Functional abilities of cultivable plant growth promoting bacteria associated
with wheat (
Triticum aestivum
L.) crops
Fernanda da S. Moreira
1,#, Pedro B. da Costa
1,#, Rocheli de Souza
1, Anelise Beneduzi
2, Bruno B. Lisboa
2,
Luciano K. Vargas
2and Luciane M. P. Passaglia
11
Departamento de Genética, Instituto de Biociências, Universidade Federal do Rio Grande do Sul,
Porto Alegre, RS, Brazil.
2
Fundação Estadual de Pesquisa Agropecuária, Porto Alegre, RS, Brazil.
Abstract
In the pursuit of sustainable agriculture, bioinoculants usage as providers of a crop’s needs is a method to limit envi-ronmental damage. In this study, a collection of cultivable putative plant growth promoting (PGP) bacteria associated with wheat crops was obtained and this bacterial sample was characterized in relation to the functional diversity of certain PGP features. The isolates were obtained through classical cultivation methods, identified by partial 16S rRNA gene sequencing and characterized for PGP traits of interest. Functional diversity characterization was per-formed using Categorical Principal Component Analysis (CatPCA) and Multiple Correspondence Analysis (MCA). The most abundant genera found among the 346 isolates werePseudomonas, Burkholderia, and Enterobacter. Oc-currence of PGP traits was affected by genus, niche, and sampling site. A large number of genera grouped together with the ability to produce indolic compounds; phosphate solubilization and siderophores production formed a sec-ond group related to fewer genera, in which the genusBurkholderia has a great importance. The results obtained may help future studies aiming prospection of putative plant growth promoting bacteria regarding the desired organ-ism and PGP trait.
Keywords: plant growth promoting bacteria; Categorical Principal Component Analysis; functional analysis;Burkholderiagenus.
Received: May 27, 2015; Accepted: August 24, 2015.
Introduction
Since the description of the association of non-sym-biotic diazotrophic bacteria with non-legume plants (Baldani et al., 1983), free-living bacteria have demon-strated positive effects in promoting plant growth, proving their agronomic potential not only through biological nitro-gen fixation (e.g., James et al., 2002; Ambrosini et al., 2012). These organisms present in the rhizosphere, root surfaces or inside plant tissues are able to directly or indi-rectly induce plant growth, as well as resistance to or pro-tection against pathogens (Lugtenberg and Kamilova, 2009). The combination of nitrogen fixation with other fea-tures of plant growth promoting (PGP) has shown that all these benefic bacteria, collectively called plant growth pro-moting bacteria (PGPB), are an efficient and viable alterna-tive to chemical fertilizers to achieve maximum production with environmental conservation (Kennedyet al., 2004).
Plant traits shape the conditions for microbial coloni-zation mostly due to organic compounds released from the roots. A fine coordinated interaction between the variety of exudates excreted by the plant and individual characteris-tics of distinct microbial populations is a crucial aspect of driving selection (Denniset al., 2010). However, from bac-teria selected for rhizosphere colonization, only a small parcel can actually be commercially used. In addition, con-cerning biotechnological purposes, the selective effects on the rhizosphere microbiota is expected to be very complex. Only at the right set of conditions, a targeted effect on the behavior of a certain microbial subpopulation,e.g.an intro-duced inoculum, which is supposed to get established and interactive with the plant root, is possible (Hartmannet al., 2009).
While species diversity comprises richness and equi-tability (i.e., the total number of species and distribution among species, respectively), functional diversity is the value and range of the functional traits of the organisms in a given ecosystem (Tilman, 2001). Functional diversity af-fects ecosystem dynamics, stability, productivity and nutri-tional balance, among other aspects of a functioning ecosystem (Tilman, 2001). Joint investigation of cultivable Send correspondence to Luciane M. P. Passaglia, Departamento
de Genética, Instituto de Biociências, UFRGS, Av. Bento Gonçal-ves, 9500, Caixa Postal 15.053, Prédio 43312, sala 207b, 91501-970 Porto Alegre, RS, Brazil. E-mail: [email protected] # F. S. M and P. B. C. contributed equally to this work and should both be considered as first author.
(by traditional isolation) and non-cultivable diazotrophic and PGP bacteria can enable the description and analysis of almost the entire composition and structure of active plant growth promoting bacterial communities (Wartiainen et al., 2008). For biotechnological applications, the character-ization of these organisms relies more often on culture-dependent methods to search, characterize and better un-derstand the diversity of functions involved in promoting plant growth. Numerous studies have demonstrated the het-erogeneity in composition of bacterial community structure of bulk soil relative to the rhizospheric bacterial commu-nity, such as between root zones (Denniset al., 2010). The objectives of this study were to isolate, identify and charac-terize a large number of putative diazotrophic and PGP bac-teria associated with wheat crops. We were able to characterize this specific bacterial population concerning the functional diversity of its PGP features associated to the sampling sites and colonization niche (either roots or rhizospheric soil), identifying in which way the heteroge-neity of PGP traits was presented. The associations here suggested may help in further studies on bioprospection that covers the interactions between genera, their roles (dif-ferent characteristics of PGP), and niche occupancy.
Materials and Methods
Sampling and location
Wheat plants (Triticum aestivum L., cv. Guamirim, three plants from each sampling site) and their respective rhizospheric soils were collected from six wheat-producing regions in Rio Grande do Sul, Brazil: São Borja (SB; 28°39’39’’S, 56°00’14’’W), Júlio de Castilhos (JC; 29°13’37’’S, 53°40’54’’W), Vacaria (VA; 28°30’43’’S, 50°56’02’’W), Campina das Missões (CM; 27°59’20’’S, 54°50’22’’W), Guarani das Missões (GM; 28°08’27’’S, 54°33’29’’W), and Boa Vista do Cadeado (BV; 28°35’06’’S, 53°47’57’’W). Sampling of mature crops was performed in September: in 2010 for the SB, JC, and VA lo-calities; and in 2011 for the CM, GM, and BV localities. All sites were maintained under standard fertilization
condi-tions for wheat culture for over ten years. Three soil sam-ples from each sampling site were also collected at 20 cm depth, bulked and analyzed for pH, phosphorus, potassium, organic matter, and clay contents using standard methods (Table 1; Helmke and Sparks, 1996; Kuo, 1996; Mulvaney, 1996; Nelson and Sommers, 1996).
Isolation of putative diazotrophic bacteria
Putative diazotrophic bacteria were isolated from roots and rhizospheric soil of three sampled plants (that were pooled to compose one composite sample) from each sampling site. The roots were superficially disinfested by immersing in 70% ethanol for 1 min and 4% hypochlorite solution for 2 min followed by rinsing five times with ster-ile distilled water. After disinfestation, roots were sliced with a sterile scalpel. Ten grams of sliced roots and rhizospheric soil (manually detached from the roots) from each sampling site were separately placed into sterile 250-ml Erlenmeyer flasks containing 90 mL of sterile sa-line solution (0.85% NaCl). Isolation of bacteria was per-formed according to Döbereiner (1988). Briefly, the samples were incubated at 4 °C with agitation (125 rpm) for 4-6 h, subjected to serial dilutions in saline solution, inocu-lated and reinocuinocu-lated for seven days into semi-solid, nitro-gen-free media NFb, LGI, and LGI-P to allow pellicles formation, and finally streaked onto agar plates supple-mented with a nitrogen source (yeast extract). Bacterial iso-lates were randomly selected from the piso-lates. Each isolate was inoculated into liquid LB medium (Sambrook and Rus-sel, 2001) at 28 °C under agitation (200 rpm) and bacterial cultures were stained using the Gram method to certify their purity; after that, the isolates were stored in 20% ster-ile glycerol at -20 °C.
Molecular identification
Extractions of genomic DNA from bacterial cultures grown in LB medium, for 72 h, at 28 °C were performed us-ing a phenol-chloroform and ethanol precipitation protocol as described by Sambrook and Russel (2001).
Table 1- Soil abiotic characteristic at sampling sites.
Pa Kb Clay OMCc pH
Sampling site mg kg-1 % H2O
Júlio de Castilhos 24.3 166 38 3 5.1
São Borja 13.5 158 26 4.8 5.3
Vacaria 6.5 151.3 42.9 5.6 6.0
Campina das Missões 18.9 167 36 2.3 5.4
Guarani das Missões 27.2 148 37 3.7 5.8
Boa Vista do Cadeado 9.6 86 61 2.8 5.5
aP = extractable phosphorus; b
K = extractable potassium;
The genomic DNA extracted from all isolates was submitted to partial 16S rRNA gene amplification (~450 bp) using primers U968 (AACGCGAAGAA CCTTAC) and L1401 (CGGTGTGTACAAGACCC, Felske et al., 1997). Amplification reactions were per-formed using approximately 50 ng of DNA template in 25mL reactions containing 20mM of each dNTP (Ludwig
Laboratories), 0.05mM of each primer, 1 mM MgCl2 and 1
U Taq polymerase (Invitrogen®) in 1 X Taq Buffer. Ampli-fication of 16S rRNA gene portions was carried out under the following conditions: an initial denaturation step at 94 °C for 5 min, 30 cycles at 94 °C for 45 s, 52 °C for 45 s, 72 °C for 45 s, and a final elongation cycle at 72 °C for 10 min. Sanger sequencing was performed on a 3500xL Genetic Analyzer (Applied Biosystems®) using the DY EnamicTM ET Dye Terminator Cycle Sequencing Kit (GE HealthCare).
Sequences were trimmed to exclude low quality se-quenced nucleotides. Data obtained from sequencing were compared to sequences available from the EzTaxon-e Server, which is an extension of the EzTaxon database. The nucleotide sequences of the partial 16S rRNA gene seg-ments determined in this study have been deposited in the GenBank database (accession numbers KC254895 to KC255222).
Evaluation of plant growth promotion characteristics
All isolates were evaluated for their capacity to pro-duce indolic compounds (ICs) and siderophores, as well as their ability to solubilize phosphate (as tricalcium phos-phate) using three replicates and appropriate controls (i. e. non-inoculated media, inoculated media with known pro-ducers/solubilizers strains).
The production of ICs was evaluated according to Glickmann and Dessaux (1995) and Ambrosini et al. (2012). ICs production was ranked in levels as follows: bacterial isolates producing less than 17mg of ICs ml-1were
considered level 1 ICs producers (IC1); those that produced between 17mg and 80mg of ICs ml-1were considered level
2 ICs producers (IC2); and those that produced more than 80mg of ICs ml-1were considered level 3 ICs producers
(IC3).
Siderophores production was evaluated according to Schwyn and Neilands (1987). After incubation for 48-72 h at 28 °C, the formation of a yellow, orange or violet halo around the colonies denoted their ability to produce and re-lease siderophore molecules. Likewise, a clear halo around a colony indicated that it was able to solubilize the phos-phate source (provided as tricalcium phosphos-phate) in glucose yeast (GY) medium as described by Ambrosini et al. (2012). Evaluation of these two abilities was conducted based on the size of the halo formed around bacterial colo-nies: isolates without a halo (halo = 0 cm) were labeled ei-ther non-siderophore producers (level 1 – Sid1) or non-phosphate solubilizers (level 1 – Phos1); isolates with
a halo wider than 0 cm up to 1 cm were considered either level 2 siderophore producers (Sid2) or level 2 phosphate solubilizers (Phos2), and isolates with a halo bigger than 1 cm were considered either level 3 siderophore producers (Sid3) or level 3 phosphate solubilizers (Phos3).
Statistical analysis
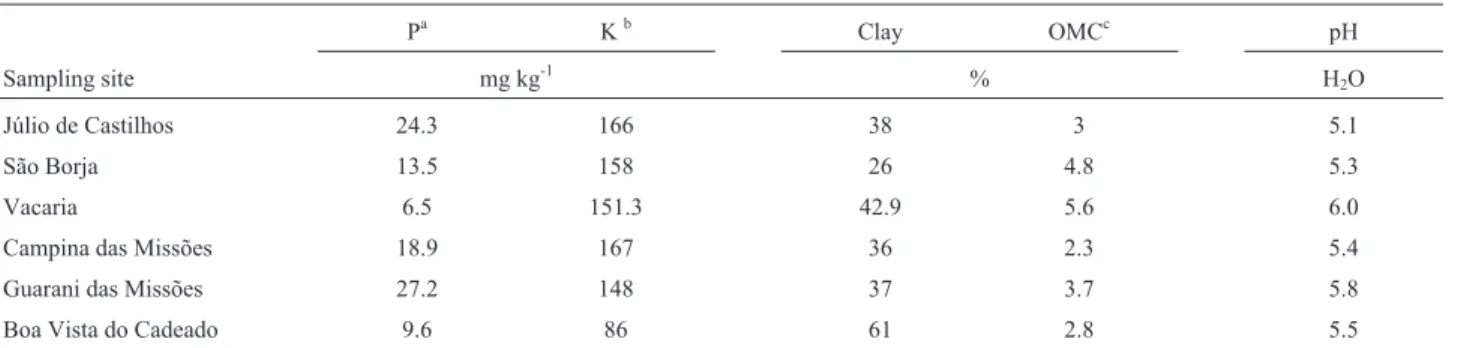
The composition of the bacterial sample communi-ties, identified by the bacterial genera at each sampling site for both roots and rhizospheric soils (referred to here as niches) was used to determine the Shannon diversity index (H‘, Shannon and Weaver, 1949). The correlation between soil properties and microbial genetic diversity was deter-mined via Principal Component Analysis (PCA) using a correlation matrix (with mean centering) to calculate eigen-values and eigenvectors (Figure 1).
To better understand and facilitate the visualization of functional diversity, the categorical data (i.e., genera, ranked PGP traits, sampling sites, and niches) was analyzed by Categorical Principal Component Analysis (CatPCA) and Multiple Correspondence Analysis (MCA). MCA is similar to PCA for qualitative data accounting only for mul-tiple nominal variables, without ordering them in a scale, such as in the case of genera and sampling sites. CatPCA, on the other hand, was used to account for the ordination of the different PGP levels, mixed with multiple nominal vari-ables such as genera. Both methods provide easy represen-tation and interprerepresen-tation of categorical data by reducing a large number of variables into a two-dimensional map,
sidering the frequency and homogeneity of the events to connect points (Hoffman, 1992). Thus, the generated hy-pothesis (null hyhy-pothesis of homogeneity) can be tested through a chi-square test via a contingency table and ad-justed residual analysis, as in this study. The differences were considered significant for standardized adjusted resi-dues <1.96 . We used IBM SPSS Statistics to build the MCA and CatPCA analyses. On the input table for both fig-ures, each line was representative of a single isolate. For MCA (Figure 2), both PGP traits and genera information are held in the columns (Table S1), while for CatPCA (Fig-ure 3) the columns are for genera, niche and sampling site (Table S2). Each dot in the plot represents the average posi-tion for the different categories of every variable.
Results
Isolation, identification and genera diversity of putative diazotrophic bacteria
A total of 346 bacterial isolates were obtained from the six sampling sites. Among them, 328 were assigned to 36 distinct genera (according to the closest match in the
Ez-taxon database, considering a minimum of 97% of simi-larity, shown in Table 2), while 18 remained unidentified.
The most abundant and the only bacterial genus iso-lated in all the sampling sites and niches wasPseudomonas (46 isolates), representing 22% of all isolates, followed by strains belonging to the genera Burkholderia (17), Enterobacter(16), andStenotrophomonas(12). These four genera were also the most ubiquitous, represented in at least five of the six sampling sites, comprising almost 60% of the bacteria isolated. The putative diazotrophic bacterial com-munity analyzed was composed by 32 other genera, with the number of representative strains ranging from one to nine isolates, including various genera belonging to the Enterobacteriaceae family, such asCedecea(8),Leclercia (8), andKlebsiella(4); and to the Xanthomonadaceae fam-ily, such asXanthomonas(6) andDyella(3). Gram positive bacteria were represented by four isolates belonging to the Microbacteriumgenus. Distribution of genera by sampling site and niche and their diversity indices are shown in Table 2.
Figure 2- Multiple Correspondence Analysis (MCA) of levels of PGP traits and genera. IC = indolic compounds production, Phos = phosphate
Evaluation of plant growth promotion traits
All isolates were evaluated for their ability to produce siderophores and indolic compounds and to solubilize phosphate under the given conditions. The results are listed in Table 3.
Phosphate solubilization was the rarest ability among the isolates, having occurred in 55 out of 346 isolates. Strains identified as phosphate solubilizers mainly be-longed to the genusBurkholderia. The ability to produce siderophores was a common feature to several strains, espe-cially of those belonging to the generaPseudomonasand Burkholderia. Siderophores production was an ability also identified in several strains belonging to the Entero-bacteriaceae family (Cedecea,Leclercia,Hafnia,Serratia, Klebsiella,Pantoea,andShigella). While phosphate solu-bilization ability was more often found in isolates from rhizospheric soil, siderophore production was found in iso-lates more closely related to the roots. All isoiso-lates were able to produce indolic compounds, but with variable effi-ciency, ranging from 0.1mg to 640 mg of ICs ml-1. The
greatest IC producers belonged to the generaEnterobacter,
Pseudomonas, Azospirillum, Rhizobium, and Herbaspirillum. Those exhibiting all three abilities concur-rently were mainly represented by strains belonging to the genusBurkholderia (17 isolates) and to members of the Enterobacteriaceae family (12 isolates).
Soil characteristics and functional analysis of PGP traits
Soil analysis (Table 1) showed that soil conditions at all sampling sites (namely, availability of nutrients and pH) were in accordance with recommendations for wheat cul-ture. Nevertheless, when Principal Component Analysis (PCA, Figure1) was used to visualize the relationships be-tween abiotic soil parameters (P, K, clay, pH, and OM con-tents) and bacterial diversity (H‘), some differences could be observed. The first two dimensions of PCA explained 77.92% of the total variation, with principal component 1 (PC1) accounting for 46.57% and principal component 2 (PC2) for 31.35% of the variance. The major abiotic soil properties related to bacterial diversity (H‘) were pH and OM content (Figure 1).
Table 2- Heatmap associations of bacterial genera and occurrence of putative endophytic (Root) and rhizospheric (Soil) isolates. Different shades show whether a certain location had more or fewer bacteria, than expected, or whether there was no association. Black cells = number of isolates lower than ex-pected in that condition; dark shaded cells = number of isolates higher than exex-pected in that condition; light shaded cells = no significant differences be-tween observed and expected values. “–” = association could not be calculated due to lack of cases (no comparable expected total marginal values), ac-cording to adjusted residual analysis (p<0.05)
Sampling Site Júlio de Castilhos
São Borja Vacaria Campina das
Missões
Guarani das Missões
Boa Vista do Cadeado
Bacterial genera Root Soilb Root Soil Root Soil Root Soil Root Soil Root Soil
Achromobactersp. 1 - - - 1 - - 1 1 - 1
-Acinetobactersp. - - - - 1 1 - - -
-Azorhizobiumsp. - - - - 2 2 - - -
-Azospirillumsp. 1 7 1 4 1 - - -
-Burkholderiasp. - 2 - - 1 4 9 17 7 5 - 8
Caulobactersp. - - - 1 - - -
-Cedeceasp. 2 3 - 3 - - -
-Chryseobacteriumsp. - - - 1 - 1
Citrobactersp. - - - 1 -
-Dyellasp. - - - 1 - 1 - - - 1
Enterobactersp. 8 3 - 2 1 - 1 - 2 1 4 4
Hafniasp. - - - - 5 - - -
-Erwiniasp. - - 1 - - -
-Herbaspirillumsp. - - 4 1 - - 1 - - - 4
-Klebsiellasp. - - - 3 1 -
-Kluyverasp. - - - 1
Leclerciasp. - - 1 5 2 - - -
-Lysobactersp. - - - 1 - - -
-Luteibactersp. 1 1 - - - 2 -
-Microbacteriumsp. - - - - 1 - - 3 - - -
-Novosphingobiumsp. - - - - 1 - - -
-Ochrobactrumsp. 1 - - - 1 1 - - -
-Pandoraeasp. - 1 - - -
-Pantoeasp. - - - 2 - - - 1 4 2
Pedobactersp. - 1 - - -
-Pseudomonassp. 3 5 9 6 3 2 18 4 8 14 5 1
Raoultellasp. - - - - 1 - - -
-Rhizobiumsp. 6 4 5 5 - 6 - 1 1 1 -
-Salmonellasp. - - - 1 3
Serratiasp. 4 - 5 - 1 3 - - -
-Shigellasp. - - - 5
-Sphingobiumsp. - - - - 1 - - -
-Sphingomonassp. - - 1 1 1 - - -
-Stenotrophomonassp. 1 - - - - 1 3 1 5 1 3
-Variovoraxsp. 1 - - -
-Xanthomonassp. 1 - 1 1 - - - 3
-Unidentified - 2 1 1 5 5 - 1 1 2 -
-Total 30 29 29 30 29 29 32 29 28 30 30 21
H‘a 2.31 2.18 2.80 1.40 1.82 2.35
a
Shannon diversity index.
Evaluation of the relationship between genera and PGP traits allowed the observation of certain patterns of as-sociation. One finding was a division between positive and negative values of dimension 1 (associated to Sid and Phos) when analyzing genera related to PGP traits (Figure. 2). A large number of genera grouped together with the ability to produce ICs (left side of the plot); levels 2 and 3 of phos-phate solubilization ability and level 3 of siderophores pro-duction ability formed a second group (right side of the plot) and were related to fewer bacterial genera than for ICs production. The sampling site and niches (roots and rhizospheric soil) did not contribute significantly to the dif-ferences, so these data were not represented in the plot.
By focusing the analysis on the most abundant gen-era, it was observed that, regardless of the sampling site or niche, strains belonging to the genusBurkholderia were strongly related to nutrient uptake (either phosphate solubilization or siderophores production). This genus is associated with siderophores production at level 3, both in the roots and in rhizospheric soil, and also associated with phosphate solubilization at level 2 in the roots and at levels 2 and 3 in rhizospheric soil. Interestingly, this genus was
associated with poor ICs production in the rhizospheric soil (more level 1 producers and less level 3 producers than ex-pected, according to chi-square analysis), and average pro-duction in root isolates (more level 2 producers and less level 1 producers than expected). Isolates belonging to Enterobacter genus were found related to phosphate solubilization (level 3 in roots) and level 3 ICs production both in roots and in rhizospheric soil. These and other sig-nificant associations are shown on Table 3 and Figure 2.
Differences in the community structure associated to the sampling site and niche can be observed in Figure 3. Bacterial samples isolated from the JC and SB localities shared similar composition, possessing most of the strains belonging to the generaRhizobiumandAzospirillum; bac-terial samples isolated from the CM and GM localities were also associated, mainly composed of strains belonging to the genera Burkholderia, Pseudomonas, and Stenotrophomonas. The bacterial sample isolated from the BV locality was distinct from all other regions and was mainly composed of strains belonging to the genera Enterobacter and Herbaspirillum. Similar behavior was observed for the bacterial sample isolated from the VA
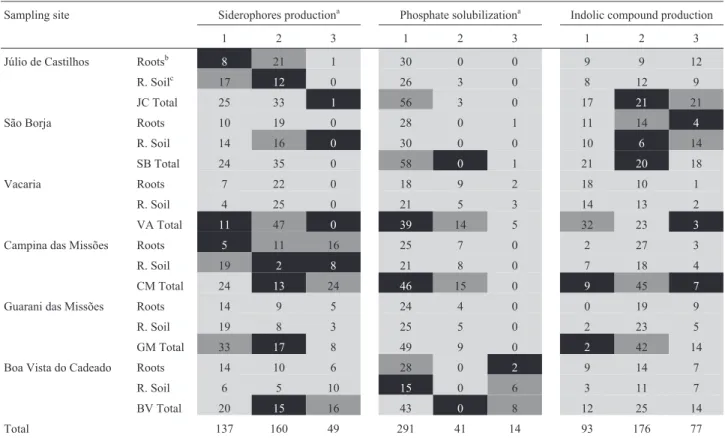
lo-Table 3- Heatmap associations of bacterial isolates and PGP traits both from roots and rhizospheric soil from each sampling site. Different shades show whether a certain location had more or fewer bacteria than expected. or whether there was no association. Black cells = number of isolates lower than ex-pected in that condition, dark shaded cells = number of isolates higher than exex-pected in that condition, light shaded cells = no significant differences be-tween observed and expected values according to adjusted residual analysis (p<0.05).
Sampling site Siderophores productiona Phosphate solubilizationa Indolic compound production
1 2 3 1 2 3 1 2 3
Júlio de Castilhos Rootsb 8 21 1 30 0 0 9 9 12
R. Soilc 17 12 0 26 3 0 8 12 9
JC Total 25 33 1 56 3 0 17 21 21
São Borja Roots 10 19 0 28 0 1 11 14 4
R. Soil 14 16 0 30 0 0 10 6 14
SB Total 24 35 0 58 0 1 21 20 18
Vacaria Roots 7 22 0 18 9 2 18 10 1
R. Soil 4 25 0 21 5 3 14 13 2
VA Total 11 47 0 39 14 5 32 23 3
Campina das Missões Roots 5 11 16 25 7 0 2 27 3
R. Soil 19 2 8 21 8 0 7 18 4
CM Total 24 13 24 46 15 0 9 45 7
Guarani das Missões Roots 14 9 5 24 4 0 0 19 9
R. Soil 19 8 3 25 5 0 2 23 5
GM Total 33 17 8 49 9 0 2 42 14
Boa Vista do Cadeado Roots 14 10 6 28 0 2 9 14 7
R. Soil 6 5 10 15 0 6 3 11 7
BV Total 20 15 16 43 0 8 12 25 14
Total 137 160 49 291 41 14 93 176 77
aCategories scored for plant growth promotion traits: 1 = non-siderophores producers, non-phosphate solubilizers or level 1 IC producers; 2 = level 2
siderophore producers, phosphate solubilizers or IC producers; 3 = level 3 siderophores producers, phosphate solubilizers or IC producers.
bBacterial isolates obtained from roots. c
cality, although this sample did not have a strong relation-ship with any particular bacterial genus. Association of the most representative genera, according to locality and whether in the roots or in rhizospheric soil, are demon-strated in Table 2.
PGP traits were also associated with locality and niche. Association between PGP traits and locality, as well as the effect of niche in each locality are shown in Table 3.
Discussion
Despite the fact that all sampling sites analyzed were traditionally used to crop wheat and that all sampled plants were at the same stage of maturity, Principal Coordinate Analysis (PCA) was able to separate the regions into dis-tinct groups according to their soil characteristics and di-versity indices. Clay, pH, and organic matter contents may exert influence on bacterial diversity and survival (Palmer and Young, 2000). Lower levels of bacterial diversity and richness might be observed in acidic soils; in a way that pH could be one of the best predictor of soil bacterial diversity and richness (Fierer and Jackson, 2006). As observed in the PCA analysis, pH is strongly associated to diversity index (H’ value). However, the bacterial sample communities from Júlio de Castilhos (JC) and São Borja (SB) localities that presented very similar pH values and also clustered to-gether with those from GM and CM localities in the PCA analysis, presented much higher diversity indices (2.31 and 2.18, respectively). This result suggested that pH should not be the only factor considered for microbiological diver-sity estimates. Although little information is available about the specific effects of parameters such as organic car-bon and total nitrogen content of the soil on microbial di-versity, information such as organic carbon and total nitrogen content could also affect the diversity of soil bac-teria, including nitrogen-fixing bacteria (Ramette and Tiedje, 2007). The Vacaria locality was the most distinctive geographical area and it also presented the lowest P con-tent, the highest organic matter concon-tent, and the highestH‘ value (2.8). Together, these features could justify its posi-tion in the PCA analysis, which was separate from all sam-ples. Similar behavior was observed for the Boa Vista do Cadeado (BV) locality, which also appeared isolated from the other sites in the PCA analysis, likely due to its most distinctive parameter, reduced organic matter content.
Various studies concerning plant-associated bacterial communities are able to shed light on factors driving micro-bial composition, especially regarding taxonomical distri-bution linked to function. 16S rRNA profiling and advanced metagenomics are powerful tools for the charac-terization of such populations (e.g.Bulgarelliet al., 2012; Lundberg et al., 2013). For example, Bulgarelli et al. (2012) conclude that 40% of the colonizing bacteria of Arabidopsis thaliana roots were due to the presence of plant cell wall residues instead of soil or plant specificity.
The isolates identified here were assigned to 36 gen-era belonging to 17 families. The most abundant family was Pseudomonadaceae, which comprised strains belonging to the most abundant genus identified,Pseudomonas. Species from this genus are known as nitrogen-fixing bacteria in various ecosystems and are commonly isolated from the rhizosphere of gramineae plants (Mirza et al., 2006; Ambrosini et al., 2012). Germida and Siciliano (2001) studied microbial communities from old, recent and mod-ern wheat cv. and observed that modmod-ern cv., (such as Guamirim), are more aggressively colonized by Pseudo-monas strains. Enterobacteriaceae was the second most abundant family identified in this study. Members of this family are related to phosphorus solubilization (Chenet al., 2006) and are known to improve plant growth (Morales-Garcíaet al., 2011). In fact, almost half of the strains identi-fied in this study as phosphate solubilizers belonged to the Enterobacteriaceae family. In addition, strains belonging to the genus Enterobacter were found to produce higher amounts of indolic compounds (ICs) and to be widely asso-ciated both with roots and rhizospheric soils. The improved production of ICs by the Enterobactereacea group is sup-ported in Costaet al.(2014), in which a wider range of iso-lates related to various crops than wheat were evaluated.
As compared to the high number of isolates able to produce ICs and siderophores, a low number of strains was observed as able to solubilize phosphate. As the soils col-lected for this study were under regular regime of phos-phate fertilization, the present result is in accordance with Huet al.(2009), who found that P-solubilizing bacteria are more metabolically active in P-deficient fertilized soils af-ter long-term fertilizer management. Although the tricalcium phosphate solubilization assay has been used for several years, it does not recreate the complexity of phos-phorus in soil environments, that include common forms of phosphate that are more difficult to solubilize, such as Fe-P and Al-P (Bashanet al., 2013). Thus we must be aware that the isolates that were considered as phosphate solubilizers might be unable to solubilize all P sources in natural soils. The ability to produce siderophores, however, was frequently demonstrated by more than 60% of the bacterial isolates. On the other hand, without exception, all isolates were able to produce ICs; the majority of which did so in high amounts. The occurrence of these PGP traits is in ac-cordance with the literature (Guerinot, 1994; Spaepen et al., 2007; Hayatet al., 2010). Costaet al.(2013) found that the number of good IC producers increased when plants were subjected to light fertilization conditions as compared to zero and high fertilizer inputs. Considering this, although it was not evaluated in this study, the high number of IC producers was in agreement with the suitable nutrient con-dition (P, K, and organic matter content) of the soils regard-ing to the harborregard-ing crop (wheat). Indeed, phosphate solubilizer strains were identified more frequently in the rhizospheric soil samples; while siderophores producers were identified in larger number and more often closely re-lated to the roots, whereas IC producers were equally dis-tributed in roots and soil. These different occurrences of PGP traits between rhizospheric soil and root related to bacterial samples suggests that, in the case of IC producers, the bacterial samples present in these two niches are simi-larly influenced by the plant exudates. Such observations provide an idea of the plant’s needs. The vegetal system tends to apply selective pressure over the microbial com-munity, in order to enrich the vicinity of plant roots with beneficial bacteria, through the modulation of root exudates (Denniset al., 2010). Even though siderophores might be able to migrate, the present results enable us to in-fer that there is a prein-ference on siderophores production, which comes along with ICs production, rather than phos-phate solubilizers in the agricultural wheat crops in this study.
Although the analyzed plants were under the same fertilization regime, it was possible to visualize differences in the distribution and structure of PGPB communities as-sociated with wheat. MCA and CatPCA were useful to es-tablish visual relationships for a large set of data; most of them confirmed through chi-square and adjusted residual analysis. Most of the relationships between the sampling
sites in the results of PCA were supported by the results of MCA (Figure 3). Distribution of genera according to their frequency and homogeneity strongly link the JC and SB lo-calities, in other words, these sampling sites might share similar communities. CM and GM localities, despite their geographical proximity, presented some differences in their bacterial communities; whereas the VA and BV local-ities remained separate from the other locallocal-ities and from each other, possibly due to the same distinctive parameters inferred by PCA analysis: altitude (not evaluated) and or-ganic matter content. Thus, the shifts in diazotrophic com-munities could not be linked to the direct geographic distance between sites, such as reported by Wakelinet al. (2010).
PGP traits were also associated to locality; however, the analysis of these associations was non-conclusive. For example, the bacterial community from the VA locality lacks phosphate solubilization activity although this site also lacks phosphorus content in soil. As a low number of phosphate solubilizers were found, it is correct to infer that organisms other than the cultivable diazotrophic commu-nity, such as mycorrhizal fungi, could supply this particular nutrient. The bacterial samples from BV and CM localities contain the best siderophores producers (strongly associ-ated with the production of level 3); that from GM locality is associated with non-siderophores production. The bacte-rial samples from JC, SB and VA localities did not present a high number of siderophores producers, and in those from SB and VA localities we found a moderate number of siderophores producers, clearly expressing diversity among localities.
MCA and CatPCA became a useful tool to identify the relationships between bacterial diversity, functional traits of plant growth promotion and sampling sites, and be-came a starting point to pursue the objectives of the study. In particular, they showed a clear separation of bacterial strains belonging to certain genera that were able to solubilize nutrients from those able to produce ICs in this system. This information combined to what can be inferred about the plant’s needs (i.e.siderophores production prefer-entially expressed close to the roots, phosphate solubili-zation in the rhizosphere, and indolic compound production in both) in the present conditions, may help further studies to drive the prospection of PGPB regarding the desired or-ganism and PGP trait.
Acknowledgments
References
Ambrosini A, Beneduzi A, Stefanski T, Pinheiro FG, Vargas LK and Passaglia LMP (2012) Screening of plant growth pro-moting rhizobacteria isolated from sunflower (Helianthus annuusL.). Plant Soil 356:245-264.
Baldani V, Baldani JI and Döbereiner J (1983) Effects of
Azospirilluminoculation on root infection and nitrogen in-corporation in wheat. Can J Microbiol 29:924-929. Bashan Y, Kamnev AA and de-Bashan LE (2013) Tricalcium
phosphate is inappropriate as a universal selection factor for isolating and testing phosphate-solubilizing bacteria that en-hance plant growth: a proposal for an alternative procedure. Biol Fertil Soils 49:465-479.
Bulgarelli D, Rott M, Schlaeppi K, van Themaat EVL, Ahma-dinejad N, Assenza F, Rauf P, Huettel B, Reinhardt R, Schmelzer E,et al.(2012) Revealing structure and assembly cues for Arabidopsisroot-inhabiting bacterial microbiota. Nature 488:91-95.
Chen W-M, de Faria SM, Straliotto R, Pitard RM, Simões-Araújo JL, Chou Y-J, Chou J-H, Barrios E, Prescott AR, Elliott GN,
et al.(2005) Proof thatBurkholderiaforms effective symbi-oses with legumes: a study of novel Mimosa-nodulating strains from South America. Appl Environ Microbiol 71:7461-7471.
Chen YP, Rekha PD, Arun AB, Shen FT, Lai W-A and Young CC (2006) Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl Soil Ecol 34:33–41.
Coenye T and Vandamme P (2003) Diversity and significance of
Burkholderiaspecies occupying diverse ecological niches. Environ Microbiol 5:719–729.
Compant S, Nowak J, Coenye T, Clément C and Barka EA (2008) Diversity and occurrence ofBurkholderiaspp. in the natural environment. FEMS Microbiol Rev 32:607-626.
Costa PB, Beneduzi A, Souza R, Schoenfeld R, Vargas LK and Passaglia LMP (2013) The effects of different fertilization conditions on bacterial plant growth promoting traits: guide-lines for directed bacterial prospection and testing. Plant Soil 368:267-280.
Costa PB, Granada CE, Ambrosini A, Moreira F, de Souza R, Passos JFM, Arruda L and Passaglia LMP (2014) A Model to explain plant growth promotion traits: a multivariate anal-ysis of 2,211 bacterial isolates. PLoS ONE 9:e116020. Dennis PG, Miller AJ and Hirsch PR (2010) Are root exudates
more important than other sources of rhizodeposits in struc-turing rhizosphere bacterial communities? FEMS Microbiol Ecol 72:313-327.
Döbereiner J (1988) Isolation and identification of root associated diazotrophs. Plant Soil 110:207-212.
Felske A, Rheims H, Wokerink A, Stackebrandt E and Akkermans DL (1997) Ribosome analysis reveals promi-nent activity of an uncultured member of the class Actino-bacteria in grasslands soils. Microbiology 143:2983-2989. Fierer N and Jackson RB (2006) The diversity and biogeography
of soil bacterial communities. Proc Natl Acad Sci U S A 103:626-631.
Germida JJ and Siciliano SD (2001) Taxonomic diversity of bac-teria associated with the roots of modern, recent and ancient wheat cultivars. Biol Fertil Soils 33:410–415.
Glickmann E and Dessaux Y (1995) A critical examination of the specificity of the Salkowskir eagent for indolic compounds
produced by phytopathogenic bacteria. Appl Environ Microbiol 61:793–796.
Guerinot ML (1994) Microbial iron transport. Annu Rev Micro-biol 48:743-772.
Gyaneshwar P, Hirsch AM, Moulin L, Chen WM, Elliott GN, Bontemps C, Estrada-de Los Santos P, Gross E, dos Reis FB, Sprent JI,et al.(2011) Legume-nodulating betaproteo-bacteria: diversity, host range, and future prospects. Mol Plant Microbe Interact 24:1276-1288.
Hartmann A, Schmid M, van Tuinen D and Berg G (2009) Plant-driven selection of microbes. Plant Soil 321:235–257. Hayat R, Ali S, Amara U, Khalid R and Ahmed I (2010) Soil
bene-ficial bacteria and their role in plant growth promotion: a re-view. Ann Microbiol 60:579–598.
Helmke PA and Sparks DL (1996) Lithium, sodium, potassium, rubidium, and cesium. In: Sparks DL (ed) Methods of soil analysis. Part 3. Chemical methods. SSSA Book Series No. 5. SSSA and ASA, Madison, WI. pp 551-574.
Hoffman DL (1992) Interpreting Multiple Correspondence Anal-ysis as a multidimensional scaling method. Market Lett 3:259-272.
Hu J, Lin X, Wang J, Chu H, Yin R and Zhang J (2009) Population size and specific potential of P-mineralizing and -solubi-lizing bacteria under long-term P-deficiency fertilization in a sandy loam soil. Pedobiologia 53:49-58.
James EK, Gyaneshwar P, Mathan N, Barraquio QL, Reddy PM, Iannetta PPM, Olivares FL and Ladha JK (2002) Infection and colonization of rice seedlings by the plant growth-promoting bacteriumHerbaspirillum seropedicaeZ67. Mol Plant Microbe Interact 15:894-906.
Kennedy IR, Choudhury ATMA and Kecske’s ML (2004) Non-symbiotic bacterial diazotrophs in crop-farming systems: can their potential for plant growth promotion be better ex-ploited? Soil Biol Biochem 36:1229-1244.
Kuo S (1996) Phosphorus. In: Sparks DL (ed) Methods of soil analysis. Part 3. Chemical methods. Soil Sci. Soc. Am. Book Series: 5. Soil Sci. Soc. Am. Madison, WI, pp 8669–920. Lugtenberg B and Kamilova F (2009) Plant-Growth-Promoting
Rhizobacteria. Annu. Rev Microbiol 63:541-556.
Lundberg DS, Yourstone S, Mieczkowski P, Jones CD and Dangl JL (2013) Practical innovations for high-throughput amplicon sequencing. Nat Methods 10:999-1002.
Mirza MS, Mehnaz S, Normand PM, Prigent-Combaret C, Moënne-Loccoz Y, Bally R and Malik KA (2006) Molecu-lar characterization and PCR detection of a nitrogen fixing
Pseudomonasstrain promoting rice growth. Biol Fertil Soils 43:163-170.
Morales-García YE, Juárez-Hernández D, Aragón-Hernández C, Mascarua-Esparza MA, Bustillos-Cristales MR, Fuentes-Ramírez LE, Martínez-Contreras RD and Muñoz-Rojas J (2011) Growth response of maize plantlets inoculated with
Enterobacter spp., as a model for alternative agriculture. Rev Argent Microbiol 43:287-293.
Mulvaney RL (1996) Nitrogen – inorganic forms. In: Sparks DL (ed.) Methods of soil analysis. Part 3. Chemical methods. SSSA Book Series No. 5. SSSA and ASA, Madison, WI, pp 1123-1184.
Palmer KM and Young JPW (2000) Higher diversity of
Rhizobium leguminosarumbiovarviciaepopulations in ara-ble soils than in grass soils. Appl Environ Microbiol 66:2445-2450.
Perin L, Martínez-Aquilar L, Castro-González R, Estrada-delos Santos P, Cabbellos-Avelar T, Guedes HV, Reis VM and Caballero-Mellado J (2006) DiazotrophicBurkholderia spe-cies associated with field-grown maize and sugarcane. Appl Environ Microbiol 72:3103-3110.
Ramette A and Tiedje JM (2007) Multiscale responses of micro-bial life to spatial distance and environmental heterogeneity in a patchy ecosystem. Proc Natl Acad Sci U S A 104:2761-2766.
Sambrook J and Russel DW (2001) Molecular cloning: a labora-tory manual. Cold Spring Harbor Laboralabora-tory Press, NewYork.
Schwyn B and Neilands JB (1987) Universal chemical assay for the detection and determination of siderophores. Anal Biochem 160:47-56.
Shannon CE and Weaver W (1949) The mathematical theory of communication. University of Illinois Press, Urbana,19 p. Spaepen S, Vanderleyden J and Remans R (2007) Indole-3-acetic
acid in microbial and microorganism-plant signaling. FEMS Microbiol Rev 31:425-448.
Tilman D (2001) Functional diversity. In: Levin SA (ed) Encyclo-pedia of Biodiversity.Elsevier, Oxford, pp 109-120. Unno Y, Okubo K, Wasaki TS and Osaki M (2005) Plant growth
promotion abilities and microscale bacterial dynamics in the
rhizosphere of lipin analyzed by phytate utilization ability. Environ Microbiol 7:396-404.
Wakelin SA, Gupta VVSR and Forrester ST (2010) Regional and local factors affecting diversity, abundance, and activity of free-living, N2-fixing bacteria in Australian agricultural soils. Pedobiology 53:391-399.
Wartiainen I, Eriksson T, Zheng W and Rasmussen U (2008) Variation in the active diazotrophic community in rice paddy:nifHPCR-DGGE analysis of rhizosphere and bulk soil. Appl Soil Ecol 39:65-75.
Internet Resources
EzTaxon database, http://eztaxon-e.ezbiocloud.net/ (accessed on October, 10th, 2015)
Supplementary Material
The following online material is available for this ar-ticle:
Table S1 – Input dataset for MCA analysis. Table S2 – Input dataset for CatPCA analysis. This material is available as part of the online article from http://www.scielo.br/gmb.
Associate Editor: Célia Maria de Almeida Soares
![Figure 1 - Principal Component Analysis (PCA) of diversity indices in re- re-lation to soil abiotic properties [clay, organic matter content (OMC), pH, P, K,] determined from the six sampling sites (JC – Júlio de Castilhos, SB – São Borja, VA – Vacaria, CM](https://thumb-eu.123doks.com/thumbv2/123dok_br/16056114.696290/3.918.496.816.661.958/principal-component-analysis-diversity-properties-determined-sampling-castilhos.webp)