Contents lists available atScienceDirect
Pharmacology, Biochemistry and Behavior
journal homepage:www.elsevier.com/locate/pharmbiochembeh
Neuroprotective e
ff
ects of valproic acid on brain ischemia are related to its
HDAC and GSK3 inhibitions
Monalisa Ribeiro Silva
a,b, Alyne Oliveira Correia
a, Gabriel Cabral Alencar dos Santos
a,
Lucas Leimig Telles Parente
a, Keicy Parente de Siqueira
a, Danielly Gonçalves Sombra Lima
a,
Jonathan Almeida Moura
a, Ana Elisa da Silva Ribeiro
a, Roberta Oliveira Costa
b,
Daniel Luna Lucetti
a,b, Elaine Cristina Pereira Lucetti
a, Kelly Rose Tavares Neves
b,
Glauce Socorro de Barros Viana
a,b,⁎aFaculty of Medicine Estácio of Juazeiro do Norte (Estácio FMJ), Juazeiro do Norte, Brazil bFaculty of Medicine of the Federal University of Ceará (UFC), Fortaleza, Brazil
A R T I C L E I N F O
Keywords: Valproic acid Neuroprotection Inflammation Oxidative stress HDAC GSK3
A B S T R A C T
Valproic acid (VA) is an antiepileptic that is also used for the treatment of bipolar disorders. The objective was to evaluate the neuroprotective effects of VA on a brain ischemia model. The groups of male Wistar rats were: SO (sham-operated), ischemic and ischemic treated with VA (25, 50 and 100 mg/kg, p.o.). After anesthesia with ketamine and xilazine, the animals were subjected to clamping of carotid arteries (30 min) and reperfusion. Except for the carotid clamping, the SO group was submitted to the same procedure. On the 7th day, the animals were behaviorally evaluated, euthanized and had their brain dissected for neurochemical and im-munohistochemical assays. The data were analyzed by ANOVA and Tukey as the post hoc test. The results showed that VA reversed partly or completely the behavioral (locomotor activity and memory deficits), neu-rochemical (striatal DA and DOPAC levels, brain nitrite and lipid peroxidation) and immunohistochemical al-terations (iNOS, COX-2, HDAC and GSK3) observed in the untreated ischemic group. VA neuroprotective effects are probably related to its anti-inflammatory and antioxidant properties, as well as to HDAC and GSK3 inhibitory effects. Thesefindings stimulate translational studies focusing on VA as a neuroprotective drug to be potentially used in the clinic for several neurological conditions.
1. Introduction
Valproic acid (VA) is a short-chain fatty acid that has been widely used, for decades, as an anticonvulsant agent and a mood stabilizer. Although VA mechanism of action is associated with the regulation of GABAergic neurotransmission, it also affects multiple signal transduc-tion pathways that result from inhibitransduc-tions of enzymes, as HDAC and GSK3, among others (Ximenes et al., 2012).
Stroke is a worldwide leading cause of death and disability, where the ischemic stroke is the most common type (Shichita et al., 2012). Furthermore, the pathophysiology of brain ischemia involves an early tissue damage phase, characterized by neuronal death, inflammation and blood-brain barrier breakdown, followed by a late phase associated with neurovascular recovery (Fessler et al., 2013). Thus, brain in-flammation occurs in necrotic tissue, following the breakdown of the blood-brain barrier and infiltration of immune cells, both responsible
for the ischemic brain inflammation by producing inflammatory med-iators (Lakhan et al., 2009;Jin et al., 2010).
Histone deacetylases (HDACs) are involved in the epigenetic (post-translational) regulations of gene expression and protein functions (Ziemka-Naleca and Zalewska, 2014). Epigenetic mechanisms as post-translational histone acetylation have been recognized for their con-tribution to gene activation and silencing in the brain, contributing to neurodegeneration (Wu et al., 2013). Most importantly, in vivo and in vitro data indicate that HDAC inhibitors present anti-inflammatory actions, due to their effects on cell death through acetylation of non-histone proteins (Adcock, 2007;Halili et al., 2009;Shakespear et al., 2011;Grabiec et al., 2011;Cantley and Haynes, 2013). In that sense, HDAC inhibitors (including VA), by inhibiting HDAC and presenting anti-inflammatory activity, appear as potential candidates for clinical trials in stroke patients, as has been already suggested (Kim et al., 2007).
https://doi.org/10.1016/j.pbb.2018.02.001
Received 16 January 2017; Received in revised form 8 February 2018; Accepted 9 February 2018 ⁎Corresponding author at: Rua Barbosa de Freitas, 130/1100, Fortaleza 60170-020, Brazil.
E-mail address:[email protected](G.S. de Barros Viana).
Available online 13 February 2018
0091-3057/ © 2018 Elsevier Inc. All rights reserved.
Glycogen synthase kinase-3 (GSK3) is recognized as a powerful regulator of inflammation, while its inhibition provides protection against inflammatory conditions (Jope et al., 2007). GSK3 is a serine/ threonine kinase known for regulating several cellular functions, as gene expression, among others. While GSK3 leads to the production of inflammatory molecules and cell migration, making this enzyme a powerful regulator of inflammation, its inhibition provides protection from inflammatory conditions (Jope et al., 2007;Yuskaitis and Jope, 2009;Beurel, 2011). In addition, GSK3 inhibitors have been reported to be beneficial in brain ischemic models and could be a novel and pro-mising therapy for ischemic stroke (Valerio et al., 2011,Chuang et al., 2011,Venna et al., 2015).
VA, probably due to its HDAC and GSK3 inhibitory properties, is a potent anti-inflammatory agent, as has been already shown by us and others as well (Zhang et al., 2008;Suda et al., 2013;Ximenes et al., 2013; Zheng et al., 2014;Costalonga et al., 2016). Thus, the objectives of the present study were to evaluate the neuroprotective effects of VA on a global ischemic model in rats, by behavioral (openfield and water maze tests), biochemical (DA and DOPAC measurements and nitrite and TBARS determinations) and, mainly, by immunohistochemical ap-proaches, attempting to clarify its neuroprotective mechanism of action.
2. Material and methods
2.1. Drugs and reagents
Sodium valproate (syrup, 250 mg/5 mL; from the Teuto/Pfizer Laboratory, Anápolis, GO, Brazil). Ketamine and xylazine were pur-chased from König SA, Argentina. 2,3,5-Triphenyltetrazolium chloride (TTC), standard monoamines and standard amino acids were from Sigma-Aldrich, USA. All other reagents were of analytical grade.
2.2. Animals
Male Wistar rats (200–250 g) were obtained from the Animal House of the Faculty of Medicine Estácio of Juazeiro do Norte, Brazil. The animals were housed at 24 ± 2 °C, under a 12-h light/12-h dark cycle, and had free access to a standard pellet diet (Purina chow) and tap water. They were deprived of food for 8 h, before the experiments, except for drinking water. The animals were treated in accordance with the current law and the NIH Guide for the Care and Use of Laboratory Animals. The project was previously approved by the Animal's Ethics
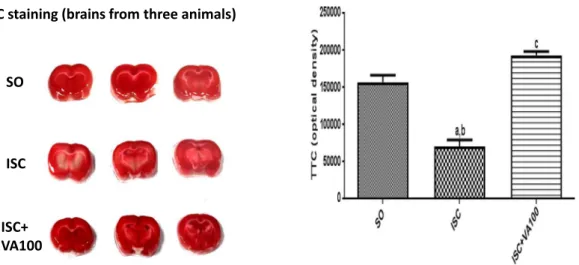
TTC staining (brains from three animals)
SO
ISC
ISC+
VA100
Fig. 1.The valproic acid (VA) treatment attenuates brain ischemic areas, as evaluated by TTC staining. The left panel shows coronal slices photos of three different animals from each of
the SO, ISC and ISC + VA100 groups. The graph shows the means ± SEM from the photos in the left and was produced by the Image J software (NIH, USA). a. vs. SO, q = 10.71, p < 0.001; b. vs. ISC + VA100, q = 15.25, p < 0.001; c. vs. SO, q = 4.536, p < 0.05 (One way ANOVA and Tukey as the post hoc test).
SO
ISC
ISC+VA
25
ISC
+VA
50
ISC
+VA
100
0
5
10
15
20
a,b,c
ni
m
5/
s
g
ni
s
s
or
c
f
o
.
o
N
Fig. 2.The valproic acid (VA) treatment reverses behavioral changes (locomotor activity,
measured as the number of crossings/5 min), observed in the untreated ischemic-group, as evaluated by the openfield test. The values are means ± SEM from groups of 11 to 24 animals. a. vs. SO, q = 7.591, p < 0.001; b. vs. ISC + VA50, q = 4.860, p < 0.01; c. vs. ISC + VA100, q = 7.134, p < 0.001 (One way ANOVA and Tukey as the post hoc test).
SO
ISC
ISC+VA
25
ISC+VA
50
IS
C+VA
10
0
0
10
20
30
40
50
a,b,c,d
)
s(
mr
of
t
al
p
e
ht
d
ni
f
ot
e
mi
T
Fig. 3.The valproic acid (VA) treatment reverses the memory impairment, observed in
Committee of the Faculty of Medicine of the Federal University of Ceará, Brazil (No. 59/2011).
2.3. Global ischemia protocol
The animals were anesthetized with ketamine (75 mg/kg, i.p.) and xylazine (10 mg/kg, i.p.) and submitted to the model of transitory global ischemia, by the bilateral occlusion of both carotids, for 30 min. After reperfusion, the incision was sutured and the animals were placed in individual plastic cages for recovery, with water and food ad libitum. The sham-operated group (SO, control) was submitted to the same procedure, except for the clamping of the carotids. After surgery, the animals were orally treated (by gavage) with VA, at the doses of 25, 50 and 100/kg, daily for 7 days. The SO and untreated ischemic groups received distilled water (1 mL/kg), under the same conditions as the VA treated groups. The animals were distributed into 5 groups, as follows: SO, ischemic untreated and ischemic groups after treatments with VA (25, 50 and 100 mg/kg, p.o.). On the 7th day of treatment and 1 h after the last drug administration, the animals were submitted to behavioral
tests (openfield and water maze tests) and euthanized forstriata dis-sections and hippocampus slicings. The striatawere used for homo-genate preparations and DA and DOPAC determinations. The VA anti-oxidant activity was evaluated by nitrite and lipid peroxidation measurements in the cortex, striatum and hippocampus areas. Hippocampus slices were also processed for immunohistochemistry assays.
2.4. Determination of brain ischemic areas by TTC staining
TTC staining is considered a reliable method for detection of cere-bral infarction, in rats after ischemia (Bederson et al., 1986). It mea-sures tissue viability and thus infarct size. Although the formation of formazan from TTC depends on both the staining method and the me-tabolic burden of the brain tissue (Benedek et al., 2006), it can be used for defining the size of cerebral infarction and penumbra (Kramer et al., 2010). In the present study, the animals were decapitated and their brains removed for TTC (2,3,5-triphenyl tetrazolium chloride) staining. Then, 2 mm coronal sections were obtained with the rodent brain
SO ISC ISC+VA 25 ISC+V A50 ISC +VA 100 0 1000 2000 3000 4000 5000
a,b
) e u s si t g/ g n( A D SO ISC ISC+V A25 ISC+VA 50 ISC+VA 100 0 1000 2000 3000a,b,c
d
) e u s si t g/ g n( C A P O DFig. 4.The valproic acid (VA) treatment reverses
dose-de-pendently the decreases in DA and DOPAC striatal contents, demonstrated in the untreated ischemic-animals. The va-lues are means ± SEM from groups of 5 to 11 animals). DA: a. vs. SO, q = 4.491, p < 0.05; b. vs. ISC + VA100, q = 4.128, p < 0.05. DOPAC: a. vs. SO, q = 4.189, p < 0.05; b. vs. ISC + VA50, q = 4.260, p < 0.05; c. vs. ISC + VA100, q = 7.496, p < 0.001; d. vs. ISC + VA100, q = 5.847, p < 0.01 (One way ANOVA and Tukey as the post hoc test).
SO ISC ISC+VA 25 ISC+VA50 ISC+VA 100 0 5 10 15 20
a,b,c
Cortex ( eti rti N) e u s si t g/l o mSO ISC
ISC+VA 25 ISCV A50 ISC+VA100 0 5 10 15 20 25
a,b,c,d
Hippocampus ( eti rti N) e u s si t g/ l o mSO ISC
ISC+VA 25
ISC+VA50 ISC+V A100 0 5 10 15 20
a,b,c,d
e,f,g
Striatum ( eti rti N) e u s si t g/ l o m µ µ µFig. 5.The valproic acid (VA) treatment partly reversed the
matrix apparatus (Harvard, USA). The slices were incubated with TTC, for 30 min at 37 °C in the dark, andfixed by immersion in 4% buffered formaldehyde solution (pH 7.4), for 24 h. Afterwards, the sections were photographed and ischemic areas were quantified by the Image J software (NIH, USA). In those cell populations, where the electron mitochondrial transport is maintained, the TTC shows a red color that decreased in brain ischemic areas.
2.5. Behavioral testing
2.5.1. Openfield test
This test is used for measuring locomotor activity in rodents and can also serve to determine motor deficits and anxiety. It was originally described for the study of emotionality in rats and, now, is one of the most popular models in animal psychology (Prut and Belzung, 2003). C Locomotor activity is measured by determining the amount of distance traveled and stereotyped behaviors, as rearing and grooming. The test is sensitive to motor dysfunction, as well as hippocampus and basal
ganglia damage. For that, we used an arena (50 × 50 × 20 cm) divided into four equal quadrants. Each animal was evaluated for 5 min, at the 7th day after ischemia and the number of quadrants crossed with all 4 paws by the animal was determined.
2.5.2. Morris water-maze test
This is a test of spatial learning for rodents that relies on distal cues to navigate, from start locations, around the perimeter of an open swimming arena, in order to locate a submerged escape platform. The original procedure was described as Morris water maze (Morris, 1981; Vorhees and Williams, 2006). Spatial learning is assessed across re-peated trials and reference memory. The test has proven to be a robust and reliable one and is strongly correlated with hippocampus synaptic plasticity and NMDA receptor function. For that, we used a circular black pool (1.7 m of diameter and 1 m height),filled with water (0.59 m deep), at a 25 °C temperature. The pool is divided into four quadrants and has a10 cm diameter platform, immersed 0.5 cm below the water surface. The room is provided with four cues located on North, South,
SO ISC ISC+VA 25 ISC +VA5 0 ISC +VA 100
0
500
1000
1500
a,b,c,d
Cortex
( S R A B T) e u s si t g/ A D M l o m SO ISC ISC+V A2 5 ISC+VA 50 ISC+VA 1000
500
1000
1500
a,b,c,d
Hippocampus
( S R A B T) e u s si t g/ A D M l o mSO ISC
ISC+VA 25 ISC+VA 50 ISC+ VA 100
0
500
1000
1500
a,b,c,d
Striatum
( S R A B T) e u s si t g/ A D M l o m µ µ µ µFig. 6.The valproic acid treatment partly reversed the lipid peroxidation, seen in the untreated ischemic-rats. The values are means ± SEM from groups of 4 to 7 animals. Cortex: a. vs.
East and West of the walls. The animals were subjected to two trials, for two consecutive days (pre-training) and, 48 h after the last trial, to the test in the water maze. The animal had a maximum time of 54 s (cutoff time) for finding the platform, where it would stay for 15 s, having however a second chance in case of failure.
2.6. DA and DOPAC determinations by HPLC
For measurements of dopamine (DA) and its metabolite (3,4 dihy-droxyphenylacetic acid, DOPAC),striatafrom all groups were used for preparing 10% homogenates. Homogenates were sonicated in 0.1 M HClO4, for 30 s, centrifuged at 4 °C for 15 min at 15,000 rpm, and the supernatants werefiltered (0.2μm, Millipore). Twenty-microliter sam-ples were then injected into a high-performance liquid chromatography (HPLC) column. The mobile phase was 0.163 M citric acid, pH 3.0, containing 0.02 mM EDTA with 0.69 mM sodium octanesulfonic acid (SOS), as an ion pairing reagent, 4% v/v acetonitrile and 1.7% v/v tetrahydrofuran. The monoamines were electrochemically detected, using an amperometric detector (Shimadzu, Japan), by oxidation on a glassy carbon electrode at 0.85 V relatively to the Ag–AgCl reference electrode. Their concentrations were determined by comparison with standards injected into the HPLC column on the day of experiment, and the values expressed as ng/g tissue.
2.7. Determination of nitrite contents
The anion nitrite (NO2−) is considered a biochemical reservoir for nitric oxide (NO), and nitrite reduction to NO occurs at increasing rates in brain ischemia (Dezfulian et al., 2007). Furthermore, evidences
indicate that the monitoring of plasma nitrite can be used as a marker of ischemic stroke (Silver and Lapchak). In this assay, the Griess reagent (1 part 0.1% naphthylethylenediamine dihydrochloride in distilled water plus 1 part 1% sulfanilamide in 5% H3PO4) shows the presence of ni-trites in the sample. Brain homogenates from hippocampus, striatum and cortex areas (10% in KCl buffer) were centrifuged (12,000 rpm for 10 min) and 100μL supernatants were added to 100μL Griess reagent, and the mixture stayed on RT for 10 min. The standard NaNO2curve was obtained (in spectrophotometer, at 520 nm) and used for calcu-lating the results expressed asμmol nitrite per g tissue (Green et al., 1982).
2.8. Determination of lipid peroxidation (TBARS assay)
Lipid peroxidation expresses oxidative stress induced by ROS re-activity. A largely used method for measuring it is the determination of malondihaldehyde (MDA) in biological samples (Draper and Hadley, 1990). Previous data (Bromont et al., 1989) showed a delayed and long-lasting increase in lipid peroxidation that occurs in ischemia-sensitive brain regions, such as hippocampus, striatum and cortex that parallels the development of neuronal necrosis, supporting the hypothesis that free radical processes participate in post-ischemia neuronal damage. In the present work, 10% homogenates from hippocampus, striatum and cortex prepared in 1.15% KCl were added (250μL) to 1 mL 10% TCA, followed by the addition of 1 mL 0.6% thiobarbituric acid. After agi-tation, this mixture was maintained in a water-bath (95–100 °C), for 15 min. Then, the mixture was cooled on ice and centrifuged (4000 rpm/5 min). The TBARS content was determined in a plate reader, at 540 nm, with results expressed inμmol MDA per g tissue. A
SO
ISC
ISC+VA100
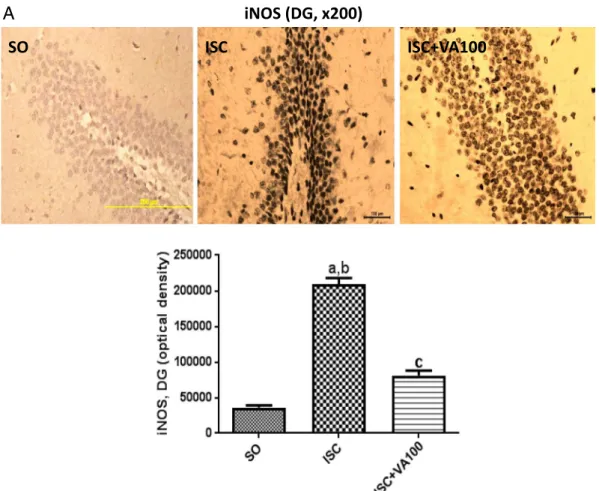
iNOS (DG, x200)
A
Fig. 7.The valproic acid (VA) treatment partly or completely reversed the iNOS immunoreactivity, in the dentate gyrus (DG) and temporal cortex (TC), respectively, seen in the untreated
standard curve with several MDA concentrations was also performed.
2.9. Immunohistochemistry assays for iNOS, COX-2, HDAC and GSK3
Brain hippocampus sections (from the temporal cortex, TC and the dentate gyrus, DG) werefixed in 10% buffered formol, for 24 h, followed by a 70% alcohol solution, and embedded into paraffin wax for slices processing on appropriate glass slides. These were placed into the oven at 58 °C, for 10 min, followed by deparaffinization in xylol, rehydration in alcohol at decreasing concentrations, washing in distilled water and PBS (0.1 M sodium phosphate buffer, pH 7.2), for 10 min. The endogenous peroxidase was blocked with a 3% hydrogen peroxide solution, followed by incubation with the appropriate primary anti-antibody for iNOS and COX-2, HDAC and GSK3 and diluted according to the manufacturers' instructions (Santa Cruz or Millipore, USA), for 2 h at room temperature, in a moist chamber. The glass slides were then washed with PBS (3 times, 5 min each) and incubated with the biotinylated secondary antibody, for 1 h at room temperature, in a moist chamber. Then, they were washed again in PBS and incubated with streptavidin-peroxidase, for 30 min, at room temperature (in a moist chamber) and, after afinal wash in PBS, incubated in 0.1% DAB solution (in 3% hydrogen peroxide). Finally, the glass slides were washed in distilled water and counterstained with Mayers hematoxylin, washed in tap water, dehydrated in alcohol (at increasing concentrations), diaphanized in xylol and mounted on Entelan®for optic microscopy examination. The data were quantified by the Image J software (NIH, USA).
2.10. Statistical analyses
The data are presented as means ± SEM and analyzed by One-way ANOVA, followed by Tukey as thepost-hoctest. Whenever needed, the data were analyzed by two-tailed unpaired Student's t-test. The
differences were considered statistically significant at p < 0.05. The immunohistochemistry data were analyzed by the Image J software (NIH, USA). All the statistical analyses were made with the GraphPad Prism, version 6.0.
3. Results
3.1. Measurement of brain ischemic areas by TTC staining
In order to evaluate the degree of infarcted brain areas, 3 animals from each group were used to perform the TTC staining. This method permits a fast and reliable visualization of the hypoxic brain tissue. Our data showed an average 55% decrease in the TTC staining after brain ischemia, in relation to the SO group, indicating tissue damage and metabolic impairment. This alteration was completely reversed in the ischemic group, after VA treatments with its higher dose (Fig. 1).
3.2. Openfield test
Our results (Fig. 2) from groups of 11 to 24 animals showed that the brain ischemia decreased by 51% the animal's locomotor activity, re-latively to the control group. On the other hand, treatments of the is-chemic groups with VA returned this parameter towards normality, and values similar to those of the SO group were observed after VA at the two higher doses (50 and 100 mg/kg).
3.3. Water maze test
This test was carried out in groups of 13 to 30 animals. The un-treated ischemic group showed almost a 2-times increase in the time to find the hidden platform, relatively to controls, indicating memory impairment. No significant differences were seen in the ischemic
iNOS (TC, x200)
SO
ISC
ISC+VA100
B
groups, after VA treatments with all three doses, in relation to the SO group, suggesting a significant improvement of hippocampus function and spatial memory (Fig. 3).
3.4. DA and DOPAC measurements
Groups of 5 to 11 animals were used. We demonstrated a 55% de-crease in DA contents, in the striata from untreated ischemic animals, compared to the SO group (control). The VA treatments of ischemic groups brought these values towards normality, in a dose dependent manner. As a matter of fact, DA levels in the ISC + VA100 group were even higher than those of controls. Similar results were seen in DOPAC levels, with values 74% lower than those of controls, in the untreated ischemic group, and returning to values close to normal ones after VA treatments with the two higher doses. Interestingly, the ischemic group after treatments with VA, at the dose of 100 mg/kg, showed DOPAC values 1.6-times higher than those of controls (Fig. 4).
3.5. Brain nitrite measurements
Groups of 3 to 7 animals were used. We showed > 2-times increases in nitrite contents in the ischemic groups, in relation to the control group, not only in the cortex but also in the hippocampus and striatum. Although nitrite levels were lower in the ischemic groups after VA treatments, the values were still somewhat higher in all three areas, than those of controls (Fig. 5).
3.6. Brain lipid peroxidation measurements
Groups of 4 to 7 animals were used. Increases ranging from 2.6- to 3.5-times were observed in the cortex, hippocampus and striatum in the ischemic groups, compared to the control group. Although higher levels of lipid peroxidation were demonstrated in the cortex, the values in the three brain areas were very close to those of the control group, in is-chemic groups after VA treatments (Fig. 6).
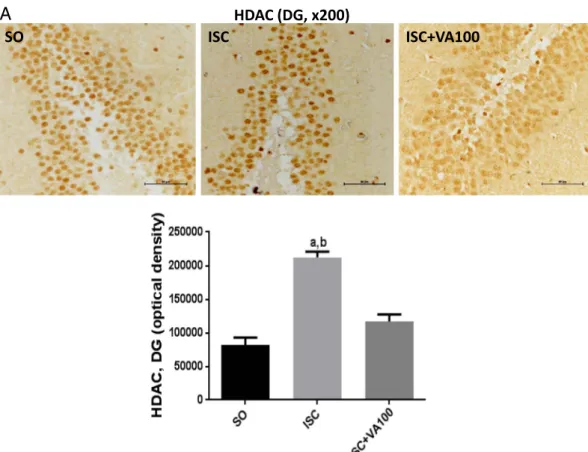
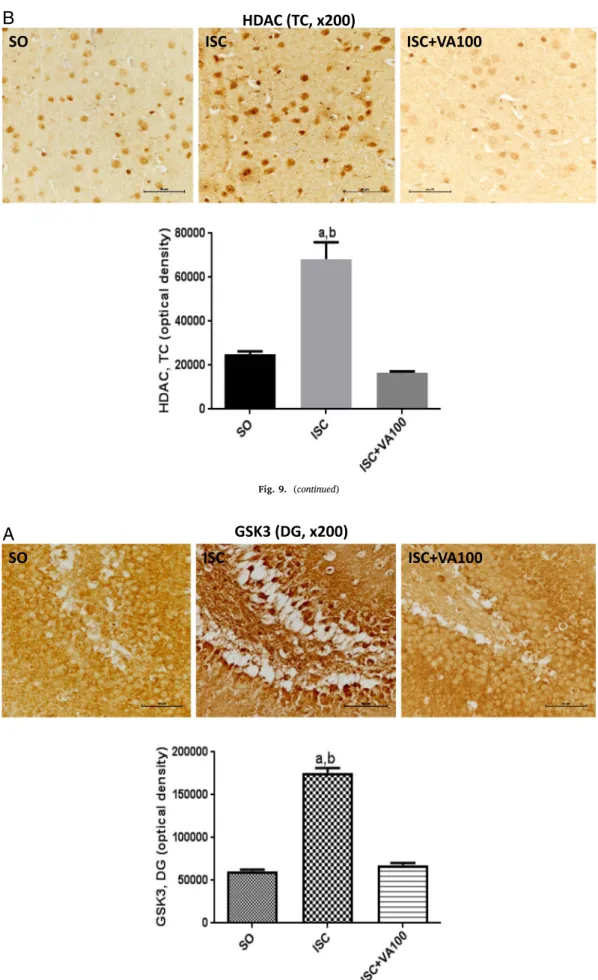
3.7. Immunohistochemistry analyses
The immunohistochemistry analyses were performed in the dentate gyrus (DG) and temporal cortices (TC) from three animals per group. The untreated ischemic animals presented 5.8- and 3.6-times increases of the iNOS immunoreactivity in the DG and TC, respectively, as related to the SO group. Although the ISC + VA100 group showed significant higher values (2.2-times) in the DG area, relatively to the SO group, no significant difference was detected between the SO and ISC + VA100 groups in the TC area (Fig. 7). In the case of COX-2 immunoreactivities, 2.2- and 2.1-times increases were respectively seen in DG and TC areas, from the untreated ischemic group, compared with the SO group. No significant differences were noticed between the SO and ISC + VA100 groups from both DG and TC areas (Fig. 8). Similar results were de-monstrated for HDAC and GSK3 immunoreactivities in these two brain areas. Thus, 2.6- and 2.7-times increases were respectively observed for HDAC immunoreactivities, in DG and TC areas from the untreated is-chemic animals, compared with the SO group. In these areas, the values were closer or even lower, in the ISC + VA100 groups relatively to the SO group (Fig. 9). In the case of GSK3, an almost 3-times increase was
SO
ISC
ISC+VA100
COX-2 (DG, x200)
A
Fig. 8.The valproic acid (VA) treatment partly reversed the COX-2 immunoreactivity, in the dentate gyrus (DG) and temporal cortex (TC), seen in untreated ischemic-rats. The values are
seen in DG from the untreated ischemic group, compared with the SO group. In addition, a 6.6-times increase was observed in the TC area from the untreated ischemic group, in relation with the SO group. The values were closer (DG) or even lower (TC) than those of the SO groups, after VA treatments (Fig. 10).
4. Discussion
Stroke is a leading cause of death and disability worldwide. The pathophysiology of stroke involves complex mechanisms, resulting from a cascade of events leading to energy depletion and cell death (Kanekar et al., 2012). Furthermore, mitochondria play a key role in tissue injury, since in stroke these organelles lead to energy depletion and to apoptotic and necrotic cell death (Sims and Muyderman, 2010). During and after stroke, the loss of blood-brain barrier (BBB) integrity and edema formation are prominent events contributing to the stroke evolution (Shi et al., 2016).
Angiogenesis, or vascular reorganization, is known to play a role in recovering and remodeling processes after stroke (Prakash and Carmichael, 2015). VA is an HDAC inhibitor presenting neuroprotective actions in several neurological conditions, including brain ischemia (Ziemka-Naleca and Zalewska, 2014). Besides, chronic VA treatment was shown to enhance angiogenesis and to promote functional recovery after brain ischemia, effects probably involving the HDAC inhibition (Wang et al., 2012).
Recentfindings showed that the use of antiepileptic drugs acting on hepatic enzymes as inducers, such as carbamazepine and phenytoin is not associated with an increased risk of ischemic stroke. VA is the only antiepileptic drug inhibiting hepatic enzymes; and its use instead is associated with a decreased risk of ischemic stroke (Renoux et al., 2015). In addition, brain ischemia is always associated with edema, and
VA, due to its HDAC inhibitory properties, is a potent anti-in-flammatory drug as already shown (Zhang et al., 2012;Ximenes et al., 2013).
We have previously demonstrated that VA, at lower doses, de-creased the TNF-alpha immunostaining in the paw-edema induced by carrageenan and this effect was potentiated after its association with SAHA, also a HDAC inhibitor. Another important finding was the neuroprotective effect of VA as observed by us in a PD model in rats, which is probably related to the drug anti-inflammatory and HDAC inhibitory properties (Ximenes et al., 2013;Ximenes et al., 2015).
In the present work, we showed that VA partially and completely reversed behavioral alterations, as seen in the ischemic group after VA treatments, in the openfield and water maze tests for locomotor ac-tivity and spatial memory evaluations, respectively. Interestingly, VA was able to block the memory impairment observed in the ischemic group, suggesting recovery of the hippocampus function. These effects were already observed with the dose of 50 mg/kg.
Studies on the memory effects of VA are contradictory, some of them showing improvements (Bredy and Barad, 2008;Yao et al., 2014) and some showing impairments or no effects (Umka et al., 2010;Sintoni et al., 2013;Pinheiro et al., 2015). A recent study (Jellet et al., 2014) raises the argument that standard VA doses do not cause a cognitive impact, in a model of epilepsy. In human's epilepsy, the daily dosage can be up to 60 mg/kg and the great majority of experimental works use VA doses higher than 100 mg/kg and on a chronic basis. Furthermore, under the conditions of the present work, the beneficial VA effects on memory result from lower VA doses.
We showed that VA completely reversed DA and DOPAC contents in striata from ischemic rats. These effects probably reflect upon the partial recovery of locomotor activity, as seen in the openfield test. Our results are consistent with earlier data (Vriend and Alexiuk, 1996)
SO
ISC
COX-2 (TC, x200)
ISC+VA100
B
showing that valproate influences the metabolism of monoamines, substantially increasing DA metabolites in mice striata. However, these authors used doses of 180 and 360 mg/kg. According to them, these effects could contribute to the VA anticonvulsive effect. Recently (Ximenes et al., 2015), we demonstrated that VA partly reversed the decreased striatal DA and DOPAC concentrations, in a model of PD.
The oxidative stress is known to play an important role in brain ischemia and reperfusion (Love, 1999;Saito et al., 2005;Chen et al., 2011), contributing to increased levels of nitrite and lipid peroxidation. We showed that VA, at the doses of 25, 50 and 100 mg/kg, decreased both brain nitrite contents and lipid peroxidation, in ischemic rats, and these effects correlated to the decrease of infarcted areas, as compared to the untreated ischemic rats. VA use in the clinic is associated with idiosyncratic hepatotoxicity, which can be lethal, and this effect seems to be related to VA-induced oxidative stress (Chang and Abbott, 2006). In addition, VA hepatotoxicity has been proposed to be metabolite-in-duced and dose-dependent, observed with doses up to 500 mg/kg (Lee et al., 2009;Jung et al., 2010) and, thus at a higher dose range.
In the present study, VA treatments partly or completely blocked the increased immunoreactivity for iNOS and COX-2 as seen in brain areas from untreated brain ischemic rats. Previously, we found that VA de-creased TNF-alpha immunostaining in paws from carrageenan-induced edema and MPO activity from human neutrophils, indicating a potent anti-inflammatory action (Ximenes et al., 2013). VA has been shown to decrease the oxidative stress and iNOS expression in a model of spinal cord injury (Lee et al., 2014). Previously (Bosetti et al., 2003), VA was also shown to reduce the arachidonic acid turnover in rat brain and to suppress LPS-induced COX-2 expression in murine brain (Chuang et al., 2014). All these studies corroborate with ourfindings suggesting that the VA neuroprotective effects are related to its antioxidant and anti-inflammatory actions.
Similarly, both HDAC and GSK3 have an important role in in-flammation and brain function (Kim et al., 2007;Ziemka-Naleca and Zalewska, 2014;Volmar and Wahlestedt, 2014;Kim et al., 2007;Jope et al., 2007; Yuskaitis and Jope, 2009;Beurel, 2011). These enzyme inhibitions by VA were shown to reduce ischemic cerebral damage (Valerio et al., 2011; Pang et al., 2016), by restoring mitochondrial bioenergetic failure and preventing ROS production. We showed that VA treatments significantly attenuated the increased HDAC and GSK3 immunoreactivities, demonstrated in brain areas from untreated is-chemic animals. Interestingly, VA and lithium are two widely used drugs for the treatment of bipolar disorders (Jin et al., 2014) and may act on common therapeutic targets, as HDAC and GSK3 (Wu et al., 2013;Stertz et al., 2014).
Although the action mechanism of lithium, a drug used for bipolar disorders as VA, is largely due to its GSK3 inhibition (Harwood and Agam, 2003; Rowe et al., 2007), this question was not completely solved for VA. However, earlier in vitro studies (Chen et al., 1999) suggested that VA inhibits GSK3, a hypothesis which was supported later (Hall et al., 2002). These authors point out that the two mood stabilizers, lithium and valproate, inhibit GSK3, although through dif-ferent pathways. Recentfindings (Zhang et al., 2017) indicate that VA neuroprotective effect in the MPP+ model of PD is associated with GSK3-beta phosphorylation via Akt and ERK activation in the mi-tochondrial intrinsic apoptotic pathway. Others (Qing et al., 2008) found that VA decreased Aβproduction by inhibiting GSK-3β-mediated ϒ-secretase cleavage of APP in vitro and in vivo.
As far as we known, GSK3 inhibition in the ischemic rat brain by VA was demonstrated here, for thefirst time, supporting the view that the potentiation of the VA effect by lithium, as previously shown (Leng et al., 2008), is probably due to GSK3 inhibitions, a common ther-apeutic target for both drugs, as raised above. Furthermore, preclinical
SO
ISC
ISC+VA100
HDAC (DG, x200)
A
Fig. 9.The valproic acid (VA) treatment partly or completely reversed the HDAC immunoreactivity in the dentate gyrus (DG) and temporal cortex (TC), respectively, seen in the
GSK3 (DG, x200)
SO
ISC
ISC+VA100
A
Fig. 10.The valproic acid (VA) treatment partly or completely reversed the GSK3 immunoreactivity in the dentate gyrus (DG) and temporal cortex (TC), respectively, seen in the
untreated ischemic-rats. The values are means ± SEM from groups of 3 animals. DG: a. vs. SO, q = 32.01, p < 0.001; b. vs. ISC + VA100, q = 30.01, p < 0.001. TC: a. vs. SO, q = 42.74, p < 0.001; b. vs. ISC + VA100, q = 50.40, p < 0.001; c. vs. SO, q = 7.660, p < 0.01 (One way ANOVA and Tukey as the post hoc test).
SO
ISC
ISC+VA100
HDAC (TC, x200)
B
studies have shown the potential benefit of VA on brain injury (Rem et al., 2004;Sinn et al., 2007;Chen et al., 2014). The presentfindings should stimulate translational studies focusing on the clinical use of VA, alone or co-administered with lithium, for neurological conditions be-yond bipolar disorders.
Acknowledgments
The authors are grateful to the Brazilian National Research Council (CNPq) and the Foundation for Scientific and Technological Development of the Ceará State, Brazil, for thefinancial support, also to Prof. M.O.L. Viana for the orthographic revision of the manuscript.
Conflict of interest
The authors declare no conflict of interest.
References
Adcock, I.M., 2007. HDAC inhibitors as anti-inflammatory agents. British J. Pharmacol. 150, 829–831.
Bederson, J.B., Pitts, L.H., Germano, S.M., Nishimura, M.C., Davis, R.L., Bartkowski, H.M., 1986. Evaluation of 2,3,5-triphenyltetrazolium chloride as a stain for detection and quantification of experimental cerebral infarction in rats. Stroke 17, 1304–1308.
Benedek, A., Móricz, K., Jurányi, Z., Gigler, G., Lévay, G., Hársing Jr., L.G., Mátyus, P., Szénási, G., Albert, M., 2006. Use of TTC staining for the evaluation of tissue injury in the early phases of reperfusion after focal cerebral ischemia in rats. Brain Res. 116, 159–165.
Beurel, E., 2011. Regulation by glycogen synthase kinase-3 of inflammation and Tcells in CNS diseases. Front. Mol. Neurosci. 4, 18.http://dx.doi.org/10.3389/fnmol.2011. 00018.(Aug 31).
Bosetti, F., Weerasinghe, G.R., Rosenberger, T.A., Rapoport, S.I., 2003. Valproic acid downregulates the conversion of arachidonic acid to eicosanoids via cyclooxygenase-1 and -2 in rat brain. J. Neurochem. 85, 690–696.
Bredy, T., Barad, M., 2008. The histone deacetylase inhibitor valproic acid enhances acquisition, extinction, and reconsolidation of conditioned fear. Learn. Mem. 15, 39–45.
Bromont, C., Marie, C., Bralet, J., 1989. Increased lipid peroxidation in vulnerable brain
regions after transient forebrain ischemia in rats. Stroke 20, 918–924.
Cantley, M.D., Haynes, D.R., 2013. Epigenetic regulation of inflammation: progression from broad acting histone deacetylase (HDAC) inhibitors to targeting specific HDACs. Inflammopharmacology 21, 301–307.
Chang, T.K., Abbott, F.S., 2006. Oxidative stress as a mechanism of valproic acid-asso-ciated hepatotoxicity. Drug Metab. Rev. 38, 627–639.
Chen, G., Huang, L.D., Jiang, Y.M., Manji, H.K., 1999. The mood-stabilizing agent valproate inhibits the activity of glycogen synthase kinase-3. J. Neurochem. 72, 1327–1330.
Chen, H., Yoshioka, H., Kim, G.S., Jung, J.E., Okami, N., Sakata, H., Maier, C.M., Narasimhan, P., Goeders, C.E., Chan, P.H., 2011. Oxidative stress in ischemic brain damage: mechanisms of cell death and potential molecular targets for neuroprotec-tion. Antioxid. Redox Signal. 14, 1505–1517.
Chen, S., Wu, H., Klebe, D., Hong, Y., Zhang, J., 2014. Valproic acid: a new candidate of therapeutic application for the acute central nervous system injuries. Neurochem. Res. 39, 1621–1633.
Chuang, D.-M., Wang, Z., Chiu, C.-T., 2011. GSK3 as a target for lithium-induced neu-roprotection against excitotoxicity in neuronal cultures and animal models of is-chemic stroke. Front. Mol. Neurosci. 4, 15.http://dx.doi.org/10.3389/fnmol.2011. 00015.
Chuang, Y.F., Yang, H.Y., Ko, T.L., Hsu, Y.F., Sheu, J.R., Ou, G., Hsu, M.J., 2014. Valproic acid suppresses lipopolysaccharide-induced cyclooxygenase-2 expression via MKP-1 in murine brain microvascular endothelial cells. Biochem. Pharmacol. 88, 372–383.
Costalonga, E.C., Silva, F.M.O., Noronha, I.L., 2016. Valproic acid prevents renal dys-function and inflammation in the ischemia-reperfusion injury model. Biomed. Res. Int. 2016, 5985903.http://dx.doi.org/10.1155/2016/5985903.(10 pp).
Dezfulian, C., Raat, N.J.H., Shiva, S., Gladwin, M.T., 2007. Role of the anion nitrite in ischemia-reperfusion cytoprotection and therapeutics. Cardiovasc. Res. 75, 327–338.
Draper, H.H., Hadley, M., 1990. Malondialdehyde determination as index of lipid per-oxidation. Methods Enzymol. 186, 421–431.
Fessler, E.B., Chibane, F.L., Wang, Z., Chuang, D.M., 2013. Potential roles of HDAC in-hibitors in mitigating ischemia-induced brain damage and facilitating endogenous regeneration and recovery. Curr. Pharm. Des. 19, 5105–5120.
Grabiec, A.M., Tak, P.P., Reedquist, K.A., 2011. Function of histone deacetylase inhibitors in inflammation. Crit. Rev. Immunol. 31, 233–263.
Green, L.C., Wagner, D.A., Godowsky, J., Skipper, P.L., Wishnok, J.S., Tannenbaum, S.R., 1982. Analysis of nitrate, nitrite and [15N] nitrate in biologicalfluids. Anal. Biochem. 126, 131–138.
Halili, M.A., Andrews, M.R., Sweet, M.J., Fairlie, D.P., 2009. Histone deacetylase in-hibitors in inflammatory disease. Curr. Top. Med. Chem. 9, 309–319.
Hall, A.C., Brennan, A., Goold, R.G., Cleverley, K., Lucas, F.R., Gordon-Weeks, P.R., Salinas, P.C., 2002. Valproate regulates GSK-3-mediated axonal remodeling and sy-napsin I clustering in developing neurons. Mol. Cell. Neurosci. 20, 257–270.
Harwood, A.J., Agam, G., 2003. Search for a common mechanism of mood stabilizers.
GSK3 (TC, x200)
SO
ISC
ISC+VA100
B
Biochem. Pharmacol. 66, 179–189.
Jellet, A.P., Jenks, K., Lucas, M., Scott, R.C., 2014. Standard dose valproic acid does not cause additional cognitive impact in a rodent model of intractable epilepsy. Epilepsy Res. 110, 88–94.
Jin, R., Yang, G., Li, G., 2010. Inflammatory mechanisma in ischemic stroke: role of
in-flammatory cells. J. Leukoc. Biol. 87, 779–789.
Jin, W., Uscinska, M., Ma, Y., 2014. Review of double mood stabilizer treatments for bipolar disorder in China. Open J. Psychiat. 4, 1–4.
Jope, R.S., Yuskaitis, C.J., Beurel, E., 2007. Glycogen synthase kinase-3 (GSK3):
in-flammation, diseases, and therapeutics. Neurochem. Res. 32, 577–595.
Jung, B.H., Kim, B.J., Lee, M.S., Lee, J.H., Oh, J.H., Lee, Y.J., 2010. Dose-dependent pharmacokinetics of toxic metabolites is not related to increased toxicity following high-dose valproic acid in rats. J. Appl. Toxicol. 30, 775–778.
Kanekar, S.G., Zacharia, T., Roller, R., 2012. Imaging of stroke: part 2, pathophysiology at the molecular and cellular levels and corresponding imaging changes. Am. J. Roentgenol. 198, 63–74.
Kim, H.J., Rowe, M., Ren, M., Hong, J.-S., Chen, P.-S., Chuang, D.-M., 2007. Histone deacetylase inhibitors exhibit anti-inflammatory and neuroprotective effects in a rat permanent ischemic model of stroke: multiple mechanisms of action. J. Pharmacol. Exp. Ther. 321, 892–901.
Kramer, M., Dang, J., Baerting, F., Denecke, B., Clamer, T., Kirsch, C., Beyer, C., Kipp, M., 2010. TTC staining of damaged brain areas after MCA occlusion in the rat does not constrict quantitative gene and protein analyses. J. Neurosci. Methods 187, 84–89.
Lakhan, S.E., Kirchgessner, A., Hofer, M., 2009. Inflammatory mechanisms in ischemic stroke: therapeutic approaches. J. Transl. Med. 7, 97.http://dx.doi.org/10.1186/ 1479-5876-7-97.
Lee, M.S., Lee, Y.J., Kim, B.J., Shin, K.J., Chung, B.C., Baek, D.J., Jung, B.H., 2009. The relationship between glucuronide conjugate levels and hepatotoxicity after oral ad-ministration of valproic acid. Arch. Pharm. Res. 32, 1029–1035.
Lee, J.Y., Maeng, S., Kang, So.R., Choi, H.Y., Oh, T.H., Ju, B.G., Yune, T.Y., 2014. Valproic acid protects motor neuron death by inhibiting oxidative stress and endoplasmic reticulum stress-mediated cytochrome c release after spinal cord injury. J. Neurotrauma 31, 582–594.
Leng, Y., Liang, M.-H., Ren, M., Marinova, Z., Leeds, P., Chuang, D.-M., 2008. Synergistic neuroprotective effects of lithium and valproic acid or other histone deacetylase in-hibitors in neurons: roles of glycogen synthase kinase-3 inhibition. J. Neurosci. 28, 2576–2588.
Love, S., 1999. Oxidative stress in brain ischemia. Brain Pathol. 9, 119–131.
Morris, R.G.M., 1981. Spacial localization does not require the presence of local cues. Learn. Motiv. 12, 239–260.
Pang, T., Wang, Y.-J., Gao, Y.-X., Xu, Y., Li, Q., Zhou, Y.-B., Xu, L., Huang, Z.-J., Liao, H., Zhang, L.-Y., Gao, J.-R., Ye, Q., Li, J., 2016. A novel GSK-3βinhibitor YQ138 prevents neuronal injury induced by glutamate and brain ischemia through activation of the Nrf2 signaling pathway. Acta Pharmacol. Sin. 37, 741–752.
Pinheiro, R.M.C., Lima, M.N.M., Portal, B.C.D., Busato, S.B., Falavigna, L., Ferreira, R.D.P., Paz, A.C., Aguiar, B.W., Kapczinski, F., Schröder, N., 2015. Long-lasting re-cognition memory impairment and alterations in brain levels of cytokines and BDNF induced by maternal deprivation: effects of valproic acid and topiramate. J. Neural Transm. 122, 709–719.
Prakash, R., Carmichael, S.T., 2015. Blood-brain barrier breakdown and neovascular-ization processes after stroke and traumatic brain injury. Curr. Opin. Neurol. 28, 556–564.
Prut, L., Belzung, C., 2003. The open-field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. Eur. J. Pharmacol. 463, 3–33.
Qing, H., He, G., Ly, P.T.T., Fox, C.J., Staufenbiel, M., Cai, F., Zhang, Z., Wei, S., Sun, X., Chen, C.-H., Zhou, W., Wang, K., Song, W., 2008. Valproic acid inhibits Aβ produc-tion, neurite plaque formaproduc-tion, and behavioral deficits in Alzheimer's disease mouse models. J. Exp. Med. 205 (12), 2781–2789.
Rem, M., Leng, Y., Jeong, M., Leeds, P.R., Chuang, D.M., 2004. Valproic acid reduces brain damage induced by transient focal cerebral ischemia in rats: potential roles of histone deacetylase inhibition and heat schock protein induction. J. Neurochem. 89, 1358–1367.
Renoux, C., Dell'Aniello, S., Saarela, O., Filion, K.B., Boivin, J.-F., 2015. Antiepileptic drugs and the risk of ischemic stroke and myocardial infarction: a population-based cohort study. BMJ Open 2015 (5), e008365. http://dx.doi.org/10.1136/bmjopen-2015-008365.
Rowe, M.K., Wiest, C., Chuang, D.M., 2007. GSK3 is a viable potential target for ther-apeutic intervention in bipolar disorder. Neurosci. Biobehav. Rev. 31, 920–931.
Saito, A., Maier, C.M., Narasimhan, P., Nishi, T., Song, Y.S., Yu, F., Liu, J., Lee, Y.S., Nito, C., Kamada, H., Dodd, R.L., Hsieh, L.B., Hassid, B., Kim, E.E., González, M., Chan, P.H., 2005. Oxidative stress and neuronal death/survival signaling in cerebral ischemia. Mol. Neurobiol. 31, 105–116.
Shakespear, M.R., Halili, M.A., Irvine, K.M., Fairlie, D.P., Sweet, M.J., 2011. Histone deacetylases as regulators of inflammation and immunity. Trends Immunol. 32, 335–343.
Shi, Y., Leak, R.K., Keep, R.F., Chen, C., 2016. Translational stroke research on blood-brain barrier damage: challenges, perspectives, and goals. Transl Stroke Res 7, 89–92.
Shichita, T., Sakaguchi, R., Suzuki, M., Yoshimura, A., 2012. Post-ischemic inflammation
in the brain. Front. Immunol. 3, 132.http://dx.doi.org/10.3389/fimmu.2012.00132.
(May 2012).
Sims, N.R., Muyderman, H., 2010. Mitochondria, oxidative metabolism and cell death in stroke. Biochim. Biophys. Acta 1802, 80–91.
Sinn, D.I., Kim, S.J., Chu, K., Jung, K.H., Lee, S.T., Song, E.C., Kim, J.M., Park, D.K., Kun, L.S., Kim, M., Roh, J.K., 2007. Valproic acid-mediated neuroprotection in in-tracerebral hemorrhage via histone deacetylase inhibtion and transcription activa-tion. Neurobiol. Dis. 26, 464–472.
Sintoni, S., Kurtys, E., Scandaglia, M., Contestabile, A., Monti, B., 2013. Chronic valproic acid administration impairs contextual memory and dysregulates hippocampal GSK-3βin rats. Pharmacol. Biochem. Behav. 106, 8–15.
Stertz, L., Fries, G.R., Aguiar, B.W., Pfaffenseller, B., Valvassori, S.S., Gubert, C., Ferreira, C.L., Moretti, M., Ceresér, K.M., Kauer-Sant'Anna, M., 2014. Histone deacetylase activity and brain-derived neurotrophic factor (BDNF) in a pharmacological model of mania. Rev. Bras. Psiquiatr. 36, 39–46.
Suda, S., Katsura, K.-I., Kanamaru, T., Saito, M., Katayama, Y., 2013. Valproic acid at-tenuates ischemia-reperfusion injury in the rat brain through inhibition of oxidative stress and inflammation. Eur. J. Pharmacol. 707, 26–31.
Umka, J., Mustafa, S., ElBeltagy, M., Thorpe, A., Latif, L., Bennet, G., Wigmore, P.M., 2010. Valproic acid reduces spatial working memory and cell proliferation in the hippocampus. Neuroscience 166, 15–22.
Valerio, A., Bertolotti, P., Delbarba, A., Perego, C., Dossena, M., Ragni, M., Spano, P., Carruba, M.O., De Simoni, M.G., Nisoli, E., 2011. Glycogen synthase kinase-3 in-hibition reduces ischemic cerebral damage, restores impaired mitochondrial bio-genesis and prevents ROS formation. J. Neurochem. 116, 1148–1159.
Venna, V.R., Benashski, S.E., Chauhan, A., McCullough, L.D., 2015. Inhibition of glycogen synthase kinase-3βenhances cognitive recovery after stroke: the role of TAK1. Learn. Mem. 22, 336–343.
Volmar, C.-H., Wahlestedt, C., 2014. Histone deacetylases (HDACs) and brain function. Neuroepigenetics 2014.http://dx.doi.org/10.1016/jnepig.2014.10.002.
Vorhees, C.V., Williams, M.T., 2006. Morris water maze: procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 1, 848–858.
Vriend, J.P., Alexiuk, N.A.M., 1996. Effects of valproate on amino acid and monoamine concentrations in striatum of audiogenic seizure-prone balb/c mice. Mol. Chem. Neuropathol. 27, 307.
Wang, Z., Tsai, L.-K., Munasinghe, J., Leng, Y., Fessler, E.B., Chibane, F., Leeds, P., Chuang, D.-M., 2012. Chronic valproate treatment enhances post-ischemic angio-genesis and promotes functional recovery in a rat model of ischemic stroke. Stroke 43, 2430–2436.
Wu, X., Li, S., Wu, Q., Peng, Y., Yu, D., Wang, H., Chui, D., Zhao, J., 2013. Histone deacetylase inhibition leads to neuroprotection through regulation on glial function. Mol. Neurodegener. 8 (Suppl. 1), P49.
Ximenes, J.C.M., Lima Verde, E.C., Naffah-Mazzacoratti, M.G., Viana, G.S.B., 2012. Valproic acid, a drug with multiple molecular targets related to its potential neuro-protective action. Neurosci. Med. 3, 107–123.
Ximenes, J.C.M., de Oliveira Gonçalves, D., Siqueira, R.M., Neves, K.R., Santos Cerqueira, G., Correia, A.O., Félix, F.H., Leal, L.K., de Castro Brito, G.A., da Graça Naff ah-Mazzacorati, M., Viana, G.S., 2013. Valproic acid: an anticonvulsant drug with potent antinociceptive and anti-inflammatory properties. Naunyn Schmiedeberg's Arch. Pharmacol. 386, 575–587.
Ximenes, J.C.M., Neves, K.R.T., Leal, L.K.M.A., do Carmo, M.R.S., Brito, G.A.C., Naff ah-Mazzacoratti, M.G., Cavalheiro, E.A., Viana, G.S.B., 2015. Valproic acid neuropro-tection in the 6-OHDA model of Parkinson's disease is possible related to its anti-inflammatory and HDAC inhibitory properties. J. Neurodegenerative Dis. 2015, 313702.http://dx.doi.org/10.1155/2015/313702.
Yao, Z.-G., Liang, L., Liu, Y., Zhang, L., Zhu, H., Huang, L., Qin, C., 2014. Valproate improves memory deficits in Alzheimer's disease mouse model: investigation of possible mechanisms of action. Cell. Mol. Neurobiol. 34, 805–812.
Yuskaitis, C.J., Jope, R.S., 2009. Glycogen synthase kinase-3 regulates microglial mi-gration, inflammation, and inflammation-induced neurotoxicity. Cell. Signal. 21, 264–273.
Zhang, Z., Zhang, Z.Y., Fausae, U., Schluesenger, H.J., 2008. Valproic acid attenuates inflammation in experimental autoimmune neuritis. Cell. Mol. Life Sci. 65, 4055–4065.
Zhang, Z., Zhang, Z.Y., Wu, Y., Schluesener, H.J., 2012. Valproic acid ameliorates
in-flammation in experimental autoimmune encephalomyelitis rats. Neuroscience 221, 140–150.
Zhang, C., Yuan, X., Hu, Z., Liu, S., Li, H., Wu, M., Yuan, J., Zhao, Z., Su, J., Wang, X., Liao, Y., Liu, Q., 2017. Valproic acid protects primary dopamine neurons from MPP+
- induced neurotoxicity: involvement of GSK3βphosphorylation by Akt and ERK trough the mitochondrial intrinsic apoptotic pathway. Biomed. Res. Int. 2017, 8124501 (12 pp).
Zheng, Q., Liu, W., Liu, Z., Zhao, H., Han, X., Zhao, M., 2014. Valproic acid protects septic mice from renal injury by reducing the inflammatory response. J. Surg. Res. 192, 163–169.