Effect of simulated microgravity on the cell cycle in cultured cells of Arabidopsis thaliana
Texto
Imagem

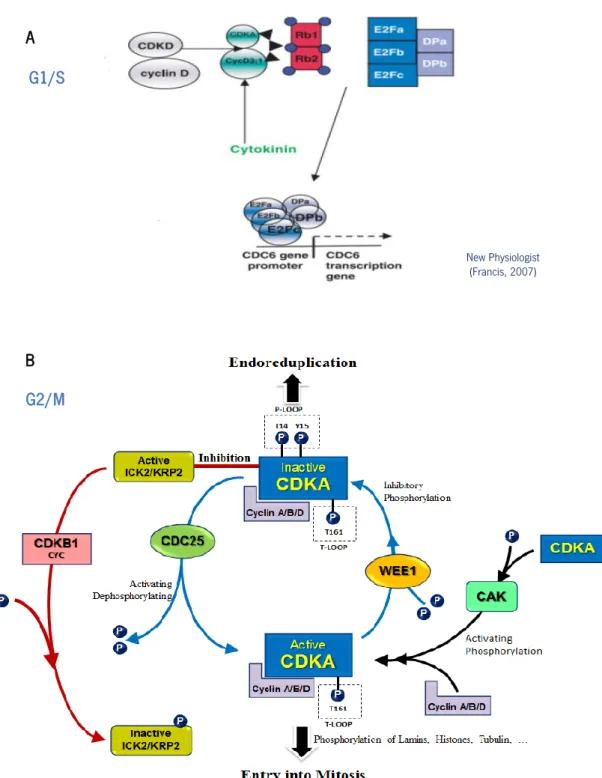
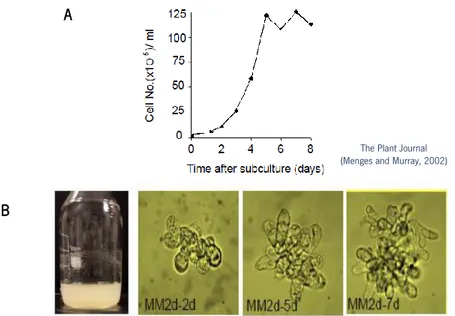
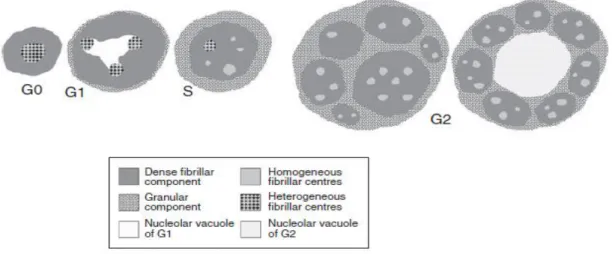

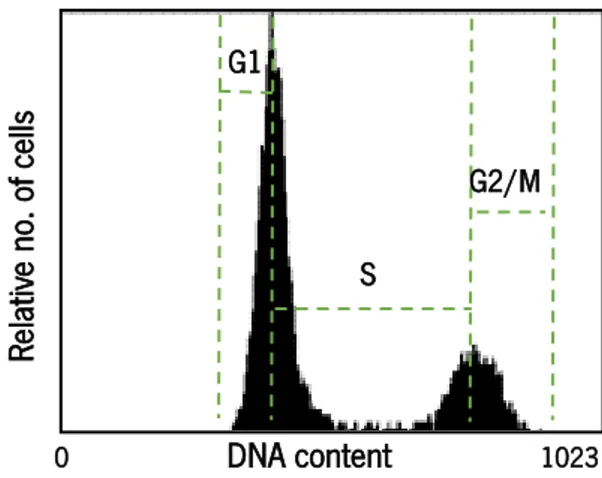
Documentos relacionados
Class 0: no damage; Class 1: little damage with a tail length shorter than diameter of the nucleus; Class 2: medium damage with a tail length one or two times the diameter of
The suppression of cancer cell growth has been known to be primarily mediated by apoptosis and cell cycle arrest. We next applied flow cytometry to analyze cell cycle phase
Effects of polydatin on cell apoptosis in liver cancer cell lines treated with different concentrations of polydatin for 48 h (A), determined by fl ow cytometry assay; B, effect of
Cell cycle analysis of A2058 cells grown for 72 h in control cell culture medium (A) or cell culture medium containing zeaxanthin 40 M (B). The majority of control or treated
Surpri- singly, we found that Fas expression positively correlates to spontaneous lymphoproliferation in vitro ( Figure 6A ), which might imply that the observed defect
Analysis and discrimination of necrosis and apoptosis (programmed cell death) by multiparameter flow cytometry.. Novel approach for simultaneous evaluation of cell phenotype,
The effects of a 72-h exposure to 50 µM 13- cis retinoic acid on HC11 and HC11ras cell proliferation and HC11 cell differentiation were investigated by flow cytometric cell
In addition, SNHG12 overexpression boosted the cell cycle progression of SW480 cells transfected with pcDNA- SNHG12 and SNHG12 knockdown inhibited the cell cycle progression of
