Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Animal
Fitness dos híbridos entre duas espécies de ratos fossadores, Microtus
lusitanicus (Gerbe, 1879) e M. duodecimcostatus (de Selys-Longchamps,
1839) - uma abordagem multidisciplinar
Joana Margarida Afonso Soares
Dissertação
Mestrado em Biologia Humana e Ambiente
Universidade de Lisboa
Faculdade de Ciências
Departamento de Biologia Animal
Fitness dos híbridos entre duas espécies de ratos fossadores, Microtus
lusitanicus (Gerbe, 1879) e M. duodecimcostatus (de Selys-Longchamps,
1839) - uma abordagem multidisciplinar
Joana Margarida Afonso Soares Dissertação
Mestrado em Biologia Humana e Ambiente
Dissertação orientada por Prof.ª Doutora Maria da Luz Mathias
Doutora Ana Mota Cerveira
Todas as informações efectuadas no presente documento são de exclusiva responsabilidade do seu autor, não cabendo qualquer responsabilidade à Faculdade de Ciências da Universidade de Lisboa pelos conteúdos nele apresentados.
Agradecimentos
Em primeiro lugar, à minha orientadora professora Maria da Luz Mathias por me ter dado a oportunidade de dissertar num tema que mais se adequa ao meu interesse, bem como o apoio ao longo da execução do mesmo.
Um agradecimento muito especial à Ana Cerveira por me ter orientado ao longo do meu trabalho, pela amizade e apoio incondicional.
À Rita Monarca, Joaquim Tapisso, Sofia Gabriel e Gabriel Martins pela colaboração e apoio laboratorial.
4
Índice
Inserção no Mestrado de BHA ... 6
Resumo ... 7 Abstract ... 8 Índice de figuras ... 9 Índice de tabelas ... 10 Índice de anexos ... 11 1. Introdução ... 12
1.1. Mamíferos subterrâneos e fossadores... 12
1.2. Roedores fossadores em Portugal ... 13
1.3. Hibridação ... 15
1.4. Objectivos ... 17
2. Enquadramento e planeamento ... 18
2.1. Crescimento e Fertilidade ... 18
2.2. Sistemas de reconhecimento de parceiro sexual ... 20
2.3. Metabolismo ... 21
3. Material e métodos ... 23
3.1. Manutenção dos animais ... 23
3.2. Crescimento e fertilidade ... 24
3.2.1. Análise estatística ... 24
3.3. Análise dos Espermatozóides ... 25
3.3.1. Procedimento experimental ... 25
3.3.2. Análise estatística ... 27
3.4. Testes de preferência sexual ... 28
3.4.1. Procedimento experimental ... 28
3.4.2. Análise de dados ... 30
5 3.5. Metabolismo Basal ... 31 3.5.1. Procedimento experimental ... 31 3.5.2. Análise de dados ... 33 3.5.3. Análise estatística ... 33 4. Resultados ... 35 4.1. Crescimento e fertilidade ... 35
4.2. Análise dos Espermatozóides ... 39
4.2.1. Medições corporais ... 39
4.2.2. Dimensões do esperma ... 40
4.2.3. Velocidade do esperma ... 47
4.3. Testes de preferência sexual ... 47
4.3.1. Padrões de preferência sexual entre uma fêmea conspecífica e uma fêmea híbrida 47 4.4. Metabolismo Basal ... 51
5. Discussão ... 53
6. Conclusão ... 61
7. Referências bibliográficas ... 63
6
Inserção no Mestrado de BHA
O rato-toupeira (Microtus lusitanicus) e o rato-toupeira mediterrânico (M. duodecimcostatus) são roedores fossadores frequentemente considerados espécies praga nos sistemas agrícolas causando enormes prejuízos financeiros. A recente descoberta de híbridos das duas espécies na natureza, torna o estudo destes indivíduos especialmente relevante, tendo em conta que por vezes os híbridos apresentam um fitness superior ao das espécies parentais (vigor híbrido), o qual, caso se verifique, poderá representar um grave problema em termos ambientais. O estudo do fitness dos híbridos destas duas espécies animais revela-se assim de grande importância, de modo a poder avaliar a sua viabilidade e impacto na natureza.
Para além da importância ambiental do estudo destas espécies, as técnicas realizadas ao longo de todo o trabalho com os ratos-toupeira e seus híbridos podem ser usadas em inúmeros estudos de biologia, noutros modelos e explorando outros objectivos. As áreas desenvolvidas nesta tese são o desenvolvimento, a fertilidade, o metabolismo e o comportamento.
7
Resumo
Os ratos-toupeira Microtus lusitanicus (Gerbe, 1879) e M. duodecimcostatus (de Selys-Longchamps, 1839) são espécies irmãs fossadoras passando grande parte das suas vidas no subsolo. O início da sua divergência evolutiva data de 1,2 a 2 milhões de anos. Em termos evolutivos, o isolamento reprodutor entre elas ainda não está totalmente estabelecido, tendo sido encontrados híbridos recentes numa zona onde as duas espécies coexistem. Neste estudo foi feita uma análise multidisciplinar do fitness dos híbridos, considerando em particular a taxa de crescimento, a fertilidade, a preferência sexual e a taxa de metabolismo em repouso (TMR). A análise das curvas de crescimento dos híbridos mostrou que estes apresentam taxas de crescimento intermédias relativamente às espécies parentais. No entanto, os dados de fertilidade, a análise dos espermatozóides, da preferência sexual e do metabolismo basal revelaram que os híbridos terão provavelmente um fitness inferior na natureza, comparativamente aos seus parentais. Nos backcrosses verificou-se que as fêmeas híbridas têm fertilidade reduzida, e os machos híbridos são estéreis, não tendo sido encontrados espermatozóides à excepção de um único indivíduo. Em geral, os machos das duas espécies parentais apresentaram uma preferência sexual pelas fêmeas conspecíficas relativamente às fêmeas híbridas. Relativamente às taxas de metabolismo em repouso observou-se que os híbridos apresentam geralmente TMRs mais elevadas do que as espécies parentais. Em conjunto, os resultados obtidos sugerem que a ocorrência de híbridos entre M. lusitanicus e M. duodecimcostatus será um fenómeno cada vez mais raro na natureza, sendo provavelmente estabelecido o isolamento reprodutor entre as duas espécies.
Palavras-chave: micromamíferos; rato-toupeira; rato-toupeira mediterrânico; hibridação; metabolismo; preferência sexual; fertilidade.
8
Abstract
The pine voles Microtus lusitanicus (Gerbe, 1879) and M. duodecimcostatus (de Selys-Longchamps, 1839) are fossorial sister species spending most of the time underground. Their evolutionary divergence began about 1.2 to 2 million years ago. Reproductive isolation between the two species hasn’t, however, been fully established, recent hybrids having been found in a region where both species coexist. In this study, a multidisciplinary approach of hybrid fitness was made, in particular the growth rate, the fertility, mate choice and resting metabolic rate (RMR) were analyzed. The analysis of hybrid growth rate revealed that these animals show intermediate growth patterns relatively to the parental species. However, the results obtained from fertility, sperm analysis, mate choice, and basal metabolism suggest that the hybrids should have a lower fitness than their parental species under natural conditions. Results from backcrosses show that female hybrids have a reduced fertility, and male hybrids are sterile, no sperm having been found with the exception of a single individual. In general, males from both parental species showed a preference for conspecific females relatively to hybrid females. Concerning the resting metabolic rate, hybrids generally showed a higher RMR than parental species. Altogether, the results obtained suggest that the occurrence of hybrids between M. lusitanicus e M. duodecimcostatus in nature will become a rarer phenomenon with time, and reproductive isolation will probably be established.
Keywords: small mammals; Lusitanian pine vole; Mediterranean pine vole; hybridization; metabolism; mate choice; fertility.
9
Índice de figuras
Figura 1. Microtus duodecimcostatus (esquerda) e M. lusitanicus (direita) ... 13
Figura 2. Distribuição geográfica de Microtus lusitanicus e M. duodecimcostatus ... 14
Figura 3. Híbridos resultantes do cruzamento de Microtus duodecimcostatus e M. lusitanicus. Classe MDL (♀ M. duodecimcostatus x ♂ M. lusitanicus) (esquerda) e classe MLD (♀ M. lusitanicus x ♂ M. duodecimcostatus) (direita). ... 23
Figura 4. Olfactómetro ... 29
Figura 5. Sistema de respirometria de fluxo aberto ... 32
Figura 6. Peso das duas espécies de rato-toupeira, Microtus lusitanicus (ML), M. duodecimcostatus (MD), (Cerveira et al., em preparação) e seus híbridos MLD (♀ M. lusitanicus x ♂ M. duodecimcostatus) e MDL (♀ M. duodecimcostatus x ♂ M. lusitanicus) relativamente à sua idade. Curvas de crescimento para cada grupo obtidas através da aplicação do modelo de Gompertz. ... 36
Figura 7. Dispersão do logaritmo das médias da massa dos testículos relativamente ao logaritmo das médias da massa corporal de Microtus duodecimcostatus (MD), M. lusitanicus (ML) e das duas classes de híbridos (MDL e MLD) ... 40
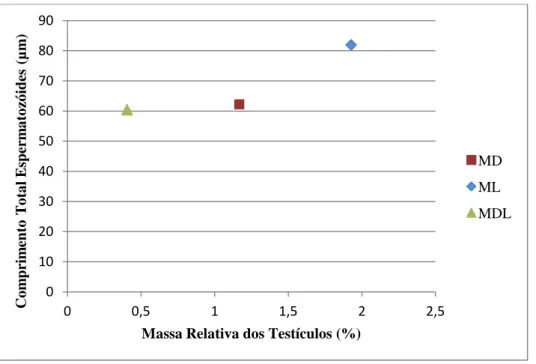
Figura 8. Relação entre a massa relativa dos testículos e o comprimento total dos espermatozóides de híbridos MDL e das espécies Microtus duodecimcostatus (MD) e M. lusitanicus (ML). ... 42
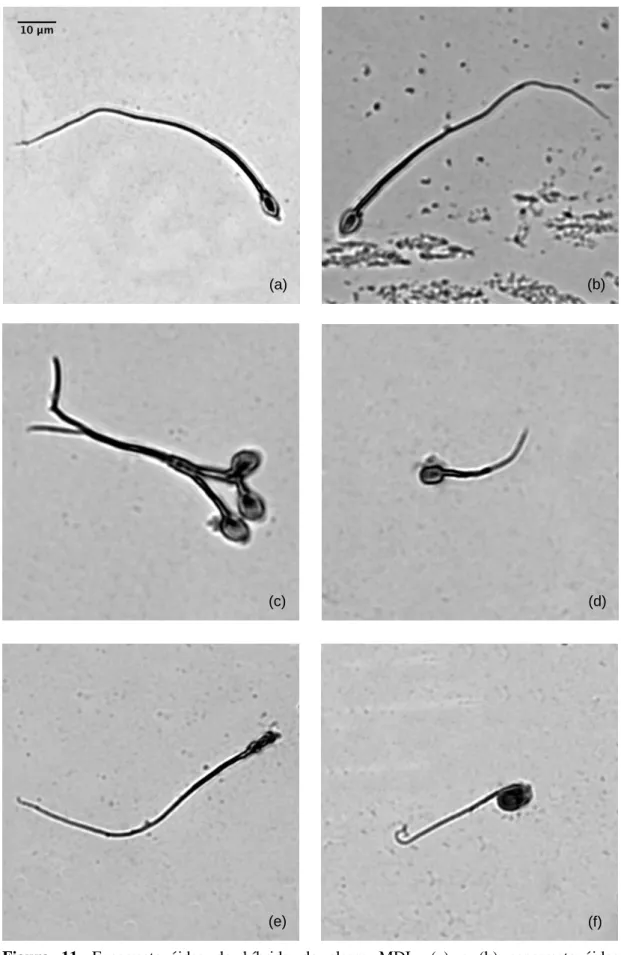
Figura 9. Espermatozóides da espécie Microtus lusitanicus ... 44
Figura 10. Espermatozóides da espécie Microtus duodecimcostatus... 45
Figura 11. Espermatozóides do híbrido da classe MDL ... 46
Figura 12. Tempo médio que os machos M. duodecimcostatus (MD) passaram em cada caixa estímulo ... 48
Figura 13. Tempo médio que os machos M. lusitanicus (ML) passaram em cada caixa estímulo . 49 Figura 14. Tempo médio que os machos M. duodecimcostatus (MD) estiveram em contacto directo com o estímulo ... 50
Figura 15. Tempo médio que os machos M. lusitanicus (ML) estiveram em contacto directo com o estímulo ... 50
10
Índice de tabelas
Tabela 1. Parâmetros de fertilidade dos backcrosses das duas espécies de rato-toupeira, Microtus
duodecimcostatus (MD) e M. lusitanicus (ML) ... 38
Tabela 2. Coeficientes de variância (CV), médias das medições corporais e massa relativa dos
testículos (MRT) dos grupos de teste, Microtus duodecimcostatus (MD), M. lusitanicus (ML), e das duas classes de híbridos (MDL e MLD) ... 39
Tabela 3. Média e coeficiente de variância (CV) das dimensões das várias estruturas dos
espermatozóides do híbrido da classe MDL e das espécies parentais Microtus duodecimcostatus (MD) e M. lusitanicus (ML) ... 41
Tabela 4. Valores médios medidos e valores médios estimados (ANCOVA) do volume de oxigénio
(VO2) consumido pelas duas classes de híbridos (MDL e MLD) às temperaturas de 5ºC, 20ºC e
11
Índice de anexos
Anexo I - Protocolo de análise dos espermatozóides ... 69 Anexo II - Hepes-buffered modified Tyrode's medium ... 70 Anexo III - Solução de Eosina-Nigrosina ... 71
12
1. Introdução
1.1. Mamíferos subterrâneos e fossadores
A evolução convergente das formas de vida subterrâneas é um fenómeno fascinante (Nevo, 1999). Estas formas de vida desenvolveram-se em muitas ordens de mamíferos, existindo actualmente cerca de 250 espécies de roedores que passam grande parte das suas vidas em galerias subterrâneas (Begal et al., 2007). O grau de dependência do meio subterrâneo dos roedores que habitam o subsolo é variável, formando um continuum entre as chamadas “espécies estritamente subterrâneas”, passando pelas “espécies subterrâneas facultativas”, até às “espécies fossadoras”. Enquanto as primeiras realizam a grande maioria das suas actividades (ex: acasalamento, reprodução e alimentação) no subsolo, raramente vindo à superfície (McNab, 1966), as últimas, à semelhança das espécies estritamente subterrâneas, também constroem os seus abrigos no subsolo, mas alimentam-se predominantemente à superfície (Burda et al., 2007).
O ecótipo subterrâneo é considerado estável e previsível a nível do microclima, em particular da temperatura (Nevo, 1999), funcionando como um tampão relativamente a flutuações e extremos ambientais externos predominantes à superfície do solo. Adicionalmente, os abrigos subterrâneos conferem aos seus habitantes protecção dos predadores, particularmente importante nos períodos de maior vulnerabilidade, nomeadamente, durante os períodos de repouso, hibernação, estivação e reprodução (Nevo, 1999). No entanto, apesar das vantagens oferecidas pelo modo de vida subterrâneo, este microclima apresenta alguns desafios à sobrevivência destes organismos, em particular a ausência de luz, uma circulação de ar muito limitada, níveis de humidade elevados, elevada concentração de dióxido de carbono (hipercapnia) e baixa concentração de oxigénio (hipoxia). Para além de condições microclimáticas adversas, os roedores subterrâneos têm ainda que se confrontar com os elevados custos energéticos inerentes à actividade subterrânea, nomeadamente a construção, manutenção e expansão dos seus abrigos através da escavação (Ebensperger & Bozinovic, 2000).
As condições únicas encontradas no meio subterrâneo originaram o desenvolvimento nos mamíferos subterrâneos de diversas adaptações quer ao nível da sua morfologia como também da sua fisiologia (Nevo, 1999). Em geral, estes mamíferos
13 apresentam corpos compactos, pescoço curto, cauda rudimentar, olhos reduzidos, patas dianteiras robustas, cintura escapular e músculos associados bem desenvolvidos, assim como uma baixa taxa de metabolismo basal (TMB), baixa temperatura corporal e uma elevada condutância térmica (Nevo, 1999). Apesar dos mamíferos fossadores também apresentarem adaptações ao meio subterrâneo, estes são em geral menos especializados quer em termos morfológicos quer em termos fisiológicos, quando comparados com os organismos estritamente subterrâneos (Biknevicius, 1993).
1.2. Roedores fossadores em Portugal
A classificação taxonómica do género Microtus é complexa e controversa devido à rapidez com que evoluiu e, muitas vezes, à variação gradual das características morfológicas e moleculares entre as 65 espécies que constituem o género (Mitchell-Jones et al., 1999). O rato-toupeira e o rato-toupeira mediterrânico, Microtus lusitanicus (Gerbe, 1879) e M. duodecimcostatus (de Selys-Longchamps, 1839) respectivamente, são consideradas espécies distintas devido às diferenças na sua morfologia e ecologia (Mitchell-Jones et al., 1999). A sua divergência iniciou-se há aproximadamente 1,2 a 2 milhões de anos a partir de um ancestral comum (Jaarola et al., 2004).
14 Ambas as espécies são endemismos europeus tendo um padrão de distribuição geográfico principalmente alopátrico, M. lusitanicus ocorrendo no Noroeste da Península Ibérica, incluindo os Pirenéus Franceses (Mira & Mathias, 2007), e M. duodecimcostatus no Centro-Sul da Península Ibérica e Sul de França (Cotilla & Palomo, 2007). Em Portugal, M. lusitanicus encontra-se distribuído a Norte do país e M. duodecimcostatus a Sul, existindo apenas uma área estreita de simpatria no centro do país entre Porto Covo e Castelo Branco (Madureira, 1984; Santos et al., 2010) (Fig. 2).
A distinção morfológica das duas espécies na natureza é por vezes dificultada pela sua enorme semelhança (Vinhas, 1993), embora M. lusitanicus apresente dimensões ligeiramente inferiores (comprimento: cabeça + corpo = 77,5-105mm; peso: 14-19g), relativamente a M. duodecimcostatus (comprimento: cabeça + corpo = 89-122mm; peso: 19-32g) (Madureira, 1984; Vinhas, 1993; Mathias et al., 1999).
Figura 2. Distribuição geográfica de Microtus lusitanicus e M. duodecimcostatus (modificada de
Santos, 2009).
Ambas as espécies possuem hábitos fossadores, construindo complexos sistemas de galerias subterrâneas nas quais realizam a maioria das suas actividades, alimentando-se no entanto maioritariamente à superfície. O rato-toupeira ocupa preferencialmente áreas com solos férteis, bem drenados, ácidos e pouco compactos, temperaturas mais baixas e com
15 maiores níveis de precipitação em relação ao rato-toupeira mediterrânico. Este último ocorre em solos argilosos e áreas de pastagem, temperaturas mais elevadas e menor precipitação (Santos et al., 2009). Ambas as espécies preferem habitats abertos que possam ser influenciados pela agricultura (Cotilla & Palomo, 2007; Mira & Mathias, 2007) ou com grande cobertura vegetal, a qual lhes fornece protecção dos predadores e alimento (Madureira, 1984), preferencialmente raízes, tubérculos, frutos e outras partes das plantas (Cotilla & Palomo, 2007; Mira & Mathias, 2007).
A profundidade das galerias parece estar relacionada com as propriedades do solo, nomeadamente a textura, a humidade, a temperatura, o tipo e a densidade da vegetação (Sezinando, 1982; Madureira, 1984). O rato-toupeira parece no entanto ter uma actividade mais superficial, construindo as suas galerias em menor profundidade, chegando mesmo no Inverno a construir ninhos à superfície e a utilizar caminhos na cobertura vegetal. Pelo contrário, o rato-toupeira mediterrânico parece ocorrer a uma maior profundidade apresentando um estilo de vida mais dependente do meio subterrâneo ao longo do ano (Vinhas, 1993; Paradis, 1995; Santos et al., 2009). Segundo Madureira (1984) estas diferenças poderão, por um lado, dever-se aos diferentes tipos de solo em que as duas espécies ocorrem: solos mais macios e com maior teor de água na área de ocorrência M. lusitanicus, e solos mais duros e secos na área de distribuição de M. duodecimcostatus, já que em solos com alto teor de humidade existe uma maior dificuldade na difusão dos gases levando a que os animais se desloquem mais à superfície (Mathias, 1990). Por outro lado, as temperaturas por vezes extremamente elevadas que caracterizam a área de ocorrência de M. duodecimcostatus (i.e., sul do país), podem ter levado a uma existência mais dependente do ecótipo subterrâneo por parte desta espécie relativamente a M. lusitanicus.
1.3. Hibridação
Designa-se por hibridação a produção de descendência a partir do cruzamento de indivíduos de duas populações geneticamente diferentes (Harrison, 1990). Existem duas formas de hibridação, nomeadamente hibridação primária e hibridação secundária. A primeira acontece in situ através da selecção natural alterando as frequências alélicas em duas ou mais populações que se distribuem continuamente, normalmente devido a grandes alterações de um ou mais factores ambientais no local. A segunda dá-se quando duas
16 populações que divergiram geneticamente contactam secundariamente e acasalam entre si (Endler, 1977).
Nos animais, a hibridação é considerada um fenómeno relativamente raro na natureza (cerca de 10% das espécies animais; Mallet, 2005), sendo no entanto aceite como um fenómeno generalizado (Hochkirch & Lemke, 2011) e com interesse em termos evolutivos, visto que em determinados casos pode dar origem a novas adaptações ou mesmo novas espécies (Arnold, 1997; Mallet, 2007).
Apesar do processo de especiação de M. duodecimcostatus e M. lusitanicus se ter iniciado há cerca de dois milhões de anos, o isolamento reprodutor entre estes dois taxa não parece estar ainda totalmente estabelecido, tendo um estudo recente revelado a ocorrência de animais híbridos na zona de simpatria destas espécies (Bastos-Silveira et al., 2012). Este fenómeno indica que no caso dos ratos-toupeira parece verificar-se um processo de hibridação secundária na zona de simpatria, visto que as duas espécies acasalam devido a um segundo contacto desde o início da sua suposta divergência evolutiva.
Em zonas de hibridação secundária, os híbridos têm frequentemente um baixo fitness intrínseco devido a desvantagem heterogâmica e incompatibilidade epistática num determinado loci (Barton & Gale, 1993). Estas razões parecem estar envolvidas no cruzamento de duas subespécies Europeias de ratinho caseiro (Mus musculus domesticus e M. m. musculus) em que os machos são estéreis e as fêmeas férteis (Britton-Davidian et al., 2005). No entanto, um trabalho recente demonstra que o cruzamento de duas espécies de gaivota (Larus glaucescens e L. occidentalis) numa zona de hibridação secundária produz híbridos com um maior fitness relativamente às espécies parentais (Good et al., 2000).
A ocorrência de híbridos de M. duodecimcostatus e M. lusitanicus na natureza apresenta três possíveis cenários dependendo do seu fitness (i.e., capacidade de produzir descendência) relativamente ao fitness das espécies parentais. Assim, os híbridos podem apresentar: a) um fitness inferior que, através da selecção natural, irá favorecer alelos que estabeleçam o isolamento reprodutor entre as duas espécies (ex.: papa-moscas Ficedula hypoleuca e F. albicollis; Veen et al., 2001) e eventualmente impedir o fluxo genético e a formação de híbridos; b) um fitness superior, ou vigor híbrido, resultando no aumento da frequência dos alelos que favorecem o fitness dos híbridos e, em caso extremo, podendo
17 quebrar a barreira pós-zigótica de fluxo genético entre as duas espécies e dar origem a uma única espécie (ex.: salamandras Ambystoma californiense e A. tigrinum mavortium; Fitzpatrick & Shaffer, 2007); c) um fitness igual, podendo tornar-se isolados reprodutivamente das formas parentais e formar uma terceira espécie (ex.: relas Hyla cinerea e H. gratiosa; Lamb et al., 1990) (para mais exemplos ver Arnold & Hodges, 1995).
Assim sendo, o estudo de populações simpátricas das duas espécies e dos seus híbridos reveste-se de especial interesse para a compreensão dos processos e mecanismos de divergência evolutiva destas espécies.
1.4. Objectivos
O principal objectivo deste trabalho é estudar o fitness dos híbridos entre duas espécies de ratos-toupeira, M. duodecimcostatus e M. lusitanicus, considerando que o fitness respeita não só o sucesso reprodutivo de um indivíduo, mas também algumas das suas características intrínsecas e, consequentemente, a capacidade de sobrevivência no seu meio natural.
Para atingir estes objectivos será utilizada uma abordagem multidisciplinar integrada que inclui o estudo de diversos parâmetros, nomeadamente o crescimento, a fertilidade, a preferência sexual e o metabolismo em repouso de híbridos produzidos em cativeiro. O cumprimento destes objectivos irá permitir esclarecer:
a) o papel das barreiras de isolamento reprodutor no cruzamento entre M. duodecimcostatus e M. lusitanicus;
b) a viabilidade (taxas de crescimento) e fertilidade (número de ninhadas e crias) dos híbridos entre M. duodecimcostatus e M. lusitanicus;
c) a influência das características dos espermatozóides na fertilidade;
d) o papel de estímulos químicos no reconhecimento de parceiros sexuais conspecíficos e híbridos;
18
2. Enquadramento e planeamento
Os mecanismos através dos quais as espécies se formam e mantêm a sua integridade envolvem muitas vezes o desenvolvimento de barreiras de isolamento reprodutor que impedem o fluxo de genes entre as populações (Coyne & Orr, 2004). Um dos objectivos deste trabalho é investigar algumas destas de barreiras e determinar se estão a actuar na manutenção da integridade das duas espécies de ratos-toupeira quando em simpatria, reduzindo quer a probabilidade de cruzamentos interespecíficos, como o fitness dos híbridos.
As barreiras de isolamento reprodutor podem ser classificadas em:
a) barreiras pré-acasalamento que impedem o fluxo de genes devido a isolamento ecológico, mecânico ou comportamental;
b) barreiras pré-zigóticas que previnem o sucesso da formação de zigotos híbridos mesmo depois do acasalamento, devido a uma falha na união dos gâmetas;
c) barreiras pós-zigóticas, que consistem na inviabilidade (taxas de sobrevivência baixas), fertilidade reduzida ou esterilidade dos híbridos devido a diferenças estruturais entre cromossomas e diferenças entre genes (Coyne & Orr, 2004; Futuyma, 2005).
Devido à diferenciação da maior parte dos organismos entre os quais ocorre esterilidade dos híbridos, existem argumentos de que as barreiras pós-zigóticas se manifestam muito posteriormente à formação das barreiras pré-zigóticas, sendo estas a iniciar o processo de especiação (Mallet, 2006).
2.1. Crescimento e Fertilidade
A importância de estudos de crescimento e fertilidade deve-se à informação que estes dois factores transmitem acerca do papel das características genéticas na anatomia, fisiologia e padrão de comportamento nos adultos, facilitando a compreensão da diversidade filogenética dos organismos (Nadeau, 1985). Relativamente ao crescimento serão monitorizados pares de cruzamentos interespecíficos de modo a verificar a existência de novas ninhadas e acompanhar o desenvolvimento das crias já existentes, monitorizando
19 o seu peso corporal durante o crescimento. Estes dados serão depois comparados com os dados dos indivíduos das espécies parentais, de modo a avaliar possíveis alterações nas taxas de crescimento e fertilidade (número de crias e tamanho das ninhadas), por vezes observadas devido a uma desvantagem heterogâmica e incompatibilidade epistática num determinado loci (Barton & Gale, 1993), como referido anteriormente.
Quando os híbridos são viáveis podem ainda surgir outras limitações na descendência dos cruzamentos interespecíficos, como a ausência de um dos sexos, uma predominância de um deles ou mesmo a sua infertilidade. Este fenómeno, conhecido como a regra de Haldane, estabelece que quando um sexo é ausente, raro ou estéril na primeira geração resultante do cruzamento entre duas espécies, esse sexo é normalmente o sexo heterogâmico (Haldane, 1922). Quer o sexo heterogâmico seja o macho (mamíferos e maioria dos insectos) quer seja a fêmea (aves e lepidópteros), a origem da esterilidade híbrida é um fenómeno complexo estando ainda hoje em debate quais os mecanismos que levam à sua ocorrência (Wu et al., 1996). Sendo que as espécies M. duodecimcostatus e M. lusitanicus são mamíferos poderá se verificada infertilidade nos machos.
Nos mamíferos quando uma fêmea acasala com mais do que um macho ocorre competição de espermatozóides para fertilizar o oócito (Parker, 1970; Gomendio et al., 1998). Como resultado de adaptação, este acréscimo da competição de espermatozóides está associado a um aumento da velocidade de movimentação dos espermatozóides até ao oócito, que em determinados estudos tem demonstrado associações positivas com o tamanho do espermatozóide envolvendo um aumento de todas as características, tais como o alongamento da cabeça (Malo, 2006), uma zona intermédia mais volumosa ou mais longa suportando mais mitocôndrias e um flagelo maior (Gomendio & Roldan, 1991).
Relativamente à fertilidade realizar-se-á uma análise da qualidade das características dos espermatozóides dos machos híbridos no sentido de os comparar com os das espécies parentais. A qualidade das características dos espermatozóides e a sua influência na fertilidade geralmente inclui a proporção de espermatozóides móveis e viáveis, sem anomalias nas suas diferentes estruturas e com um acrossoma intacto (Florman & Ducibella, 2006). Se um espermatozóide possuir anomalias torna-se incapaz de fertilizar o oócito visto que a sua estrutura afecta a mobilidade (Pukazhenthi et al., 2006). Já a integridade do acrossoma é importante quando o espermatozóide atinge o oócito e precisa de penetrar a parede mucosa para o fertilizar (Smith & Yanagimachi,
20 1990). Uma grande proporção de espermatozóides morre durante o percurso no sistema reprodutivo da fêmea, por isso a viabilidade dos mesmos é essencial para o sucesso reprodutivo do macho (Scott, 2000). Assim, se os machos híbridos forem estéreis como diz a regra de Haldane será de esperar que os seus espermatozóides tenham mais anomalias do que os espermatozóides de indivíduos das espécies parentais, impedindo-os de produzir descendência.
2.2. Sistemas de reconhecimento de parceiro sexual
Os sistemas sensoriais e os sinais que estes recebem desempenham um papel particularmente relevante na especiação, actuando frequentemente como barreiras pré-zigóticas através dos sistemas de reconhecimento específico (i.e., reconhecimento de parceiro sexual) (Smadja & Butlin, 2009). No caso dos mamíferos, o olfacto é em geral o sentido dominante no reconhecimento de indivíduos conspecíficos, apesar de a informação adquirida através dos outros sentidos, tais como a visão e a audição, ser também essencial (Wyatt, 2003). Existe uma enorme variedade de moléculas libertadas para o ambiente por parte destes organismos, desde pequenas moléculas voláteis a complexas proteínas de grande dimensão. Estas substâncias podem ser libertadas através da urina, fezes, ou secreções da pele, tracto reprodutivo ou glândulas odoríferas especializadas, transmitindo informação sobre o sexo, a idade, saúde, estado reprodutivo, entre outras, representando o perfil completo do indivíduo que as produz (Brennan & Keverne, 2004). Assim, e tendo em conta que a preferência olfactiva pode funcionar como um mecanismo comportamental de isolamento pré-zigótico (D'Udine & Alleva, 1983), será importante determinar se no caso dos ratos-toupeira também existem evidências de reconhecimento conspecífico através de um estímulo químico, ou seja, se o isolamento reprodutor poderá ser significativamente influenciado pela capacidade de identificação química, já que estas espécies ocorrem em simpatria em parte da sua distribuição. Assim, o objectivo é verificar se as espécies parentais apresentam preferência na escolha de parceiro sexual entre conspecíficos e híbridos. Para tal será analisada a sua preferência entre conspecíficos e híbridos, utilizando a urina e os dejectos dos animais como estímulo químico. Espera-se que as espécies parentais tenham preferência pelos estímulos químicos dos seus conspecíficos. No entanto, tendo em conta que os híbridos resultam do cruzamento das duas espécies parentais, há também a possibilidade dos estímulos químicos dos híbridos
21 apresentarem características mais semelhantes com uma das espécies parentais levando a uma ausência de preferência.
2.3. Metabolismo
Uma das características fisiológicas de maior realce das espécies subterrâneas é a sua baixa taxa de metabolismo basal (TMB) quando comparada com a observada em espécies epígeas de dimensão corporal semelhante (McNab, 1966). A TMB representa o mínimo de energia que um indivíduo necessita para manter o funcionamento do corpo quando está em repouso a uma temperatura ambiental neutra para a termorregulação (Sedláček, 2007). Duas hipóteses não exclusivas têm sido sugeridas na tentativa de relacionar este fenómeno com as condições microambientais impostas pelo ecótipo subterrâneo, nomeadamente a “hipótese do stress térmico”, e a “hipótese dos custos de construção do abrigo”. Enquanto a primeira sugere que uma TMB baixa minimiza a possibilidade de sobreaquecimento no sistema de galerias onde a perda de calor por convecção e transpiração é substancialmente reduzida (McNab, 1966, 1979), a segunda hipótese propõe que uma TMB baixa poderá compensar o dispêndio de energia extremamente alto resultante da escavação das galerias, já que esta actividade pode representar um gasto energético 300 vezes superior àquele necessário para percorrer a mesma distância à superfície (Vleck, 1979, 1981). Num estudo comparativo considerando cerca de 100 espécies subterrâneas e fossadoras, ambas as hipóteses são verificadas: espécies subterrâneas de habitats áridos apresentando uma TMB inferior relativamente a espécies fossadoras de ambientes mésicos (hipótese do custo de escavação), e espécies subterrâneas e fossadoras de ambientes mésicos apresentando TMB semelhantes (hipótese do stress térmico). As duas hipóteses não são no entanto mutuamente exclusivas e ambas tentam explicar o mesmo facto: os organismos subterrâneos e fossadores têm uma TMB inferior aos organismos epígeos e esta é uma das capacidades que lhes permite ocupar o meio subterrâneo.
A medição da TMB é difícil, considerando a necessidade de controlar diversos parâmetros sob condições experimentais. Devido a este facto, costuma ser utilizada a designação de taxa de metabolismo em repouso (TMR) que implica medições sob condições menos restritas (Sedláček, 2007).
22 As TMR são superiores no rato-toupeira relativamente ao rato-toupeira mediterrânico (Monarca, 2008). Madureira (1984) sugere que este facto se deve às características do solo e à temperatura dos locais que as estas espécies ocupam, visto que o rato-toupeira parece ter uma actividade mais superficial, enquanto que o rato-toupeira mediterrânico parece apresentar um estilo de vida mais dependente do meio subterrâneo (Vinhas, 1993; Paradis, 1995; Santos et al., 2009), como referido anteriormente.
Visto que o estilo de vida dos ratos-toupeira pode influenciar a redução da taxa metabólica basal seria importante saber se existem diferenças em termos de TMB entre as espécies parentais e os híbridos, e se as caraterísticas genéticas dos híbridos são transmitidas igualmente da parte materna e da parte paterna ou se, por outro lado, um dos sexos ou espécies domina sobre o/a outro/a a nível fisiológico. Assim, um dos objectivos deste trabalho consiste na determinação da TMR dos híbridos a diferentes temperaturas e na sua comparação com a TMR obtida para cada uma das espécies parentais (determinadas num trabalho anterior; Monarca, 2008) a essas mesmas temperaturas. Desta maneira poderá assim avaliar-se se a combinação genética das características das espécies parentais será vantajosa ou não para os híbridos.
23
3. Material e métodos
Este trabalho inseriu-se num projecto em curso que tem como objectivo determinar a existência das barreiras reprodutoras entre as duas espécies de ratos-toupeira, M. duodecimcostatus e M. lusitanicus. Para tal foi estabelecida no laboratório de micromamíferos da Faculdade de Ciências da Universidade de Lisboa uma colónia das duas espécies, tendo sido efectuados cruzamentos intra- e heterospecíficos e respectivos backcrosses. O presente trabalho beneficia da existência desses mesmos indivíduos, não sendo por isso aqui explicada a metodologia da experiência que levou à existência dos mesmos, a não ser quando necessário à compreensão do trabalho aqui apresentado.
3.1. Manutenção dos animais
Neste trabalho foram estudadas duas classes de animais híbridos resultantes do cruzamento de M. duodecimcostatus e M. lusitanicus em laboratório, nomeadamente, indivíduos MDL (♀ M. duodecimcostatus x ♂ M. lusitanicus) e MLD (♀ M. lusitanicus x ♂ M. duodecimcostatus), assim como indivíduos das duas espécies parentais (2ª e 3ª geração).
Figura 3. Híbridos resultantes do cruzamento de Microtus duodecimcostatus e M. lusitanicus.
Classe MDL (♀ M. duodecimcostatus x ♂ M. lusitanicus) (esquerda) e classe MLD (♀ M. lusitanicus x ♂ M. duodecimcostatus) (direita).
24 Tendo em conta as características fossadoras e sociais das duas espécies os animais foram mantidos em grupos de 2 a 3 indivíduos do mesmo sexo em gaiolas Makrolon tipo 3 com cerca de 3-4 cm de solo. De modo a fornecer enriquecimento ambiental foram também introduzidos em cada gaiola, palha para a construção do ninho, pequenos troncos de madeira, pinhas e pequenas pedras. Aos animais foi fornecido alimento fresco à base de maça, pêra, cenoura e alface ad libitum três vezes por semana, complementado com bolota e alfarroba. Não foi fornecida água, já que ambas as espécies não parecem beber água em forma livre, apenas aproveitando a contida nos alimentos. Os animais foram mantidos em condições de fotoperíodo (12h de luz : 12h de escuro) e temperatura (20 ± 2ºC) controladas durante o decorrer das experiências.
3.2. Crescimento e fertilidade
De modo a estudar o crescimento e desenvolvimento dos híbridos no laboratório, os casais intraespecíficos (já existentes no laboratório) foram monitorizados a cada dois dias, de modo a verificar a existência de novas ninhadas e acompanhar o desenvolvimento das crias já existentes. O peso das crias foi registado sempre que possível desde o seu nascimento até cerca dos 110 dias de idade de modo a estimar a curva de crescimento das duas classes de híbridos, MDL e MLD. Apenas foram calculadas as curvas de crescimento respeitantes às duas classes de híbridos, uma vez que o número de indivíduos híbridos de backcross monitorizados foi muito baixo.
A fertilidade das duas classes de híbridos foi determinada tendo em conta os seguintes parâmetros: a) número de casais que produziram descendência; b) número de ninhadas; c) tamanho das ninhadas; d) razão dos sexos. Oito tipos de backcrosses foram examinados: fêmea híbrida x macho parental (MDL x MD, MDL x ML, MLD x MD e MLD x ML) e fêmea parental x macho híbrido (MD x MDL, ML x MDL, MD x MLD e ML x MLD). Foram ainda analisados dois tipos de cruzamentos entre as duas classes de híbridos: MDL x MDL e MLD x MLD.
3.2.1. Análise estatística
A distribuição de Gompertz é uma distribuição contínua de probabilidade representada por uma função sigmoide. A função de Gompertz é um modelo de regressão
25 não linear adequado para comparar padrões de crescimento entre diferentes espécies de mamíferos (Zullinger et al., 1984). Os dados do peso e da idade dos animais foram colocados no SPSS para uma análise de regressão não linear em que se inseriu a equação de Gompertz (1), onde X(t) é o valor da variável X à idade t, A é o valor da assimptota da variável, K é a taxa de crescimento constante e I é a idade no ponto de inflecção da curva.
(1)
Os valores dos parâmetros A, K e I da curva que foram inseridos na análise para ajustar as constantes de crescimento corresponderam aos da espécie Microtus cabrerae: K=0,0567; A=36,4; I=16 (Fernández-Salvador et al., 2001), visto que é uma espécie irmã de M. duodecimcostatus e M. lusitanicus e que as suas características morfológicas não diferem muito das classes de híbridos das espécies estudadas. O objectivo da regressão foi estimar os valores dos 3 parâmetros (A, K e I) para cada classe de híbridos de acordo com o peso e a idade, sendo depois utilizados para determinar e representar as respectivas tendências do crescimento corporal em curvas de Gompertz.
Relativamente à fertilidade, foi feita uma análise estatística descritiva dos dados no programa SPSS, sendo sumarizados em médias e medidas de dispersão.
3.3. Análise dos Espermatozóides 3.3.1. Procedimento experimental
O protocolo experimental (modificado de Gómez Montoto et al., 2011b) seguido nesta experiência encontra-se no Anexo I.
Os animais das duas classes de híbridos e das duas espécies parentais foram pesados, sacrificados por deslocamento cervical e dissecados. Para obter espermatozóides maduros foram removidas ambas as caudas dos epidídimos e colocadas num gobelet (2 cm de diâmetro) pré-aquecido a 37ºC, contendo 250 µl de Hepes-buffered modified Tyrode's medium (mT-H) previamente preparado (ver Anexo II). De modo a permitir a libertação dos espermatozóides para o meio foram efectuadas algumas incisões nos epidídimos e incubados durante 10 minutos sobre uma placa de aquecimento a 37ºC, onde foi mantido o gobelet durante todo o procedimento. Os testículos de cada indivíduo foram também
26 removidos, sendo o esquerdo pesado e preservado em Bouin e o direito preservado em RNA later para estudos posteriores.
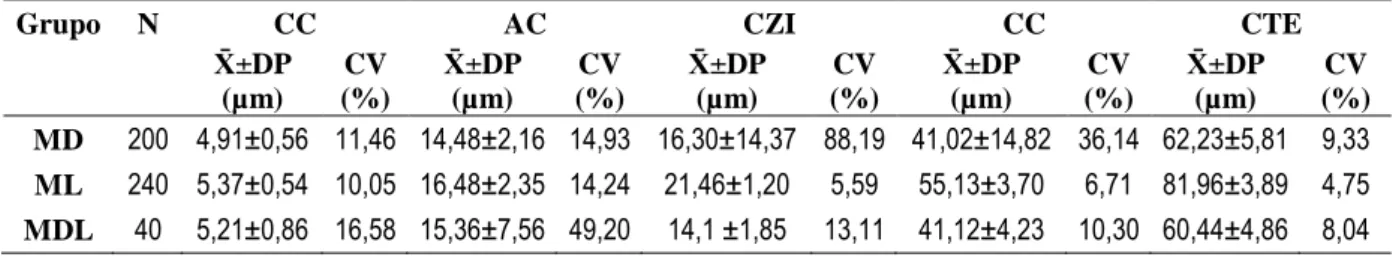
Para a análise da morfologia e viabilidade do esperma, a solução mT-H (50 µl) foi misturada num vidro de relógio com uma solução corante Eosina-Nigrosina (50 µl) previamente preparada (Anexo III). A suspensão foi depois incubada durante 30s à temperatura ambiente e realizados seis esfregaços (uma gota de 10 µl por lâmina) por animal (modificado de Björndahl et al., 2003). Após a sua secagem, as lâminas foram coradas em Giemsa a 10% durante 1 hora, lavadas e secas ao ar. Foram então fotografadas 20 células por macho num microscópio (Olympus BX60) acoplado a uma câmera fotográfica digital (Hamamatsu R2 cooled CCD) com uma lente de 60x 0.95NA. As dimensões dos espermatozóides foram obtidas traçando manualmente a curvatura de cada espermatozóide utilizando o software ImageJ v.1.47. Foram quantificados os seguintes parâmetros: a) comprimento da cabeça (CC), b) área da cabeça (AC), c) comprimento da zona intermédia (CZI); d) comprimento da cauda (CC) e e) comprimento total do espermatozóide (CTE). Todas as anomalias morfológicas encontradas foram também descritas.
De modo a analisar a mobilidade dos espermatozóides efectuou-se uma preparação com uma gota de 10 µl da solução mT-H com espermatozóides numa lâmina e cobriu-se com uma lamela de 18 x 18 mm, sendo de seguida captados vídeos de aproximadamente 5 segundos ao microscópio (Olympus BX60) acoplado a uma câmera de vídeo digital (CMOS TIS DFK 72BUC02) com uma lente de 10x 0.3NA (protocolo modificado de Gómez Montoto et al., 2011b). Os vídeos foram obtidos em modo de aquisição rápida numa área restrita do detector para permitir taxas de aquisição de 120 Hz, usando o software ICCapture v2.3. De modo a diminuir a adesão ao vidro por parte dos espermatozóides durante a recolha dos vídeos, foram usadas lâminas previamente preparadas com BSA (Albumina de Soro Bovino). Para tal, as lâminas foram primeiro mergulhadas durante 3 horas numa solução de 1% de ácido acético à temperatura ambiente. Em seguida as lâminas foram lavadas com água corrente durante 5 minutos, e após este período mergulhadas em água destilada e deixadas secar à temperatura ambiente. Por último as lâminas foram mergulhadas numa solução de 1% de BSA (diluída em tampão PBS) e secas ao ar num local protegido do pó.
27 Os vídeos foram gravados em formato "avi" e depois processados e analisados traçando manualmente o trajecto percorrido de cada espermatozóide utilizando o software ImageJ v.1.47. O parâmetro medido para este trabalho foi a velocidade curvilínea (VCL).
3.3.2. Análise estatística
Para comparação das medições corporais e das dimensões dos espermatozóides foi utilizada uma fórmula que calcula o coeficiente de variância (CV) (2), em que DP = desvio padrão e M = média.
(2)
De modo a estudar a relação entre a massa corporal e a massa dos testículos nas duas classes de híbridos e nas espécies parentais foi calculada a massa relativa dos testículos (MRT) para cada grupo. Após ser verificada a normalidade dos dados através do teste Shapiro-Wilk, a MRT de cada grupo foi então comparada através de uma análise de variância (ANOVA one-way) e posteriormente com o teste post-hoc Games-Howell (verificando-se a igualdade de variâncias) de modo a avaliar a existência de diferenças entre os grupos em estudo.
Relativamente às dimensões dos espermatozóides foi feita uma análise estatística descritiva, para além do cálculo do CV. Foi depois relacionado o comprimento total dos espermatozóides (CTE) com a massa relativa dos testículos e por fim foram analisadas as diferenças do CTE entre os grupos teste com uma ANOVA one-way após verificar a normalidade da distribuição dos dados com os testes Shapiro-Wilk e Kolmogorov-Smirnov.
Posteriormente foi efectuado um teste T independent-samples de modo a comparar a velocidade curvilínea (VCL) dos espermatozóides. A normalidade das amostras das diferentes espécies foi verificada através do teste Kolmogorov-Smirnov. Todos os testes estatísticos ao longo do trabalho foram efectuados no programa IBM SPSS Statistics versão 20. Em todos os testes o nível de significância utilizado foi de α=0,05 sendo a hipótese nula rejeitada sempre que o p-value foi inferior a 0,05.
28
3.4. Testes de preferência sexual 3.4.1. Procedimento experimental
A metodologia escolhida é a descrita para este tipo de estudo tendo já sido validada por inúmeros estudos em pequenos mamíferos (Christophe & Baudoin, 1998; Smadja & Ganem, 2002, 2005). Foram feitas apenas algumas adaptações visto que se estão a estudar espécies diferentes.
Na presente experiência pretendeu-se estudar a preferência sexual dos machos das espécies M. lusitanicus e M. duodecimcostatus entre fêmeas conspecíficas e fêmeas híbridas com base em estímulos odoríferos, nomeadamente a urina e os dejectos. Não foi testada a preferência sexual dos machos de nenhuma das duas classes de híbridos visto que, mesmo que estes animais manifestem preferência por um determinado tipo de fêmea estes animais não produzem descendência (Cerveira et al., em preparação). Assim sendo, a determinação da sua preferência sexual não parece ser de grande relevância em termos evolutivos. Pelo contrário, e uma vez que as fêmeas híbridas são férteis, foram testados apenas os machos das duas espécies parentais relativamente aos seus conspecíficos e às duas classes de fêmeas híbridas. O facto de apenas serem testados machos e não fêmeas apoia-se na verificação de que as fêmeas das duas espécies de rato-toupeira não apresentam preferência sexual por indivíduos conspecíficos (Cerveira et al., em preparação).
Todas as fêmeas utilizadas como dadoras do estímulo químico estavam em repouso reprodutor , apresentando a vagina colada (Keller, 1985). De modo a evitar a indução de estro (período em que estão sexualmente receptivas), as fêmeas foram mantidas durante to o decorrer da experiência com outras fêmeas, não havendo qualquer contacto com machos de qualquer uma das espécies.
A preferência sexual dos indivíduos foi avaliada utilizado um olfactómetro em vidro (Fig. 4), composto por uma caixa de partida (30 x 20 x 15 cm), onde se coloca o animal a ser testado (i.e., animal teste), ligada por um tubo em forma de Y (braços secundários=30 cm, braço central=35 cm, ø interno=5 cm) a duas caixas-estímulo (20 x 20 x 15 cm) onde são colocados os estímulos odoríferos.
29
Figura 4. Olfactómetro (modificado de Smadja & Ganem, 2002).
Os estímulos foram recolhidos colocando cada fêmea dadora na caixa-estímulo correspondente durante 1 hora. Ao fim dessa hora, as fêmeas dadoras foram transferidas novamente para as suas gaiolas e as caixas-estímulo encaixadas nos braços do Y. Em seguida, o macho teste foi inserido na caixa de partida dando início à experiência. Cada teste teve início assim que o animal saiu da caixa de partida e entrou no Y, tendo uma duração de 5 min. Após a entrada no Y o animal pode explorar livremente todo o olfactómetro, incluindo os dois estímulos odoríferos que lhe são apresentados. Em geral, os animais entraram no Y ao fim de alguns minutos, no entanto caso o macho não entrasse no Y ao fim de 10 min ou caso entrasse imediatamente no Y após a sua colocação na caixa de partida, o teste era anulado e o macho testado novamente uma semana depois.
Os machos de M. duodecimcostatus e M. lusitanicus foram submetidos a duas experiências de modo a testar a preferência entre fêmeas conspecíficas e as duas classes de fêmeas híbridas. A experiência 1 consistiu na exposição do animal teste ao estímulo de uma fêmea conspecífica e ao de uma fêmea híbrida MDL, enquanto a experiência 2 consistiu na exposição do animal teste ao estímulo de uma fêmea conspecífica e a outro de uma fêmea MLD.
Todos os testes foram filmados (Canon Legria HF S200) de modo a permitir a posterior análise da direção preferencial tomada pelo macho em relação ao estímulo
30 odorífero das fêmeas. Os vídeos foram analisados com o software JWatcher tendo sido registados:
a) o tempo de latência (tempo desde que o animal é colocado na caixa de partida até entrar no Y);
b) o tempo que o animal passou em cada caixa-estímulo ("caixa");
c) o tempo que passou no Y;
d) o tempo que passou a cheirar ou lamber os estímulos de cada fêmea ("estímulo").
Depois de cada teste o material foi todo lavado com água e com uma solução de álcool a 40%. Foi também alternada a posição (esquerda ou direita) da caixa-estímulo que continha o estímulo da fêmea conspecífica vs estímulo da fêmea híbrida de modo a excluir um possível efeito de lateralidade, ou seja, de os indivíduos terem uma maior tendência a escolher um dos braços do Y independentemente do estímulo presente. Todas as experiências foram realizadas entre as 10 e as 19 horas, sendo todos os animais usados neste estudo indivíduos adultos. Nenhum dos animais foi usado mais do que uma vez em cada experiência.
3.4.2. Análise de dados
Para cada macho foi utilizada uma equação (3) para estimar a razão da preferência (R) entre as fêmeas dadoras. Esta razão varia entre -1 e +1, em que um R=0 indica que não há preferência por nenhum dos estímulos, um R negativo corresponde a uma preferência pelo estímulo heteroespecífico e um R positivo a uma preferência pelo estímulo conspecífico.
(3)
Na análise da preferência sexual foram calculados dois tipos de índice R: a) Rcaixa que corresponde à razão do tempo passado nas caixas-estímulo, e b) Restímulo que
31 corresponde à razão do tempo dentro da caixa e em contacto (i.e., a cheirar ou lamber) com o estímulo.
3.4.3. Análise estatística
Os índices Rcaixa e Restímulo foram usados para verificar a normalidade da distribuição dos dados (teste Shapiro-Wilk visto que o número da amostra (N=16) é menor que 50). Nos casos em que se verificou a normalidade dos dados efectuou-se uma análise do tempo passado em cada caixa e do tempo passado em contacto com o estímulo com o teste paramétrico T paired samples para verificar a preferência através da comparação da média das amostras. Nos casos em que a distribuição dos dados não foi normal utilizou-se o teste Wilcoxon que em vez das médias compara as medianas. Os testes estatísticos foram efectuados no programa IBM SPSS, sendo considerado um valor de significância de α=0,05.
3.5. Metabolismo Basal
3.5.1. Procedimento experimental
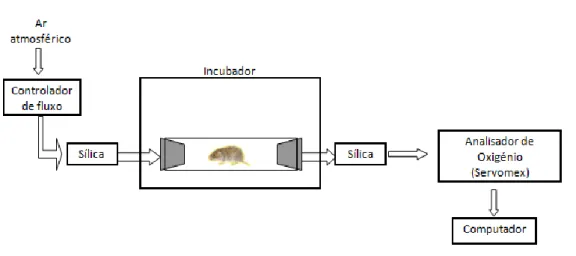
Com esta experiência pretendeu-se determinar a taxa de metabolismo em repouso (TMR) de híbridos das classes MDL e MLD através da medição do volume de oxigénio (VO2) consumido a diferentes temperaturas ambiente. A medição da TMR foi efectuada através de um sistema de respirometria de fluxo aberto com pressão positiva (Fig. 5). Este sistema é constituído por um tubo de acrílico transparente (comprimento=29 cm, ø externo=8 cm e espessura=4 mm) fechado com duas rolhas de borracha colocadas nas suas extremidades. Durante cada teste, este tubo (onde se coloca o animal) é mantido dentro de um incubador de modo a atingir a temperatura pretendida. O ar (do exterior) é introduzido no sistema a uma taxa de fluxo de 500 ml/min, com o auxílio de uma bomba, passando previamente por um tubo de sílica para remover a humidade. Em seguida o ar entra no tubo e volta a sair passando novamente por sílica e o volume de oxigénio em partes por milhão (ppm) é medido (Servomex série 4100 ligado a um computador) (modificado de Arch et al., 2006). De modo a oferecer ao animal uma superfície de repouso afastada das paredes
32 do tubo durante o decorrer do teste este é colocado sobre uma base retangular (5 x 25 cm) de arame.
Figura 5. Sistema de respirometria de fluxo aberto (imagem cortesia de R.I. Monarca).
Em cada teste, o tubo de acrílico é colocado no incubador à temperatura pretendida, são-lhe inseridos os tubos de passagem do ar e permanece assim durante aproximadamente 15 minutos para deixar o valor de oxigénio estabilizar no seu interior. Durante este período, o animal a ser testado é pesado e a sua temperatura corporal medida. Assim que o valor de oxigénio estabiliza introduz-se o animal no tubo, fecha-se o sistema e durante 2 horas é medida a quantidade de oxigénio presente no fluxo de ar. Ao fim deste tempo retira-se o animal do sistema, sendo novamente pesado e a sua temperatura corporal medida. O sistema mantem-se ligado por mais um intervalo de 15 minutos de modo a que o valor de oxigénio volte ao patamar inicial e se estabeleça uma baseline.
Esta experiência foi efectuada apenas com machos híbridos tendo cada um deles sido submetido a dois testes em dias consecutivos e diferentes períodos do dia para evitar a influência que o ciclo diário poderia ter na taxa de metabolismo dos mesmos (Halle & Stenseth, 1994).
O consumo de oxigénio foi medido a três temperaturas: 5ºC, 20ºC e 30ºC. Para as espécies parentais M. lusitanicus e M. duodecimcostatus, os intervalos da zona termoneutral são 32,5-35ºC e 32,5-37,5ºC, respectivamente (Monarca, 2008). Visto isto, a
33 temperatura de 30ºC foi escolhida por ser uma temperatura próxima da zona termoneutral, onde os animais gastam o mínimo de energia para a sua termorregulação; 5ºC é uma temperatura baixa tendo em conta o ambiente a que estas espécies estão sujeitas, possibilitando assim analisar o efeito da variável temperatura nas taxas de metabolismo destes animais; 20ºC é uma temperatura intermédia que já permite identificar um maior gasto de energia para a termorregulação. Esta é também a temperatura à qual os animais estão aclimatados no laboratório.
3.5.2. Análise de dados
Os valores de oxigénio foram medidos a cada 30 segundos ao longo das 2 horas. Em cada teste, a taxa de metabolismo em repouso foi estimada tendo em conta o valor base do patamar inicial e o valor médio das 5 leituras consecutivas mais altas de O2 (correspondente a 2min e 30s no total das 2 horas) através da fórmula de Depocas & Hart (1957) (4), onde V2 é a taxa de fluxo do ar depois do incubador e F1O2 e F2O2 são as concentrações de oxigénio do ar antes e depois do incubador.
(4)
Todas as medições de VO2 foram corrigidas padronizando os valores de temperatura e pressão (Monarca, 2008).
3.5.3. Análise estatística
Sabendo que o peso é o factor que mais influencia o consumo de oxigénio de um indivíduo, de modo a poder comparar o metabolismo dos dois grupos animais testados, foi primeiro necessário determinar se existiam diferenças significativas de peso entre os mesmos. Após ser verificada a normalidade dos dados (teste de Shapiro-Wilk) foi efectuada uma análise de variâncias (ANOVA one-way) do peso das duas classes de híbridos. Havendo igualdade de variâncias verificou-se se o peso era significativamente diferente com o teste post-hoc Tukey. Quando a igualdade de variâncias não foi verificada utilizou-se o teste post-hoc Games-Howell.
34 Tendo as duas classes de híbridos pesos significativamente diferentes foi então efectuada uma análise de covariância (ANCOVA) considerando o peso como co- variável e o VO2 consumido pelo animal como variável dependente. A normalidade (teste de Shapiro-Wilk) da variável dependente e a homogeneidade de variâncias (teste de Levene) foram também verificadas. Por fim, esses resultados foram comparados com os dados de volume de oxigénio consumido pelas espécies parentais às mesmas temperaturas (5ºC, 20ºC e 30ºC) determinados num trabalho anterior (Monarca et al., em preparação). Todos os testes estatísticos foram efectuados no programa IBM SPSS, sendo considerado um valor de significância de α=0,05.
35
4. Resultados
4.1. Crescimento e fertilidade
O peso corporal dos animais híbridos (MDL e MLD) foi registado até aos 110 dias de vida. Estes dados foram utilizados para determinar as respectivas curvas de crescimento recorrendo à equação de Gompertz. As curvas de crescimentos das espécies parentais obtidas num trabalho anterior (Cerveira et al., em preparação) foram também representadas de modo a poder avaliar possíveis diferenças entre os dois grupos (Fig. 6).
Taxas de crescimento de 0,058 ± 0,001 (N=1500) e 0,057 ± 0,002 (N=1157) (parâmetro K na curva sigmoide de Gompertz) foram observadas para as duas classes de híbridos MLD e MDL, respectivamente. A taxa de crescimento mais elevada foi observada nos primeiros 30-40 dias de idade para as duas classes de hibridos, atingindo o seu máximo (parâmetro I da equação de Gompertz) no início do período de desmame (aos 14 dias de idade na classe MLD e 13 dias de idade na classe MDL). A partir dos 30 a 40 dias de idade verificou-se um acentuado decréscimo da taxa relativa de crescimento nas duas classes de híbridos, estabilizando por volta dos 70 dias de idade para a classe MLD e 75-80 dias para a classe MDL. Um peso máximo (parâmetro A na curva sigmoide de Gompertz) de 18,7 ± 0,10 g e 24,2 ± 0,18 g foi atingido para as classes MLD e MDL, respectivamente.
Relativamente às espécies parentais, as duas classes de híbridos têm taxas de crescimento e pesos máximos intermédios, com a maior taxa de crescimento para a espécie M. lusitanicus (N=2947) (K=0,062 ± 0,001; A=16; I=12), que atinge o peso máximo mais cedo comparativamente aos outros grupos analisados (por volta dos 60 dias), mas que também tem o menor peso máximo médio. M. duodecimcostatus (N=2962) tem a menor taxa de crescimento (K=0,047 ± 0,001; A=26,7; I=18) e o seu peso médio máximo é atingido aos 80 dias. M. duodecimcostatus é no entanto o grupo de indivíduos testados que atinge um maior peso médio e que tem a taxa máxima de crescimento mais tardia (aos 18 dias) (Cerveira et al., em preparação).
36
Figura 6. Peso das duas espécies de rato-toupeira, Microtus lusitanicus (ML), M. duodecimcostatus (MD), (Cerveira et al., em preparação) e seus híbridos
MLD (♀ M. lusitanicus x ♂ M. duodecimcostatus) e MDL (♀ M. duodecimcostatus x ♂ M. lusitanicus) relativamente à sua idade. Curvas de crescimento para cada grupo obtidas através da aplicação do modelo de Gompertz.
0 5 10 15 20 25 30 35 40 45 0 20 40 60 80 100 120 P eso ( g ) Tempo (dias) ML Peso ML Gompertz MD Peso MD Gompertz MLD Peso MLD Gompertz MDL Peso MDL Gompertz
37 Os dados de fertilidade registados para os backcrosses (N=48) e para os cruzamentos híbrido x híbrido (N=9) estão representados na tabela 1. Nos cruzamentos híbrido x híbrido não foi observada descendência. De todos os backcrosses formados, nenhum dos casais em que o híbrido era o macho produziu descendência, independentemente da classe do híbrido (i.e., macho MDL ou macho MLD). Nos backcrosses com fêmeas híbridas houve descendência em 61,5% e 72,7% dos pares dependendo da classe do híbrido (fêmea MLD e fêmea MDL, respectivamente). Os backcrosses com machos M. lusitanicus tiveram um sucesso reprodutor relativamente superior (75,0%) relativamente aos backcrosses envolvendo machos M. duodecimcostatus (58,3%).
O tamanho das ninhadas foi semelhante nos diferentes tipos de backcrosses, sendo no entanto ligeiramente superior nos backcrosses envolvendo fêmeas híbridas MDL. Relativamente à razão dos sexos (♂:♀), houve em geral uma predominância do sexo feminino na descendência dos backcrosses, especialmente nos cruzamentos com machos da espécie M. lusitanicus (33:67 para as fêmeas MDL e 17:83 para as fêmeas MLD).
38
Tabela 1. Parâmetros de fertilidade dos backcrosses das duas espécies de rato-toupeira, Microtus
duodecimcostatus (MD) e M. lusitanicus (ML). MDL - híbrido (♀ M. duodecimcostatus x ♂ M. lusitanicus); MLD - híbrido (♀ M. lusitanicus x ♂ M. duodecimcostatus); R - pares que se reproduziram; NR - pares que não se reproduziram. Apenas os pares que estavam juntos há um mínimo de 3 meses foram contabilizados.
Pares Tamanho da Ninhada Sexo da ninhada
Cruzamento R NR Total ± EP Mín. Máx. N ♂ ♀ Razão dos sexos (%) (♂:♀) Backcrosses ♀ MDL x ♂ MD 4 (66.7%) 2 6 1,3 ± 0.141 1 2 11 6 4 60:40 ♀ MDL x ♂ ML 4 (80.0%) 1 5 1,2 ± 0.166 1 3 13 5 10 33:67 ♀ MLD x ♂ MD 3 (50.0%) 3 6 1,1 ± 0.143 1 2 7 3 5 34:63 ♀ MLD x ♂ ML 5 (71.4%) 2 7 1,1 ± 0.125 1 2 8 5 1 17:83 ♀ MD x ♂ MDL 0 5 5 - - - - ♀ ML x ♂ MDL 0 6 6 - - - - ♀ MD x ♂ MLD 0 5 5 - - - - ♀ ML x ♂ MLD 0 8 8 - - - - Total de Backcrosses 16 32 48 - - - 39 - - - Híbrido x Híbrido ♀ MDL x ♂ MDL 0 5 5 - - - - - - - ♀ MLD x ♂ MLD 0 4 4 - - - - - - - Total Híb x Híb 0 9 9 - - - - - - - Total 16 41 57 - - - 39 - - -
39
4.2. Análise dos Espermatozóides 4.2.1. Medições corporais
Foram analisados 45 animais no total, entre M. lusitanicus (N=12), M. duodecimcostatus (N=11) e híbridos MDL (N=11) e MLD (N=9).
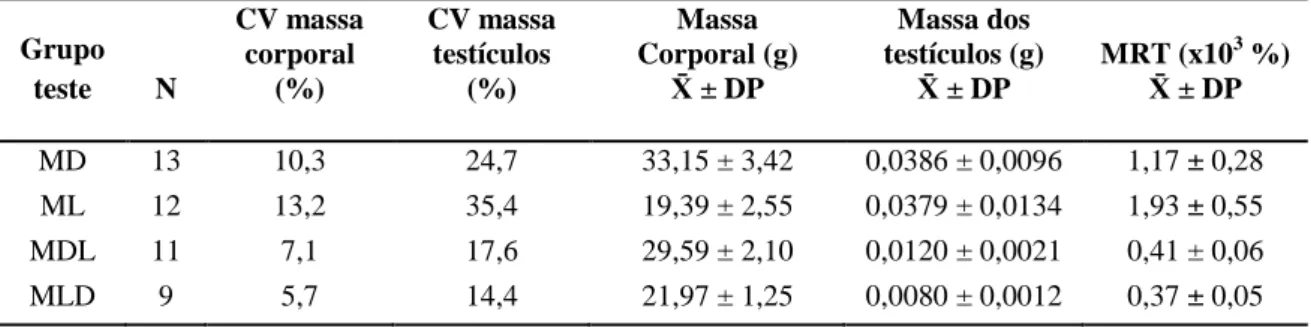
Na presente amostra de estudo destas espécies e dos dois tipos de híbridos, o peso corporal teve um coeficiente de variância (CV) de 23,8%, com um peso mínimo médio de 19,39 ± 0,74 g (média ± EP) em M. lusitanicus, e um peso máximo médio de 33,15 ± 0,95 g em M. duodecimcostatus. Os híbridos apresentaram um peso corporal intermédio, tendo os machos da classe MDL maior peso relativamente aos machos MLD (Tabela 2). Ambas as classes de híbridos apresentaram CV mais baixos que as espécies parentais. Já a variação da massa dos testículos foi muito maior (CV=63,8%), variando entre 0,0080 ± 0,0004 g nos híbridos da classe MLD, e 0,0386 ± 0,0026 g nos M. duodecimcostatus. A massa média dos testículos dos animais da classe MDL (0,0120 ± 0,0006 g) apresentou um valor próximo do valor da massa média dos testículos em MLD, sendo ambos consideravelmente menores que os valores obtidos para as espécies parentais.
Tabela 2. Coeficientes de variância (CV), médias das medições corporais e massa relativa dos
testículos (MRT) dos grupos de teste, Microtus duodecimcostatus (MD), M. lusitanicus (ML), e das duas classes de híbridos (MDL e MLD).
Grupo teste N CV massa corporal (%) CV massa testículos (%) Massa Corporal (g) ± DP Massa dos testículos (g) ± DP MRT (x103 %) ± DP MD 13 10,3 24,7 33,15 ± 3,42 0,0386 ± 0,0096 1,17 ± 0,28 ML 12 13,2 35,4 19,39 ± 2,55 0,0379 ± 0,0134 1,93 ± 0,55 MDL 11 7,1 17,6 29,59 ± 2,10 0,0120 ± 0,0021 0,41 ± 0,06 MLD 9 5,7 14,4 21,97 ± 1,25 0,0080 ± 0,0012 0,37 ± 0,05
40
Figura 7. Dispersão do logaritmo das médias da massa dos testículos relativamente ao logaritmo
das médias da massa corporal de Microtus duodecimcostatus (MD), M. lusitanicus (ML) e das duas classes de híbridos (MDL e MLD). Os símbolos a negro representam os dados obtidos no presente trabalho e os símbolos brancos representam os dados de Gómez Montoto et al., 2011a para as duas espécies.
Após ser analisada a massa relativa dos testículos (MRT) das quatro amostras de animais testados verificou-se que os indivíduos M. lusitanicus são aqueles que têm uma maior massa dos testículos relativamente ao peso do corpo (MRTML=1,93x103). Nos indivíduos da espécie M. duodecimcostatus a média da MRT é ligeiramente inferior (MRTMD=1,17x103), apresentando os híbridos valores muito inferiores comparativamente às espécies parentais (MRTMDL=0,41x103 e MRTMLD=0,37x103). A MRT foi significativamente diferente entre os 4 grupos testados (ANOVA one-way; F3,41=57,17;
p<0,05) excepto entre as duas classes de híbridos, MDL e MLD (post-hoc Games-Howell; p=0,43; NS; N=20).
4.2.2. Dimensões do esperma
Foram analisados seis esfregaços por cada animal amostrado. Dos 9 indivíduos amostrados da classe de híbridos MLD não foram encontrados espermatozóides em nenhuma das lâminas. Das amostras recolhidas aos 11 indivíduos MDL apenas se observaram espermatozóides num indivíduo e numa única lâmina. Pelo contrário, foi
-2,5 -2,3 -2,1 -1,9 -1,7 -1,5 -1,3 -1,1 -0,9 -0,7 -0,5 1 1,2 1,4 1,6 1,8 2 L o g m a ss a do s testíc ulo s
Log massa corporal
MD ML MDL MLD MD ML