Molecular evolution and population genetics of a Gram-negative binding protein gene in the malaria vector Anopheles gambiae (sensu lato)
Texto
Imagem

Documentos relacionados
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
LAERG-Laboratório de Ergonomia-UTFPR: Av. Faculdades Integradas Santa Cruz de Curitiba. Apresentação da pesquisa. V ocê está sendo convidado a participar de uma pesquisa que vai
This network model is built from past measurements of two parameters: the traffic that enters and leaves the network at the access points, and the corresponding QoS parameters. The
Inhibitory properties of the antibody response to Plasmodium vivax Duffy binding protein in an area with unstable malaria transmission.. Genetic diversity
In this work we studied 18 patients with PWS, 20 with AS, 3 with supernu- merary marker chromosomes (SMC), and their parents, by GTG banding and FISH techniques: 14 PWS patients
Hong and coworkers (26) combined adenoviral capsid proteins or adenoviral fi- ber proteins with liposomes, obtaining a con- siderable increase in the efficiency of trans- fection of
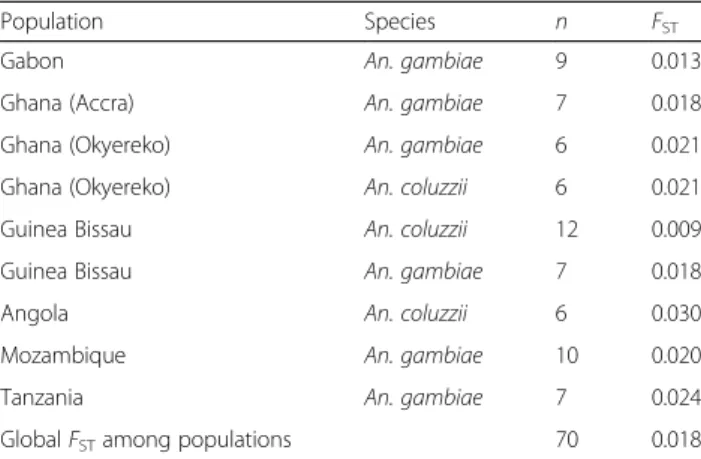
This work aims to evaluate and understand the patterns of inter- and intrapopulation genetic diversity, in order to determine the gene fl ow and study the spatial distribution
