Vasorelaxant effects of 1-nitro-2-phenylethene in rat isolated aortic rings
Loeste Arruda-Barbosa
a, Karilane Maria Silvino Rodrigues
a, Francisco das Chagas Vasconcelos Souza-Neto
a,
Gloria Pinto Duarte
b, Rosivaldo S. Borges
c, Pedro Jorge Caldas Magalhães
a, Saad Lahlou
a,⁎
aDepartment of Physiology and Pharmacology, Federal University of Ceará, CE, BrazilbDepartment of Physiology and Pharmacology, Federal University of Pernambuco, Recife, PE, Brazil cDepartment of Pharmacy, Federal University of Pará, Belém, PA, Brazil
a b s t r a c t
a r t i c l e
i n f o
Article history:
Received 14 January 2014
Received in revised form 27 July 2014 Accepted 1 August 2014
Available online 15 August 2014
Chemical compounds studied in this article:
1-Nitro-2-phenylethene (PubChem CID: 5284459)
Keywords:
Isolated thoracic aorta
Endothelium-independent vasorelaxation Myogenic
1-Nitro-2-phenylethene Normotensive rat
Previously, we showed that nitro-2-phenylethane is a vasorelaxant constituent of the essential oil ofAniba canelilla. Here, we investigated the mechanisms underlying the vascular effects of 1-nitro-2-phenylethene (NPe), a structural analog of 1-nitro-2-phenylethane obtained synthetically, in rat isolated thoracic aortic prep-arations. At 0.1–100μg/mL, NPe similarly relaxed endothelium-intact or endothelium-denuded aortic prepara-tions pre-contracted with 60 mM KCl or with phenylephrine (PHE, 1μM). Vasorelaxant effects of NPe against PHE-induced contractions remained unaffected following blockade of potassium channels by TEA, and inhibition of either nitric oxide synthase byL-NAME, cyclooxygenase by indomethacin or guanylate cyclase by ODQ. In
preparations maintained under Ca2+-free conditions, NPe signi
ficantly reduced the contractions induced (i) by PHE, but not those evoked by caffeine, (ii) by CaCl2in either PHE (in the presence of 1μM verapamil)- or KCl-stimulated preparations, (iii) by extracellular Ca2+restoration in thapsigargin-treated aortic preparations, and (iv) by the activator of protein kinase C phorbol-12,13-dibutyrate or the inhibitor of protein tyrosine phosphatase sodium orthovanadate. It is concluded that NPe induced an endothelium-independent vasorelaxation with potency greater than its structural analog 1-nitro-2-phenylethane. Such action appears to occur intracellularly probably through inhibition of contractile events that are clearly independent of Ca2+in
flux from the extracellular milieu.
© 2014 Elsevier Inc. All rights reserved.
1. Introduction
Hypertension is a common and progressive disorder which consti-tutes a major risk for diabetes, stroke, as well as cardiovascular and renal diseases. A great number of researches focused on the bioactive compounds from natural resources as potential substances for the treat-ment of hypertension. Efforts to this effect have been pursued by our group for a number of years to identify novel antihypertensive
compounds with a vasodilator activity especially derived from essential oils of aromatic plants from the north–northeast region of Brazil[1–8]. Nitroderivatives found in higher plants are rare. 1-Nitro-2-phenylethane is thefirst nitro compound isolated from plants[9], and is considered to be responsible for the plant's cinnamon scent[10]. It is the main constit-uent of the essential oil ofAniba canelilla(H.B.K.) Mez [syn.Aniba elliptica A. C. Sm.,Cryptocarya canelillaKunth] (EOAC), (Lauraceae), an aromatic
plant abundant in the Amazon region commonly known as“
casca-preciosa”(precious bark).
Previously, we showed that intravenous (i.v.) administration of 1-nitro-2-phenylethane (Fig. 1A) induced two periods of hypotension and bradycardia in either normotensive[11]or hypertensive[12]rats. Initially a rapid bradycardia (onset time of 1–2 s) occurred coincidental-ly (onset time of 2 s) with an arterial hypotension (phase 1) and then, a delayed decrease in blood pressure (peak between 4 and 10 s) associat-ed with a second bradycardia (phase 2)[11,12]. Several lines of
evi-dence confirmed that phase 1 (rapid bradycardiac and depressor
responses to EOAC and 1-nitro-2-phenylethane) was of reflex origin (vago-vagal reflex). Phase 1 appears not to involve the activation of ei-ther vanilloid TPRV1or 5-HT3receptors located on vagal sensory nerves
[11,12]. The second hypotensive response (phase 2) of 1-nitro-2-phenylethane results from a direct vasodilatory effect, as evidenced by the concentration-dependent reduction of phenylephrine (PHE)-Abbreviations:cGMP, cyclic guanosine-5′-monophosphate; EGTA, ethylene glycol
tetraacetic acid; EOAC, essential oil ofAniba canelilla; i.v., intravenous; IP3, inositol
triphosphate; KHS, Krebs–Henseleit solution;L-NAME,L-N(G) nitroarginine methyl ester; NO, nitric oxide; NPe, 1-nitro-2-phenylethene; ODQ, 1H-[1,2,4]oxadiazolo[4,3-a] quinoxalin-1-one; PDB, phorbol-12,13-dibutyrate; PHE, phenylephrine; ROCCs, receptor-operated channels; SERCA, sarcoplasmic/endoplasmic reticulum Ca2 +pump; sGC, soluble
guanylate cyclase; SOCCs, store-operated Ca2 +channels; TEA, tetraethylammonium
chloride; VOCCs, voltage-operated calcium channels.
⁎ Corresponding author at: Department of Physiology and Pharmacology, Federal University of Ceará, Rua Cel. Nunes de Melo 1127, Fortaleza, CE, Brazil. Tel.: +55 85 33668334; fax: +55 85 33668333.
E-mail addresses:[email protected](L. Arruda-Barbosa),
karilane.fi[email protected](K.M.S. Rodrigues),[email protected]
(F.C.V. Souza-Neto),[email protected](G.P. Duarte),[email protected]
(R.S. Borges),[email protected](P.J.C. Magalhães),[email protected](S. Lahlou).
http://dx.doi.org/10.1016/j.vph.2014.08.002
1537-1891/© 2014 Elsevier Inc. All rights reserved.
Contents lists available atScienceDirect
Vascular Pharmacology
induced contractions in either isolated aortic[11]or superior mesenter-ic artery[12,13]preparations. Recently, it was shown that the vasodila-tor actions of 1-nitro-2-phenylethane appeared to derive from its ability to augment cGMP levels in aortic smooth muscle cells, afinding sugges-tive for the involvement of stimulating the soluble guanylate cyclase (sGC)–cGMP pathway in such vasodilator effects[14].
The sp3carbon atoms are responsible for different conformations of 1-nitro-2-phenylethane. Thus, a conformational restriction by substitu-tion of the alkane for the alkene moiety allows the formasubstitu-tion of 1-nitro-2-phenylethene (NPe,Fig. 1B) which may exhibit different degrees of potency or different types of activities. Therefore, the present investiga-tion was undertaken to assess the vascular effects of NPe in rat isolated thoracic aortic preparations and the putative mechanisms underlying these effects.
2. Materials and methods
2.1. Synthesis of 1-nitro-2-phenylethene
NPe or 1-((E)-2-nitro-vinyl)-benzene (β-nitrostyrene) was synthe-sized by employing the Claisen–Schmitd's procedure[15,16]using
benzaldehyde and nitromethane as substrates (0.1 and 0.12 eq., respec-tively). The benzaldehyde was‘one-pot’converted, in 89% yield, to the corresponding styrene by treatment with 0.05 eq. of NaOH in methanol and water at 0–10 °C.
The resulting precipitate wasfiltered out and dried under vacuum to give the desired styrene derivative, NPe. Thetransproduct is more preferential thancisdue to steroselective reactions. The title compound was crystallized in ethanol as a cream-colored solid; m.p. 56.2–58.6 °C (55–58 °C; Sigma-Aldrich standard). IR νmax 1600, 1550, 1498, 1375 cm−1;1H NMR (CD
3SOCD3, 200 MHz)δ7.2–7.3 (d, 2 H), 7.35– 7.4 (d, 2H), 7.55 (d, 1H), 7.7 (d, 1H), 7.9 (d, 1H);13C NMR (CD
3SOCD3, 50 MHz)δ127.52, 128.84, 129.69, 136.82, 137.99, 140.96. Thefinal product was identified by NMR (1H NMR) and FT-IR spectroscopic techniques and compared with literature[17].
2.2. Animals
Adult male Wistar rats (280–340 g) were kept under conditions of
constant temperature (22 ± 2 °C) with a 12 h light/12 h dark cycle and free access to food and water. All animals were cared for in compli-ance with the Guide for the Care and Use of Laboratory Animals, published by the US National Institutes of Health (NIH Publication 85-23, revised 1996). All procedures described here were reviewed by and had prior approval from local animal ethics committee (11043633-4).
2.3. Tissue preparation and experimental protocols
Rats were sacrificed by asphyxia induced by CO2inhalation. For iso-metric tension recording, thoracic aorta was removed and placed in cold oxygenated KHS buffer. Segments of this artery (3 mm in length), free of fat and connective tissue, were mounted between two steel hooks in a 5 mL isolated tissue chamber containing gassed (95% O2and 5% CO2) Krebs-Henseleit solution (KHS), at 37 °C, under a resting tension of 1 g, which was readjusted every 15 min during a 45-min equilibration
period before drug administration. Isometric tension was recorded by using an isometric force–displacement transducer (Grass Model FTO3,
Quincy, MA, USA) connected to an acquisition system (PM-1000; CWE Inc., Akron, OH, USA). Vessels were initially exposed twice to 60 mM KCl to check their functional integrity. After 30 min, rings were contracted with a concentration (0.1μM) of PHE inducing 50–70% of
the contraction induced by KCl and acetylcholine (1μM) was then added to assess endothelium integrity. Sixty min later, the series of ex-periments 1 to 8 were performed.
2.3.1. Series 1
This series of experiments was carried out to assess the effects of NPe (0.1–100μg/mL) on the resting tone of endothelium-containing aortic preparations. In other experiments, effects of vehicle at the same concentrations used to dissolve NPe were also studied.
2.3.2. Series 2
In this series of experiments, the effects of cumulative concentra-tions (0.1–100μg/mL) of NPe on the sustained contractile responses
to KCl (60 mM) or PHE (1μM) were studied in either endothelium-intact or endothelium-denuded aortic ring preparations maintained in Ca2+-containing medium. In order to assess the role of the sGC, nitric oxide (NO) synthase, prostaglandins or potassium channels in the ef-fects of NPe in PHE-contracted tissues, experiments were performed in endothelium-containing aortic ring preparations incubated for
20 min with ODQ (10μM),L-NAME (100μM), indomethacin (10μM)
or TEA (5 mM), respectively. Effects of vehicle at the same concentra-tions used to dissolve NPe were also studied.
2.3.3. Series 3
This series of experiments was carried out to assess the inhibitory ef-fects of NPe on the contractions induced by exogenous addition of Ca2+ in aortic ring preparations depolarized by KCl (60 mM) in a Ca2+-free
medium. Calcium-free solutions were prepared by omitting CaCl2
from normal KHS. After verification of tissue responsiveness in a
Ca2+-containing medium, the preparation was maintained in the
Ca2+-free KHS in the presence of 60 mM K+and EGTA (50μM) to pro-mote voltage-operated calcium channel (VOCC) activation. Thereafter, a cumulative concentration–response curve for Ca2+(0.1–20 mM) was then performed. After a washing preparation by changing the bath chamber solution to remove just Ca2+from the medium, NPe (at 1 or 3μg/mL) was added to the preparation for 5 min, and their effects on cu-mulative concentration–response curve for CaCl2(0.1–20 mM) were then evaluated. Maximal response to exogenous Ca2+was obtained when an increase in the Ca2+concentration did not induce a significant additional contraction. The contractile response obtained with thefirst concentration–response curve for CaCl2was taken as control, and the contractions were calculated as a function of the value observed to 20 mM Ca2+.
2.3.4. Series 4
The same protocol as that of series 3 (construction of cumulative
concentration–response curve for Ca2+ from 0.1 to 20 mM) was
performed in aortic ring preparations under Ca2+-free conditions. These preparations were stimulated with PHE (1μM) in the presence of verapamil (1μM) to obtain contractile responses preferentially mediated by receptor-operated calcium channels (ROCCs).
2.3.5. Series 5
To investigate whether NPe inhibitory actions could also be effective on contractions elicited by sarcoplasmic reticulum Ca2+channels acti-vated by inositol triphosphate (IP3), the effects of NPe on PHE-induced contractions in Ca2+-free medium were determined as follows. After the usual stabilization time, the tissues were washed with Ca2+-free so-lution for 6 min (with 1 mM EGTA). PHE (1μM) was added to produce a transient contraction. After washing the tissues with Ca2+-containing
KHS, 60 mM KCl was added for Ca2+internal stores reloading. The prep-arations were washed with Ca2+-free solution followed by the addition of NPe (10 or 30μg/mL) 1 min later. After 5 min, PHE (1μM) was administered still in the presence of NPe.
2.3.6. Series 6
In this series of experiments, the effects of NPe on caffeine-induced contractions in Ca2+-free medium were determined. After the usual stabilization time, the tissues were washed with Ca2+-free solution for 6 min, with the temperature maintained at 25 °C. Thereafter, caffeine (20 mM) was added in the medium and produced a transient contrac-tion. After washing the tissues with KHS, high KCl (60 mM) was added for Ca2+internal stores reloading. The preparations were washed with Ca2+-free solution and the transient contraction elicited by caf-feine was determined in the presence of NPe (3 or 10μg/mL) added 5 min before the administration of caffeine.
2.3.7. Series 7
This series of experiments was carried out to assess the inhibitory ef-fects of NPe on the contractions induced by the exogenous addition of Ca2+after depletion of intracellular Ca2+stores with thapsigargin in aor-tic ring preparations depleted by PHE (1μM) in the Ca2+-free medium. After the equilibration period, the preparation was kept in the Ca2+-free KHS (containing EGTA 100μM). Under this condition, three successive contractions were obtained with PHE (1μM) to produce depletion of the Ca2+intracellular stores. Each stimulus was followed by washing of the preparation with the Ca2+-free KHS to remove PHE from the extra-cellular medium. Thapsigargin (0.1μM) was then added to the prepara-tion to inhibit the activity of the sarcoplasmic/endoplasmic reticulum Ca2+pumps (SERCA), and thereafter a control cumulative concentra-tion–response curve for Ca2+(0.1
–20 mM) was then performed. In a separate set of aortic rings, NPe (at 1 or 3μg/mL) was added following the thapsigargin treatment and its effects on cumulative concentra-tion–response curve for CaCl2(0.1–20 mM) were then evaluated.
2.3.8. Series 8
In this series of experiments, inhibitory effects of increasing concen-trations (1–100μg/mL) of NPe on the contraction elicited by PDB (1μM)
or sodium orthovanadate (3 mM) were studied in aortic preparations with intact endothelium incubated in Ca2+-free medium with 1 mM EGTA.
2.4. Solutions and drugs
NPe was dissolved in ethanol, made up with the perfusion medium and sonicated just before use. The perfusion medium used was fresh Krebs–Henseleit solution (KHS) buffer (pH 7.4) of the following compo-sition (in mM): NaCl 118; KCl 4.7; NaHCO325; CaCl2·2H2O 2.5; KH2PO4 1.2; MgSO4·7H2O 1.2; glucose 11. PHE hydrochloride, indomethacin, ace-tylcholine chloride, ethylene glycol tetraacetic acid (EGTA), 1H-[1,2,4] oxadiazolo[4,3-a]quinoxaline-1-one (ODQ), tetraethylammonium chlo-ride (TEA), sodium orthovanadate, thapsigargin, phorbol-12,13-dibutyrate (PDB),L-N(G) nitroarginine methyl ester (L-NAME)
hydro-chloride, caffeine and verapamil were all purchased from Sigma,first dissolved in distilled water and were made up with KHS in order to achieve the desired concentration in the bath chamber (except EGTA which was added directly in the Ca2+-free KHS).
2.5. Statistical analysis
Results are expressed as mean ± S.E.M. Peak deflections were used to measure the magnitude of the concentration–response curves, which were expressed as a percentage of to a given contractile agent alone (in the absence of NPe). The IC50value, defined as the NPe concentration (μg/mL) required producing a half maximum reduction of PHE-, PDB-, sodium orthovanadate- or KCl-induced contractions was used to
evaluate vascular sensitivity to NPe. It was calculated by interpolation from semi-logarithmic plots, and was expressed as geometric mean [95% confidence interval]. The significance (Pb0.05) of results was
assessed by a paired Student'st-test, Mann–Whitney U-test, and one-or two-way analysis of variance (ANOVA), followed by Dunnett's multi-ple comparison test when appropriate.
3. Results
3.1. Effects of NPe on the resting tonus (Series 1)
In endothelium-containing aortic ring preparations, neither vehicle (n = 6) nor NPe (0.1–100μg/mL, n = 6) had significant (PN0.05,
two-way ANOVA) effects on the resting tone (data not shown).
3.2. Relaxant effects of NPe on the sustained contraction induced by KCl or PHE (Series 2)
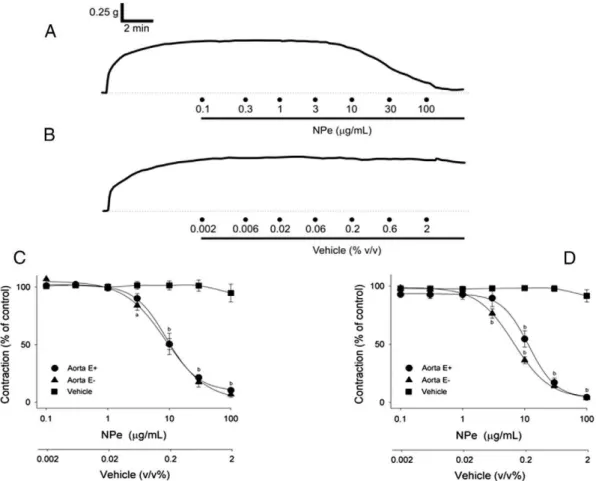
At 0.1–100μg/mL, NPe evoked a significant and concentration-dependent (Fig. 2A, C Pb0.001, one-way ANOVA) relaxation of the
endothelium-containing aortic ring preparations (n = 7) pre-contracted with 60 mM KCl with an IC50value which was not signifi -cantly (Table 1;PN0.05, Mann–Whitney U-test) different from that
ob-tained using endothelium-denuded preparations (n = 6). In both preparations, relaxant effects of NPe became significant at a concentra-tion of 10μg/mL (Fig. 2C Pb0.05, one-way ANOVA and Dunnett's
tests).
In both endothelium-containing (n = 7) and endothelium-denuded (n = 6) aortic ring preparations pre-contracted with PHE (1μM), NPe (0.1–100μg/mL) also induced a similar (PN0.05, Mann–Whitney
U-test) and concentration-dependent (Fig. 2D;P b0.001, one-way
ANOVA) relaxation with IC50values of the same order of magnitude as those obtained for relaxant K+-induced contractions (Table 1;P
N0.05,
Mann–Whitney U-test). Thefirst inhibitory effect of NPe became signifi -cant (Pb0.05, one-way ANOVA and Dunnett's tests) at 10 and 3μg/mL
for NPe-induced relaxant in intact and endothelium-denuded preparations, respectively (Fig. 2D). Maximal smooth muscle-relaxing activity of NPe against PHE-elicited contractions occurred at a concentration of 100μg/mL (Fig. 2D), as was observed for the inhibitory effects of NPe on K+-induced contractions (Fig. 2C).
The values for the pre-contraction induced by KCl and PHE in aortic rings with intact endothelium were 0.879 ± 0.053 and 0.823 ± 0.088 g, respectively while those in endothelium-denuded preparations were 1.237 ± 0.143 and 1.277 ± 0.098 g, respectively. Furthermore, in a sep-arate set of experiments, vehicle added cumulatively at the same con-centration range used to dissolve NPe did not affect significantly (PN0.05, one-way ANOVA) the KCl- (Fig. 2B, C) and PHE-induced
(Fig. 2D) contractions (n = 5 and n = 5, respectively).
In order to assess the role of K+channels in these effects, experi-ments have been performed in PHE-contracted endothelium-intact aor-tic ring preparations incubated for 20 min with TEA, a nonspecific K+ channel blocker. Under these conditions, NPe (0.1–100μg/mL, n = 5) re-laxed in a concentration-dependent manner the contractions evoked by PHE, with an IC50value that did not differ significantly (Table 1;PN0.05,
Mann–Whitney U-test) from that obtained in the absence of TEA (Fig. 3).
Vasorelaxant effects of NPe (0.1–100μg/mL) remained also significantly
unchanged (Fig. 3 and Table 1; P N0.05, two-way ANOVA) in
endothelium-containing aortic ring preparations incubated for 20 min withL-NAME (n = 5), indomethacin (n = 10) or ODQ (n = 9).
3.3. Inhibitory effects of NPe on Ca2+-induced contractions in KCl-stimulated preparations (Series 3)
In aortic preparations with intact endothelium incubated in Ca2+-free medium in the presence of high KCl solution, increasing concentrations of
contractions (Pb0.001, one-way ANOVA), an effect that became signifi
-cant at 0.1 mM (Pb0.05, one-way ANOVA and Dunnett's test) and
attained a maximal magnitude at 10 mM (Fig. 4A). Contractions due to Ca2+addition were signi
ficantly (Pb0.01, two-way ANOVA) reduced
or even abolished by NPe at 1 (n = 6) and 3 (n = 7)μg/mL, respectively (Fig. 4A,Pb0.01, two-way ANOVA).
3.4. Inhibitory effects of NPe on Ca2 +-induced contractions in PHE-stimulated preparations in the presence of verapamil (Series 4)
In aortic preparations with intact endothelium incubated in Ca2+-free
medium in the presence of PHE (1μM) and verapamil (1μM), the
addition of increasing concentrations of CaCl2(0.1–20 mM, n = 7) also
evoked concentration-dependent contractions (P b0.001, one-way
ANOVA), an effect that became significant at 0.1 mM (Pb0.05, one-way
ANOVA and Dunnett's tests) and attained a maximal magnitude at 10 mM (Fig. 4B). Contractions due to Ca2+addition were abolished by NPe at 3μg/mL (Fig. 4B;P b0.01, two-way ANOVA, n = 7) while
remained unaffected by NPe at 1μg/mL (Fig. 4B;PN0.05, two-way
ANOVA, n = 7).
Fig. 2.Typical trace recordings of the effect of 1-nitro-2-phenylethene (NPe, 0.1–100μg/mL) (A) and its vehicle (B) on the sustained contractions induced by 60 mM KCl in isolated aortic preparations from normotensive rats. Concentration–effect curves for the relaxant effects of NPe (0.1–100μg/mL) in endothelium-intact (E+) and -denuded (E−) aortic rings
pre-contracted with KCl (60 mM) (C) or phenylephrine (PHE, 1μM) (D). In panels C and D, note that vehicle added cumulatively at the same concentration (% v/v) range used to dissolve
NPe did not affect significantly the KCl- and PHE-induced contractions, respectively. Results are expressed as means ± SEM (n = 5–7 per group).aP
b0.05,bPb0.001 by two-way ANOVA followed by Dunnett's test. Calibrations: vertical 0.25 g, horizontal 2 min.
Table 1
IC50values for the vasorelaxant effects of 1-nitro-2-phenylethene in aortic ring
prepara-tions pre-contracted with 60 mM KCl or with 1μM phenylephrine subjected to various pre-treatments. Values are expressed as geometric mean [95% confidence interval] and number within parentheses indicates the number of preparations for each group.
Contractile agent/pre-treatments IC50(μg/mL) n
Phenylephrine
Control 10.47 [4.68–16.26] (7) Endothelium removal 6.71 [5.27–8.39] (6)
+TEA 10.59 [7.19–13.98] (5)
+L-NAME 12.83 [2.45–23.22] (5)
+ODQ 12.03 [8.86–15.37] (9)
+Indomethacin 8.42 [5.48–11.36] (10)
KCl
Control 10.59 [7.19–13.98] (7) Endothelium removal 10.58 [4.66–16.49] (6) Control: with intact endothelium.
Fig. 3.Effects of the pretreatment with tetraethylammonium (TEA, 5 mM),L-NAME
3.5. Effects of NPe on PHE-induced contractions in Ca2+-free medium (Series 5)
Under Ca2+-free conditions (with 1 mM EGTA), PHE (1μM) induced a transient contraction which corresponded to 37.18 ± 4.23% of the K+ -induced response under nominally normal Ca2+. Pre-exposure of aortic ring preparations for 5 min to 10 (n = 8) or 30 (n = 12)μg/mL of NPe significantly (P b0.05, paired Student's t-test) reduced or even
abolished respectively the phasic contraction induced by PHE to 12.11 ± 2.11 and 2.45 ± 0.66%, respectively (Fig. 5A).
3.6. Effects of NPe on caffeine-induced contraction in Ca2+-free medium (Series 6)
In this series of experiments, caffeine was used as a pharmacological tool to assess whether NPe is able to inhibit a contraction induced by the release of Ca2+-releasable Ca2+from the sarcoplasmatic reticulum stores. In aortic ring preparations with intact endothelium maintained under Ca2+-free conditions, caffeine-induced contraction remained sig-nificantly (P N0.05, paired Student's t-test) unaltered by either 3
(14.12 ± 3.61 vs. 11.16 ± 2.22% of the contraction induced by
60 mM K+in the absence of NPe, n = 5) or 10 (13.26 ± 5.13 vs.
13.26 ± 5.13% of the contraction induced by 60 mM K+in the absence of NPe, n = 5)μg/mL of NPe (Fig. 5B).
3.7. Effects of NPe on contractions induced in aortic rings after depletion of intracellular Ca2+stores (Series 7)
In aortic rings with intact endothelium submitted to depletion of in-tracellular Ca2+stores by PHE followed by thapsigargin (in the presence of 100μM EGTA), the addition of increasing concentrations of CaCl2 (0.1–20 mM, n = 7) evoked concentration-dependent contractions
(P b0.001, one-way ANOVA) that became significant at 3 mM
Fig. 4.Inhibitory effects of 1-nitro-2-phenylethene (NPe, 1 and 3μg/mL) on the cumula-tive concentration–effect curve dependent on extracellular Ca2+addition (0.1–20 mM)
in endothelium-intact, KCl-stimulated (A) or phenylephrine-stimulated (B) aortic ring preparations from normotensive rats incubated in the Ca2+-free medium. Results are
expressed as means ± SEM (n = 5–11 per group); *first significant effect for control curve or for curves obtained in the presence of NPe (Pb0.05, one-way ANOVA followed by Dunnett's test);#P
b0.01 by two-way ANOVA, NPe vs. concentration–contraction curve of Ca2+alone (control).
Fig. 5.Inhibitory effects of 1-nitro-2-phenylethene (NPe) (3–30μg/mL) on the contraction evoked by phenylephrine (1μM) (A) or caffeine (20 mM) (B) in aortic ring preparations with intact endothelium maintained under Ca2+-free conditions. Results are expressed
as means ± SEM (n = 5–12 per group); *Pb0.01 by paired Student'st-test vs. control.
Fig. 6.Inhibitory effects of 1-nitro-2-phenylethene (NPe, 3 and 10μg/mL) on the
cumula-tive concentration–effect curve dependent on extracellular Ca2+addition (0.1–20 mM) in
endothelium-intact phenylephrine-depolarized aortic ring preparations from normoten-sive rats pretreated with thapsigargin, and incubated in the Ca2+-free medium. Results
(Pb0.05, one-way ANOVA and Dunnett's tests) and attained a maximal
magnitude at 10 mM (Fig. 6). Contractions due to Ca2+addition were not altered by NPe at 3μg/mL (n = 7) but were abolished (Pb0.001,
two-way ANOVA) by NPe at 10μg/mL (Fig. 6, n = 7).
3.8. Effects of NPe on the sustained contraction induced by PDB or sodium orthovanadate under Ca2+-free conditions (Series 8)
In aortic ring preparations incubated in Ca2+-free medium (with
1 mM EGTA), the PKC activator PDB (1μM) induced a long-lasting
and sustained contraction corresponding to 125.60 ± 11.44% of the K+-induced contraction. When the contraction reached steady state, cu-mulative addition of increasing concentrations (1–100μg/mL) of NPe (Fig. 7A, n = 6) significantly reduced the PDB-induced contractions in a concentration-dependent manner (Pb0.001, one-way ANOVA). The
effect of NPe became significant at 3μg/mL (Fig. 6A;Pb0.01, one-way
ANOVA and Dunnett's tests) with an IC50value (15.61 [10.13–21.10] μg/mL) which was of the same order of magnitude (PN0.05, Mann–
Whitney U-test) as that evoked by NPe against the contractions elicited by PHE.
The addition of increasing concentrations (1–100μg/mL, n = 5) of
NPe also reduced significantly the contractions evoked by sodium orthovanadate (3 mM) under Ca2+-free conditions (in the presence of 1 mM EGTA) reaching thefirst significant inhibitory effect at 3μg/mL (Fig. 7B;Pb0.05, one-way ANOVA and Dunnett's tests) and an IC50 value (13.91 [9.44–37.25]μg/mL) similar to that evoked by NPe against the contractions induced by PDB.
4. Discussion
This study demonstrated that NPe has interesting vasorelaxant properties that are independent of the integrity of endothelium. NPe inhibited with the same potency either the electromechanical or pharmacomechanical couplings as well as the contractions induced by PDB or sodium orthovanadate under Ca2+-free conditions, suggesting that its vasorelaxation is of myogenic nature probably through inhibi-tion of contracinhibi-tions that are independent of Ca2+in
flux from the extracellular milieu.
Previously, we showed that 1-nitro-2-phenylethane fully relaxed endothelium-intact aortic ring preparations pre-contracted with PHE in a concentration-dependent manner with an IC50value of 37.65 [23.11–52.2]μg/mL corresponding to 231.5 [154.1–348.0]μM[14]. In the present study, NPe induced its vasorelaxant effect with an IC50 value of 10.47 [4.68–16.26]μg/mL corresponding to 70.74 [31.62–
109.86]μM. It is reasonable to hypothesize that the conformational restriction imposed to NPe molecule by the substitution of the alkane for the alkene moiety is involved in the 3.3-fold increased potency of NPe to relax aortic ring preparations, probably because such change im-pairs the expected rotationalflexibility of the aliphatic side-chain of the 1-nitro-2-phenylethane molecule. Recently, conformational analysis of 1-nitro-2-phenylethane showed that this molecule can adopt different conformations varying dihedral angles between phenyl and nitro groups linked at ethylene carbon atoms[18]. The arrangement related to the mains-transconformer with the dihedral angle of the 180° for O-2N\C1\C2\C6H5was significantly more stable than thes-cis conform-er (angle 0°). Othconform-er two conformations of low enconform-ergies such as 60° and 120°, which may constitute one of the factors responsible for the loss of selectively of a drug and/or of interactions with other targets[19], were also observed[18]. It noteworthy that NPe has only the 180° conforma-tion structure.
Putative participation of the vascular endothelium in modulating NPe-induced relaxation has been investigated in the current study. Our results show that the smooth muscle-relaxing activity of NPe in aor-tic preparations is independent on the integrity of endothelial layer as it remained unaltered by vascular endothelium removal. This was evi-denced by the non-significant differences between IC50values obtained in endothelium-containing and those in endothelium-denuded aortic preparations pre-contracted with PHE. Notwithstanding, since maximal efficacy for the relaxant effect induced by NPe was not changed by en-dothelium removal, it is reasonable to postulate that its myorelaxant effects have a nature predominantly myogenic. The hypothesis of a myogenic action is reinforced by the results obtained with ring prepara-tions depolarized with high K+concentrations. In fact, NPe relaxed preparations maintained in the presence of 60 mM [K+], suggesting that it might also interfere with contractions induced by electrome-chanical coupling. Such afinding allows us to conclude that: (i) the re-laxant effects of NPe could not be explained by an intramural neuron influence on vascular smooth cell layers because so high [K+]
o depolar-izes cell membrane to approximately−20 mV[20], a value that impairs
neuronal action potential by slowing Na+channel recovery from
inactivation[21], (ii) the inhibitory actions of NPe also could not be at-tributed exclusively to a putative ability of this substance in producing K+-channel opening because when smooth muscle cell is depolarized by high K+concentrations, the resulting transmembrane potential is turned very close to the Nernst's K+equilibrium potential[22]. Under such despolarizing conditions, it is expected that a K+-channel opener is useless to displace the transmembrane potential as a result of its ability to opening K+channels. This conclusion was therefore supported by thefinding that pre-incubation of aortic preparations with the non-selective potassium channel blocker TEA was unable to alter signifi cant-ly the vasorelaxant effects of NPe. Thus, in conjunction with the results obtained in endothelium-denuded aortic preparations, it is improbable that relaxation of KCl-induced contraction may be due to a direct or even indirect (e.g. by means of endothelium-derived hyperpolarizing Fig. 7.Effects of cumulative and increasing concentrations (1–100μg/mL) of
1-nitro-2-phenylethene (NPe) on the sustained contractions induced by phorbol 12,13-dibutyrate (PDB) (1μM) (A) or by sodium orthovanadate (3 mM) (B) in aortic ring preparations from normotensive rats maintained under Ca2+-free conditions. Results are expressed
factor-EDHF release) hyperpolarization caused by NPe. Furthermore, neither endothelial NO release nor activation of sGC is involved in the vasorelaxant effect of NPe as it remained unchanged following pre-incubation withL-NAME and ODQ, respectively. On the other hand,
prostaglandin release is also not involved as pretreatment with indo-methacin, a non-selective cyclooxygenase inhibitor, has no significant effect on the NPe-induced vasorelaxation. Although the relative de-gree of pre-constriction as a % of the maximum as this has the poten-tial to have a major influence on relaxation responses, it should be highlighted that the vasorelaxant effects of NPe were not influenced by the higher magnitude of the PHE-induced contractions in
the presence of L-NAME or ODQ, since the contractile response
was fully reversed by NPe without any change in its IC50value. For that reason, a titration of PHE for each vascular segment was unnecessary.
Regarding the excitation–contraction coupling in vascular smooth
muscle, it is well known that high KCl induces membrane depolariza-tion, which, in turn, opens the VOCCs, promotes Ca2+in
flux, increases [Ca2+]
i, and elicits sustained contraction following myosin light chain phosphorylation[23]. This study shows that NPe was able to inhibit the electromechanically mediated KCl-induced contractions and it also reduced or even abolished the contractions induced by CaCl2in aortic preparations depolarized with high KCl in Ca2+-free medium. Under these conditions, the contractions induced by CaCl2are due to an in-crease in Ca2+in
flux through VOCCs. In addition, we also showed that NPe was able to interfere with pharmacomechanical coupling. It is well-known that PHE contracts vascular smooth muscle as a result of theα1-adrenoceptor causing an initial phase of contraction elicited by IP3-induced intracellular Ca2+release from sarcoplasmic reticulum, followed by a second sustained phase that develops slowly and depends upon dihydropyridine-insensitive Ca2+in
flux through ROCCs. In the presence of verapamil to remove the indirect influence of VOCC-mediated Ca2+in
flux, NPe reduced the CaCl2-induced contractions in aortic preparations pretreated with PHE under Ca2+-free conditions. Thisfinding, together with the ability of NPe to reduce PHE-induced contractions, suggests that NPe has an antispasmodic activity against contractile events mediated by Ca2+entry through ROCCs. It is note-worthy, however, that thefinding that NPe relaxed contractions evoked pharmacomechanically or electromechanically with the same potency reinforces the idea of a myogenic action as the underlying mechanism by which NPe induces its vasodilator effect.
Therefore, the effects of NPe on the contractions induced by intracel-lular events, such as Ca2+release from the internal stores located in sar-coplasmic reticulum were addressed in the present study and experiments were performed in the Ca2+-free KHS. Under such condi-tions, transient tension development activated by a PHE-induced stim-ulus is expected to result from Ca2+released from sarcoplasmic reticulum stores upon activation of IP3-sensitive Ca2+channels[24]. Our results showed that these transient contractions were also reduced by NPe (10μg/mL). In contrast, aortic preparations in Ca2+-free solu-tion suffered no change in the amplitude of the caffeine-elicited tran-sient contractions after treatment with the same concentration of NPe, revealing that while NPe is effectively inhibitory on contractions induced by the increased intracellular availability of Ca2+caused by IP3, it does not interfere with contractions induced by Ca2+-induced Ca2+release from the sarcoplasmic reticulum via ryanodine receptors
[25].
We also investigated whether NPe was able to inhibit contractions evoked by the influx of Ca2+through store-operated Ca2+channels (SOCCs)[26]. It is known that SOCCs are activated by depletion of the Ca2+stores within the sarcoplasmic reticulum allowing to a capacitative Ca2+influx into the cytosol and a sustained contraction which may occur
directly or indirectly through membrane depolarization [27].
Thapsigargin is a pharmacological tool used to deplete intracellular Ca2+stores since it inhibits the Ca2+-ATPase pump (SERCA) localized at the sarcoplasmic reticulum membrane[28]. Ourfindings showed
that when aortic preparations were repeatedly depleted by PHE followed by treatment with thapsigargin, the exogenous addition of Ca2+evoked sustained contractions that were abolished by NPe, as was previously reported for 1-nitro-2-phenylethane[14].
In pharmacomechanical coupling, it is well-known that receptor ag-onists cause smooth muscle contraction by increasing Ca2+influx, [Ca2+]
iand by causing Ca2+sensitization[29]. Activated receptors can trigger additionally Ca2+sensitizing mechanisms involving PKC that are independent of Ca2+in
flow from the extracellular environment
[30]. In order to evaluate the effect of NPe on Ca2+sensitivity compo-nent of contraction, precisely on PKC signaling pathway, we used PDB, a potent-specific PKC activator[31]. PDB induced a sustained contrac-tion of endothelium-containing aortic preparacontrac-tions incubated in the Ca2+-free medium with EGTA. This contractile effect was reversed after treatment with NPe, indicating that this substance may interfere with cellular events that increase PKC-mediated Ca2+sensitivity to con-tractile proteins. In this direction, it has been also shown that phosphor-ylation of tyrosine by tyrosine kinases is able to activate many intracellular signaling pathways, resulting in various cellular events, in-cluding the contraction of vascular smooth muscle[32]. The level of phosphorylated tyrosine residues is regulated by a balance between the actions of tyrosine kinases and tyrosine phosphatases. The sodium orthovanadate, a potent protein tyrosine phosphatase inhibitor, induces smooth muscle contraction due to a reduction of dephosphorylated ty-rosine residues, thus increasing indirectly the amount of phosphorylat-ed tyrosine[33]. In aortic preparations with intact endothelium in the medium without Ca2+, NPe was able to relax the contractions induced by sodium orthovanadate in a concentration-dependent manner with a potency that was similar to that evoked by NPe against PBD-induced contractions. This suggests that vasorelaxant effects of NPe are partly dependent on the amount of phospholyrated tyrosine residues. Further studies with isolated vascular smooth cells will be required to study detailed Ca2+signaling mechanisms. On the other hand, experiments using rat isolated mesenteric artery to assess the vasorelaxant effects of NPe are underway in our laboratory. As was seen with 1-nitro-2-phenylethane[14], preliminary results showed higher potency of NPe in these vessels. Although we are unable to presently explain this finding, such a feature suggests a preferential action of this compound in tissues that are more involved in hemodynamic control and ultimately determine peripheral vascular resistance.
5. Conclusions
In conclusion, ourfindings indicate that NPe induced endothelium-independent vasorelaxation, an effect that appears to occur intracel-lularly probably through inhibition with a similar potency of con-tractions clearly independent of Ca2 +in
flux from the extracellular milieu. Because of the conformational restriction of its alkene moie-ty, NPe was shown to be nearly 3.5 times more potent than its struc-tural analog 1-nitro-2-phenylethane to induce vasorelaxation. Thus, NPe can be used as scaffold for the synthesis of more potent novel de-rivatives which could be promising candidates as antihypertensive drugs.
Acknowledgments
This work was supported by the“Conselho Nacional de Pesquisa”
(CNPq), the“Coordenação de Aperfeiçoamento de Pessoal de Nível
Supe-rior”(CAPES) and the“Fundação Cearense de Apoio ao Desenvolvimento
References
[1]Lahlou S, Leal-Cardoso JH, Magalhães PJ. Essential oil ofCroton nepetaefolius decreases blood pressure through an action upon vascular smooth muscle: studies in DOCA-salt hypertensive rats. Planta Med 2000;66:138–43.
[2]Lahlou S, Magalhães PJ, Carneiro-Leão RF, Leal-Cardoso JH. Involvement of nitric oxide in the mediation of the hypotensive action of the essential oil ofMentha ×villosain normotensive conscious rats. Planta Med 2002;68:694–9.
[3]de Siqueira RJ, Magalhães PJ, Leal-Cardoso JH, Duarte GP, Lahlou S. Cardiovascular effects of the essential oil ofCroton zehntnerileaves and its main constituents, anethole and estragole, in normotensive conscious rats. Life Sci 2006;78:2365–72.
[4]Interaminense LF, Jucá DM, Magalhães PJ, Leal-Cardoso JH, Duarte GP, Lahlou S. Pharmacological evidence of calcium-channel blockade by essential oil ofOcimum gratissimumand its main constituent, eugenol, in isolated aortic rings from DOCA-salt hypertensive rats. Fundam Clin Pharmacol 2007;21:497–506.
[5]Pinto NV, Assreuy AM, Coelho-de-Souza AN, Ceccatto VM, Magalhães PJ, Lahlou S, et al. Endothelium-dependent vasorelaxant effects of the essential oil from aerial parts ofAlpinia zerumbetand its main constituent 1,8-cineole in rats. Phytomedicine 2009;16:1151–5.
[6]Magalhães PJ, Lahlou S, Jucá DM, Coelho-de-Souza LN, da Frota PT, da Costa AM, et al. Vasorelaxation induced by the essential oil ofCroton nepetaefoliusand its constitu-ents in rat aorta are partially mediated by the endothelium. Fundam Clin Pharmacol 2008;22:169–77.
[7]Lahlou S, de Barros Correia Jr CA, Vasconcelos dos Santos M, David JM, David JP, Duarte GP, et al. Mechanisms underlying the cardiovascular effects of a labdenic diterpene isolated from Moldenhawera nutansin normotensive rats. Vascul Pharmacol 2007;46:60–6.
[8]Peixoto-Neves D, Silva-Alves KS, Gomes MD, Lima FC, Lahlou S, Magalhães PJ, et al. Vasorelaxant effects of the monoterpenic phenol isomers, carvacrol and thymol, on rat isolated aorta. Fundam Clin Pharmacol 2010;24:341–50.
[9]Gottlieb OR, Magalhães MT. Essential oil of bark and wood ofAniba canelilla. Perfum-ery Essent Oil Rec 1960;50:69–70.
[10]Gottlieb OR. Chemosystematics on the Lauraceae. Phytochemistry 1972;5:1537–70.
[11]de Siqueira RJ, Macedo FI, Interaminense LF, Duarte GP, Magalhães PJ, Brito TS, et al. 1-Nitro-2-phenylethane, the main constituent of the essential oil ofAniba canelilla, elicits a vago-vagal bradycardiac and depressor reflex in normotensive rats. Eur J Pharmacol 2010;638:90–8.
[12]Interaminense LF, de Siqueira RJ, Xavier FE, Duarte GP, Magalhães PJ, da Silva JK, et al. Cardiovascular effects of 1-nitro-2-phenylethane, the main constituent of the essential oil ofAniba canelilla, in spontaneously hypertensive rats. Fundam Clin Pharmacol 2011;25:661–9.
[13]Interaminense LF, dos Ramos-Alves FE, de Siqueira RJ, Xavier FE, Duarte GP, Magalhães PJ, et al. Vasorelaxant effects of 1-nitro-2-phenylethane, the main constituent of the essential oil ofAniba canelilla, in superior mesenteric arteries from spontaneously hypertensive rats. Eur J Pharm Sci 2013;48:709–16.
[14]Brito TS, Lima FJ, Aragão KS, de Siqueira RJ, Sousa PJ, Maia JG, et al. The vasorelaxant effects of 1-nitro-2-phenylethane involve stimulation of the soluble guanylate cyclase–cGMP pathway. Biochem Pharmacol 2013;85:780–8.
[15]Vogel AI. Vogel's textbook of practical organic chemistry. [revised by Furniss, B.S., Hannaford, A.J., Smith, P.W.G., Tatchell, A.R.] 5th ed. New York: Longman Scientific and Technical; 1989.
[16]Ford PW, Narbut MR, Belli J, Davidson BS. Synthesis and structural properties of the benzopentathiepins varacin and isolissoclinotoxin A. J Org Chem 1994;59:5955–60.
[17]Wang C, Wang S. The rapid synthesis of β-nitrostyrenes under microwave irradiation without solvent. Synth Commun 2002;32:3481–6.
[18]Vale JK, Lima AB, Pinheiro BG, Cardoso AS, Silva JK, Maia JG, et al. Evaluation and the-oretical study on the anti-inflammatory mechanism of the 1-nitro-2-phenylethane. Planta Med 2013;79:628–33.
[19]Tarcsay Á, KeserűGM. Contributions of molecular properties to drug promiscuity. J Med Chem 2013;56:1789–95.
[20]Grafe P, Mayer CJ, Wood JD. Synaptic modulation of calcium-dependent potassium conductance in myenteric neurones in the guinea-pig. J Physiol 1960;305:235–48.
[21]Meeks JP, Mennerick S. Selective effects of potassium elevations on glutamate signaling and action potential conduction in hippocampus. J Neurosci 2004;24: 197–206.
[22]Hodgkin AL, Horowicz P. The influence of potassium and chloride ions on the membrane potential of single musclefibres. J Physiol 1959;148:127–60.
[23]Somlyo AV, Somlyo AP. Electromechanical and pharmacomechanical coupling in vascular smooth muscle. J Pharmacol Exp Ther 1968;159:129–45.
[24]Islam MO, Yoshida Y, Koga T, Kojima M, Kangawa K, Imai S. Isolation and character-ization of vascular smooth muscle inositol 1,4,5,-trisphosphate receptor. Biochem J 1996;316:295–302.
[25]Karaki H, Weiss GB. Calcium release in smooth muscle. Life Sci 1988;44:111–22.
[26]Putney Jr JW. Type 3 inositol 1,4,5-trisphosphate receptor and capacitative calcium entry. Cell Calcium 1997;21:257–61.
[27]Clapham DE, Runnels LW, Strübing C. The TRP ion channel family. Nat Rev Neurosci 2001;2:387–96.
[28]Thastrup O, Cullen PJ, Drobak BK, Hanley MR, Dawson AP. Thapsigargin, a tumor promoter, discharges intracellular Ca2+stores by specific inhibition of the endoplas-mic reticulum Ca2+-ATPase. Proc Natl Acad Sci U S A 1990;87:2466–70.
[29]Somlyo AP, Wu X, Walker LA, Somlyo AV. Pharmacomechanical coupling: the role of calcium, G-proteins, kinases and phosphatases. Rev Physiol Biochem Pharmacol 1999;134:201–34.
[30]Kitazawa T, Eto M, Woodsome TP, Brautigan DL. Agonists trigger G protein-mediated activation of the CPI-17 inhibitor phosphoprotein of myosin light chain phosphatase to enhance vascular smooth muscle contractility. J Biol Chem 2000; 275:9897–900.
[31]Yanagita T, Kobayashi H, Yamamoto R, Takami Y, Yokoo H, Yuhi T, et al. Protein ki-nase C and the opposite regulation of sodium channel alpha- and beta1-subunit mRNA levels in adrenal chromaffin cells. J Neurochem 1999;73:1749–57.
[32]Yu J, Ogawa K, Tokinaga Y, Mizumoto K, Kakutani T, Hatano Y. The inhibitory effects of isoflurane on protein tyrosine phosphorylation-modulated contraction of rat aortic smooth muscle. Anesthesiology 2004;101:1325–31.