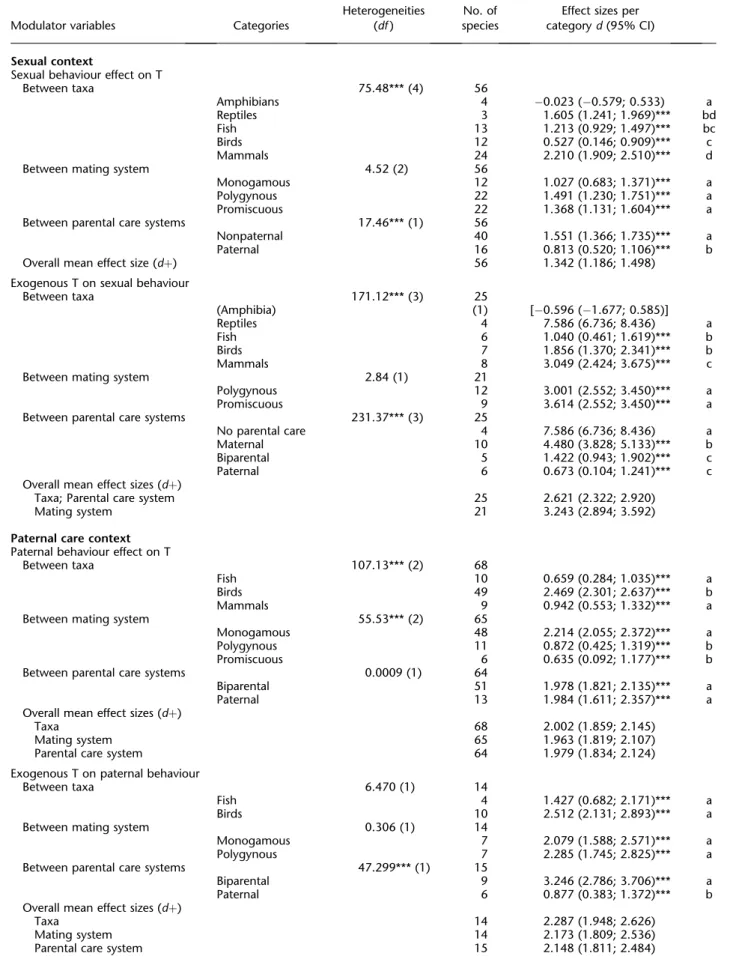
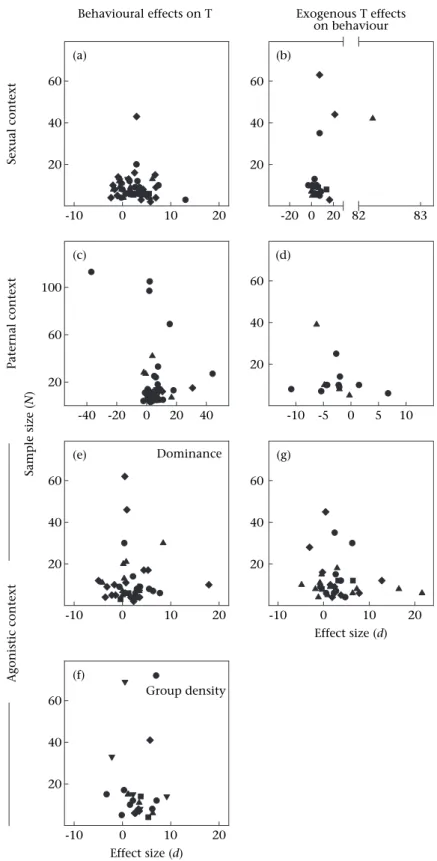
Social modulation of androgens in male vertebrates: Meta-analyses of the challenge hypothesis
Texto
Imagem

Documentos relacionados
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
Tal como no Turismo Rural, como referido no ponto 1.2, o Ecoturismo, é estruturado a partir das atividades no meio Natural, embora neste meio, pretenda-se, impulsionar
Quanto às características do solo, as diferenças mais destacáveis são, na preparação mecânica, uma maior profundidade efectiva do solo, um aumento dos
Assim o conhecimento se constitui num processo de homem com o mundo, como um conjunto de sistematizações no qual o homem produz para além de se comunicar, sobreviver e
A tríade clínica clássica da NEME é composta por dor lombar ou de membros inferiores (MMII), disfunção vesical e fraqueza de MMII. Qualquer paciente apresentando estes sintomas
A better identification of the treatment group through the cross gathering of missing data, by using different sources together with Quadros de Pessoal; The
As doenças mais frequentes com localização na cabeça dos coelhos são a doença dentária adquirida, os abcessos dentários mandibulares ou maxilares, a otite interna e o empiema da
Os controlos à importação de géneros alimentícios de origem não animal abrangem vários aspetos da legislação em matéria de géneros alimentícios, nomeadamente
