 12 h light – 12 h dark, salinity was kept at 33 and temperature varied between 20·5 and 228C. At 23 and 32 DAH, twenty and twenty-four larvae, respectively, were collected from each tray, flash frozen in liquid nitrogen and later freeze-dried in a Savant VLP120 ValuPump (Savant Instruments Inc., Holbrook, NY, USA), for the deter-mination of individual dry weight.
Fatty acid analysis
Triplicate samples of Artemia from each treatment were col-lected into liquid nitrogen, for the determination of FA composition. In addition, at 32 DAH, a pool of twenty larvae was collected from each tray and stored in liquid nitro-gen until FA analysis. Lipids were extracted with a chloro-form – methanol (2:1, v:v) mixture(14). The FA methyl esters were obtained by transesterification with 1 % sulphuric acid in methanol and were purified by adsorption chromatography on NH2 sep-pack cartridges (Waters, S.A., Milford, MA, USA)(15), and separated and quantified by GLC following the conditions described by Izquierdo et al.(16). FA methyl esters were identified by comparison to external standards.
Tube feeding experimental set-up and metabolic trial A tube feeding trial was conducted at 28 DAH, using the methodology and experimental set-up described by Rønnestad et al.(17)and Morais et al.(18), to examine the absorption and metabolism of a [1-14C]DHA label (ARC American Radio-labelled Chemicals Inc., St Louis, MO, USA; 3·7 MBq/ml), after an incubation period of 24 h. Briefly, in this set-up each larva is placed in a separate 20 ml scintillation vial (Sigma-Aldrich, St Louis, MO, USA) containing 8 ml sea-water. The incubation vials are sealed well and connected to metabolic trap vials containing 5 ml KOH (0·5M) that retain the 14CO2 resulting from oxidation or catabolism, which is gradually released from the incubation water after it has been acidified with 1 ml HCl (0·1M).
A mixture containing the label was prepared by adding the [1-14C]DHA to fish oil, to obtain a specific activity of 222 dpm/nl (or 3·7 Bq/nl), followed by evaporation of all traces of solvent by flushing with N2. The mixture was then stored at 2 208C until use. On the day before the trial, 30 min after the last meal of the day, six larvae were removed from each triplicate tray and pooled in smaller trays that were transported to the radioisotope experimental room, where the larvae were acclimated and kept unfed overnight. Temperature and salinity were maintained at 228C and 35, respectively, and the trial was conducted under continuous illumination. Before tube feeding approximately 15 000 Artemia metanauplii of each treatment were added to the corresponding tray and larvae were allowed to feed for 30 min. Twelve larvae from each treatment were first sedated in 29·4 mg/ml solution of MS-222 (Sigma-Aldrich) and then tube fed 23 nl [1-14C]DHA mixture. Each larva was removed from the incu-bation set-up 24 h after tube feeding, rinsed in clean saltwater, killed instantaneously by spinal cord severing with a scalpel and the gut was separated from the remaining body by dissec-tion. Both compartments were transferred separately to 6 ml
British
Journal
of
Nutrition
https://www.cambridge.org/core . Universidade do Algarve , on 12 Feb 2019 at 13:38:40, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms
.
scintillation vials (Sarstedt, Rio de Mouro, Portugal) and were solubilized in 200 ml 30 % (w/v) hydrogen peroxide (Sigma-Aldrich) at 608C for 24 h. After cooling, 4 ml scintillation cocktail (Ultima Gold XR, Packard Bioscience, Monza, Italy) were added to each 6 ml vial. The scintillation vials con-taining the incubation seawater (including the evacuated label) and the metabolic trap (including the 14CO2 resulting from oxidation) were prepared for scintillation counting by adding 12 or 10 ml, respectively, of Ultima Gold XR(17,18). The samples were counted on a Beckman LS 6000IC (Beckman Instruments Inc., Fullerton, CA, USA) liquid scintillation counter and the results are presented as a percentage of dpm in each compartment, in relation to the total counts.
Statistical analysis
The differences between larvae submitted to different dietary treatments, in terms of DHA content and growth, as well as the distribution of label in each compartment after tube feed-ing, were compared through one-way ANOVA, using the software Statistica 6 (StatSoft Inc., Tulsa, OK, USA). The assumption of homogeneity of variance was checked using the Bartlett’s test and a significance level of 0·05 was employed(19). Percentage data from the tube feeding trial was arcsin(x1/2) transformed. When significant differences were found, the Tukey HSD multiple range test was per-formed. All data are given as means and standard deviations.
Results
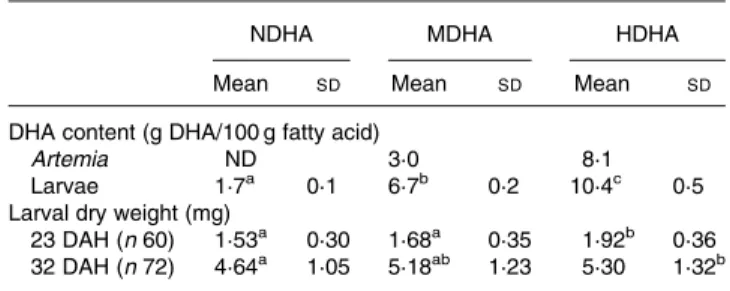
As expected, a gradient of DHA supply was accomplished by the different Artemia enrichments, with Artemia enriched on a soyabean oil emulsion (NDHA) having no measurable amounts of DHA, while the enrichments using fish oil (MDHA) or a mixture of Easy DHA Selco and Microfeed (HDHA) resulted in Artemia containing 3·0 and 8·1 g DHA/ 100 g FA, respectively (Table 1). At 32 DAH, the DHA con-tent of the larvae reflected that of their diet, with significant differences (P, 0·001) being found between the larvae fed different diets (Table 1).
Larval growth, assessed by dry weight, was measured at 23 DAH, i.e. 7 d after start of feeding on the experimental diets,
when significant differences (P, 0·001) were found between the HDHA and the other two treatments (Table 1). At 32 DAH, significant differences were found only between the NDHA and HDHA treatments (P, 0·05; Table 1).
The absorption and metabolic trial conducted at 28 DAH revealed no significant differences in DHA absorption (total tube fed minus evacuated into the incubation water), 24 h after tube feeding. DHA was thus highly absorbed (94 – 95 % of total label tube fed), independently of the feeding regime (Fig. 1). In terms of label retention in the tissues, a signifi-cantly higher amount of label was retained in the gut compart-ment of larvae fed NDHA diet (P, 0·001), compared to the other two treatments, and a concurrently lower retention was measured in the body compartment of larvae fed this diet (P, 0·01), although it was only significantly different from the MDHA treatment (Fig. 1). As for the amount of 14CO2 found in the metabolic trap, corresponding to the oxidized label, significant differences were found between the diets (P¼ 0·001), with a significantly lower amount of label being oxidized by larvae fed the NDHA diet than those fed HDHA, while an intermediate and non-significantly different amount of label was oxidized by MDHA-fed larvae (Fig. 1).
Discussion
The larval growth of Senegalese sole reflected the dietary level of DHA, although significant differences were only found between larvae fed Artemia containing no measurable amounts of DHA (NDHA) and the treatment containing higher levels of DHA (HDHA), after 16 d of feeding on the experimental treatments. It is believed that Senegalese sole larvae probably have lower DHA requirements during their larval stage than other marine fish species(11,20). Villalta et al.(11)hypothesize that the low DHA requirements of Sene-galese sole may be explained by a predominance of EPA rather than DHA in the benthic fauna, which is the natural diet of this species after settlement.
In terms of larval DHA composition, it closely reflected the quantitative dietary supply, as is commonly found in fish larvae and has been described before for Senegalese sole larvae(20). This indicates a high efficiency of absorption and retention of this EFA, as has been shown in the present study (94 – 95 % of total label absorbed, of which 89 – 94 % was retained in the gut and body tissues) and has been demonstrated previously(18). The present results are not surprising, having in mind the essential and structural role of DHA(1,2), and strengthen the idea that marine fish larvae are extremely efficient in utiliz-ing their dietary supply of EFA, possibly through a series of physiological mechanisms such as the high specificity of diges-tive enzymes towards n-3 PUFA(21)and of the presumed higher affinity of cytosolic FA-binding proteins and higher rate of ester-ification of long-chain PUFA, as well as their preferential incor-poration into polar lipids(22,23).
DHA has been formerly considered an ideal quantitative marker of FA absorption, resulting from its minimal degra-dation, synthesis or conversion through elongation – desatura-tion activities(24). This condition is also particularly useful in the type of trial conducted in the present experiment, as it can be reasonably assumed that the radiolabelled tracer found in each compartment corresponds to the FA which was tube fed. In terms of assimilation of the DHA label, the Table 1. DHA content of the enriched Artemia and of 32 days after
hatching (DAH) larvae (n 3, twenty pooled larvae), and larval dry weight (mg) at 23 and 32 DAH*
(Mean values and standard deviations)
NDHA MDHA HDHA
Mean SD Mean SD Mean SD
DHA content (g DHA/100 g fatty acid)
Artemia ND 3·0 8·1
Larvae 1·7a 0·1 6·7b 0·2 10·4c 0·5
Larval dry weight (mg)
23 DAH (n 60) 1·53a 0·30 1·68a 0·35 1·92b 0·36
32 DAH (n 72) 4·64a 1·05 5·18ab 1·23 5·30 1·32b
HDHA, high DHA; MDHA, medium DHA; ND, not detected by GLC analysis; NDHA, no dietary DHA.
a,b,cMean values within a row with unlike superscript letters were significantly
different (P, 0·05).
* For details of procedures, see Materials and methods.
British
Journal
of
Nutrition
https://www.cambridge.org/core . Universidade do Algarve , on 12 Feb 2019 at 13:38:40, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms
.
results showed that when larvae were fed a diet void of DHA a significantly higher amount of DHA was incorporated into the gut and was concomitantly less in the remaining dissected body. This was certainly related to the state of malnutrition of these larvae but it remains unclear whether the results might be explained by a preferential incorporation of dietary DHA into the membranes of the gastrointestinal mucosa or, alternatively, by a kinetic effect of slowed down intracellular transport, under nutritional deficiency conditions.
As speculated, the assumption that an increase in the dietary supply would result in its increased oxidation for energy pur-poses proved to be true. In fact, a rise in the dietary DHA con-tent led to significant differences in larval metabolism, with a significantly higher proportion of the absorbed DHA label being catabolized in larvae fed HDHA, compared to NDHA. For the MDHA treatment the catabolism was intermediate but not significantly different from NDHA or HDHA. The metabolic trial thus supports the growth results at 32 DAH, which appear to indicate that for Senegalese sole larvae fed enriched Artemia, after settlement, the requirements for DHA are probably not higher than 3 g DHA/100 g total FA. When larval growth was analysed at 23 DAH, only 7 d after the start of the dietary experiment, the results might have pointed to a higher DHA requirement (of around 8 g DHA/ 100 g total FA), since significant differences were only noted between HDHA-fed larvae and the remaining treatments. This may mean that DHA requirements of Senegalese sole larvae decrease with age and stage of development, or that the metabolic trial presented here might give an earlier indi-cation of larval requirements, compared to other biological indicators, such as survival, growth, pigmentation and stress resistance. Nonetheless, further experiments are required using a larger number of dietary treatments providing graded levels of DHA, in order to narrow down the Senegalese sole larval requirements for this EFA. If a finer gradient of dietary DHA supply is tested the requirement may be estimated more accurately by the inflection point where the oxidation of the label significantly increases. This breakpoint might be ident-ified using bi-phase linear regression analysis, as is done for
IAA requirement studies applying the indicator amino acid oxidation concept(13).
In conclusion, the present study describes a methodology, based on tube feeding of a radioactive FA tracer and by fol-lowing its metabolic fate, which may be used to estimate larval EFA requirements, in conjunction with dose – response studies. Growth has normally been used as an indicator to estimate requirements, as it normally reflects well dietary deficiencies. However, dose – response studies, which are becoming more common in larval nutritional research, may benefit from including a metabolic study of this type, particu-larly for narrowing down EFA requirements, as growth may not be a sufficiently sensitive parameter to discriminate between dietary treatments having close levels of EFA. In addition, it might be speculated that the metabolic response to a dietary supply of DHA in excess to requirements, such as an increased oxidative utilization for energy production, could be visible earlier than the physiological consequences of an unbalanced diet, although this remains to be shown. On the other hand, by tube feeding different EFA, such as DHA, EPA and arachidonic acid, to larvae fed a single set of diets differing in the relative levels and proportions between the different EFA it might be possible to estimate the requirement for multiple EFA simultaneously, in a single trial. Addition-ally, competitive interactions between dietary EPA and arachi-donic acid and optimal ratios for eicosanoid production might also be addressed using this methodology, possibly even refin-ing it further to include different body compartments of inter-est. Finally, one of the main interests of this technique may well be in studies of the effect of broodstock nutrition on early larval performance. For instance, it might be possible to correlate dietary levels of specific EFA in broodstock diets and the egg and early larvae FA composition with larval metabolism, before any effects on larval performance and quality become visible. However, most of these suggested potential applications of the methodology described here remain untested and should be explored in the next few years, hopefully helping to advance knowledge in the field of marine fish larvae nutrition.
Fig. 1. Label in each compartment (% total tube fed), 24 h after tube feeding 23 nl of a fish oil mixture containing [1-14C]DHA to 28 days after hatching Senegalese
sole larvae that were previously fed enriched Artemia containing graded concentrations of DHA (DHA/100 g fatty acid: A, 0 g; , 3 g; , 8 g). Values are means with their standard deviations depicted by vertical bars (n 12).a,bMean values within each compartment with unlike letters were significantly different (P, 0·05).
British
Journal
of
Nutrition
https://www.cambridge.org/core . Universidade do Algarve , on 12 Feb 2019 at 13:38:40, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms
.
Acknowledgements
S. M. was supported by ‘Fundac¸a˜o para a Cieˆncia e a Tecno-logia’, Portugal (SFRH/BD/4902/2001). The experiments were supported by the Project PROMAR/SP5.P117/03 (pro-gramme INTERREG IIIA, co-funded by FEDER, European Commission), and comply with the current laws in Portugal, the country in which the present work was conducted. The authors are very grateful to Dr Marisol Izquierdo and Carmen Quintana, from Grupo de Investigacio´n en Acuicultura, ULPGC and ICCM, Las Palmas de Gran Canaria, Spain, for performing the fatty acid analysis. The authors would also like to acknowledge the IPIMAR Aquaculture Research Centre (Olha˜o, Portugal) for the supply of Senegalese sole larvae. The authors declare no conflicts of interest. S. M. was responsible for designing and conducting the experiment, as well as writing the manuscript, while L. E. C. C. collaborated in the experimental design and manuscript preparation.
References
1. Sargent J, Bell G, McEvoy L, et al. (1999) Recent developments
in the essential fatty acid nutrition of fish. Aquaculture 177, 191 – 199.
2. Sargent J, McEvoy L, Estevez A, et al. (1999) Lipid nutrition of
marine fish during early development: current status and future directions. Aquaculture 179, 217 – 229.
3. Tocher DR & Harvie DG (1988) Fatty acid compositions of the
major phosphoglycerides from fish neural tissues; (n-3) and (n-6) polyunsaturated fatty acids in rainbow trout (Salmo gaird-neri) and cod (Gadus morhua) brains and retinas. Fish Physiol Biochem 5, 229 – 239.
4. Bell MV, Batty RS, Dick JR, et al. (1995) Dietary deficiency of
docosahexaenoic acid impairs vision at low light intensities in juvenile herring (Clupea harengus L.). Lipids 30, 443 – 449.
5. Este´vez A & Kanazawa A (1996) Fatty acid composition of
neural tissues of normally pigmented and unpigmented juvenile of Japanese flounder using rotifer and Artemia enriched in n-3 HUFA. Fish Sci 62, 88 – 93.
6. Watanabe T (1993) Importance of docosahexaenoic acid in
marine larval fish. J World Aquac Soc 24, 152 – 161.
7. Furuita H, Takeuchi T & Uematsu K (1998) Effects of
eicosa-pentaenoic and docosahexaenoic acids on growth, survival and brain development of larval Japanese flounder (Paralichthys olivaceus). Aquaculture 161, 269 – 279.
8. Bransden MP, Cobcroft JM, Battaglene SC, et al. (2005) Dietary
22:6n-3 alters gut and liver structure and behaviour in larval striped trumpeter (Latris lineata). Aquaculture 248, 275– 285.
9. Conceic¸a˜o LEC, Dersjant-Li Y & Verreth JAJ (1998) Cost of
growth in larval and juvenile African catfish (Clarias gariepi-nus) in relation to growth rate, food intake and oxygen consumption. Aquaculture 161, 95 – 106.
10. Kolkovski S (2005) Use of ‘tailor-made’ live-feed enrichments
and additives in fish larvae nutritional research. In Larvi 2005,
4th Fish & Shellfish Larviculture Symposium. European Aqua-culture Society Special Publication no. 36, pp. 242 – 245 [CI Hendry, G Van Stappen, M Wille and P Sorgeloos, editors]. Oostende, Belgium: European Aquaculture Society.
11. Villalta M, Este´vez A, Bransden MP, et al. (2005) The effect of
graded concentrations of dietary DHA on growth, survival and tissue fatty acid profile of Senegal sole (Solea senegalensis) larvae during the Artemia feeding period. Aquaculture 249, 353 – 365.
12. Villalta M, Este´vez A, Bransden MP, et al. (2008) Effects of
dietary eicosapentaenoic acid on growth, survival, pigmentation and fatty acid composition in Senegal sole (Solea senegalensis) larvae during the Artemia feeding period. Aquacult Nutr 14, 232 – 241.
13. Elango R, Ball RO & Pencharz PB (2008) Indicator amino
acid oxidation: concept and application. J Nutr 138, 243 – 246.
14. Folch J, Lees M & Sloane Stanley GH (1957) A simple method
for the isolation and purification of total lipides from animal tissues. J Biol Chem 226, 497 – 509.
15. Fox C (1990) Studies on polyunsaturated fatty acid nutrition in
larvae of marine fish – the herring, Clupea harengus L. PhD Thesis, University of Stirling.
16. Izquierdo MS, Watanabe T, Takeuchi T, et al. (1990)
Optimum EFA levels in Artemia to meet the EFA require-ments of red sea bream (Pagrus major). In Current Status
of Fish Nutrition in Aquaculture, pp. 221 – 232 [M
Takeda and T Watanabe, editors]. Tokyo: Tokyo University Fisheries.
17. Rønnestad I, Rojas-Garcı´a CR, Tonheim SK, et al. (2001)
In vivo studies of digestion and nutrient assimilation in marine fish larvae. Aquaculture 201, 161 – 175.
18. Morais S, Koven W, Rønnestad I, et al. (2005) Dietary protein:
lipid ratio and lipid nature affects fatty acid absorption and metabolism in a teleost larva. Br J Nutr 93, 813 – 820.
19. Zar JH (1996) Biostatistical Analysis. Upper Saddle River, NJ:
Prentice Hall International.
20. Morais S, Narciso L, Dores E, et al. (2004) Lipid enrichment
for Senegalese sole (Solea senegalensis) larvae: effect on larval growth, survival and fatty acid profile. Aquacult Int 12, 281 – 298.
21. Johnsen RI, Grahl-Nielsen O & Roem A (2000) Relative
absorption of fatty acids by Atlantic salmon Salmo salar from different diets, as evaluated by multivariate statistics. Aquacult Nutr 6, 255 – 261.
22. Sire M-F & Vernier J-M (1981) E´ tude ultrastructurale de la
synthe`se de chylomicrons au cours de l’absorption intestinale des lipides chez la Truite. Influence de la nature des acides gras inge´re´s. Biol Cellulaire 40, 47 – 62.
23. Pe´rez JA, Rodrı´guez C & Henderson RJ (1999) The uptake and
esterification of radiolabelled fatty acids by enterocytes isolated from rainbow trout (Oncorhynchus mykiss). Fish Physiol Biochem 20, 125 – 134.
24. Geurden I, Bergot P, Schwarz L, et al. (1998) Relationship
between dietary phospholipid classes and neutral lipid absorp-tion in newly-weaned turbot, Scophthalmus maximus. Fish Physiol Biochem 19, 217 – 228.
British
Journal
of
Nutrition
https://www.cambridge.org/core . Universidade do Algarve , on 12 Feb 2019 at 13:38:40, subject to the Cambridge Core terms of use, available at
https://www.cambridge.org/core/terms
.
![Fig. 1. Label in each compartment (% total tube fed), 24 h after tube feeding 23 nl of a fish oil mixture containing [1- 14 C]DHA to 28 days after hatching Senegalese sole larvae that were previously fed enriched Artemia containing graded concentrations of](https://thumb-eu.123doks.com/thumbv2/123dok_br/18032360.861426/4.892.200.680.105.370/compartment-containing-hatching-senegalese-previously-enriched-containing-concentrations.webp)
