BR-101 Norte Km 60, Bairro Litorâneo, 29932-540 São Mateus-ES, Brazil. ffsallesgmail.com
Texto
Imagem



Documentos relacionados
The WMO used a criterion that assesses the number of stations per km 2 considering population density. The analysis of the rainfall monitoring network was conducted in order to
The pattern recognition method (principal component analysis PCA and cluster analysis CA and discriminant analysis methods) have been employed to obtain the relationship between
The main purpose of time series analysis methods is to define the variability of the data in terms of dominant periodic functions, or pattern recognition. In order to apply
The data analysis was conducted in the following order: analysis of the risk of bias; assessment of heterogeneity (statistical and clinical); analysis of the quantitative
DNA gel blot analysis was carried out to study the distribution of MULE transposases related to MoTUs in Triticum and Aegilops species and the distribution of ho- mologous
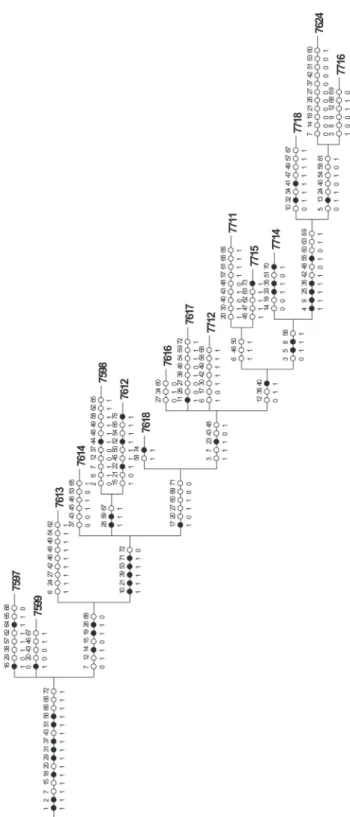
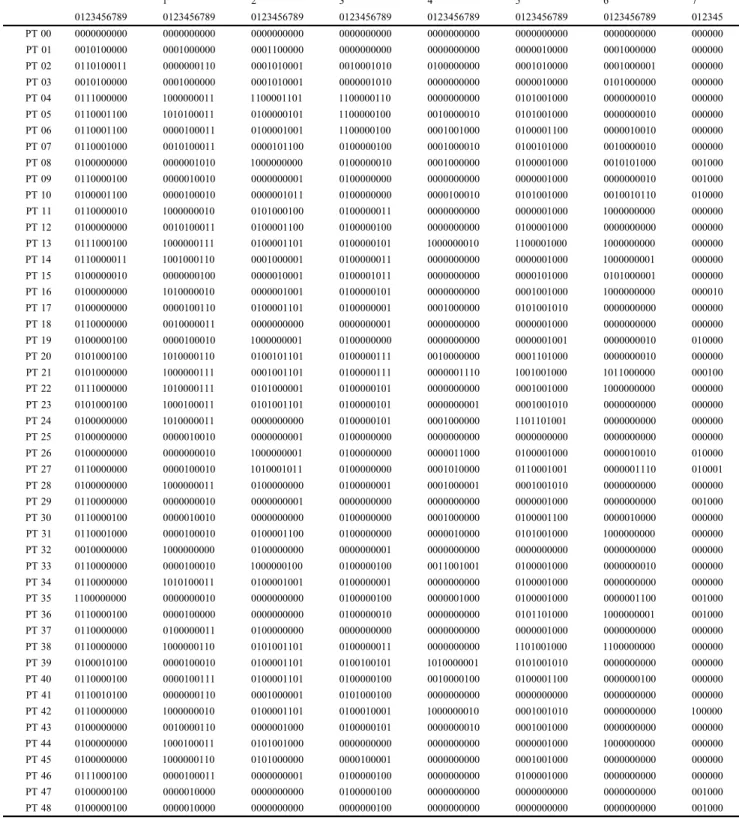
According to Ayres & Clutton-Brock (1992), the main rivers may represent natural geographic barriers for some primates. Interfluvial OGUs were delimited by: I)
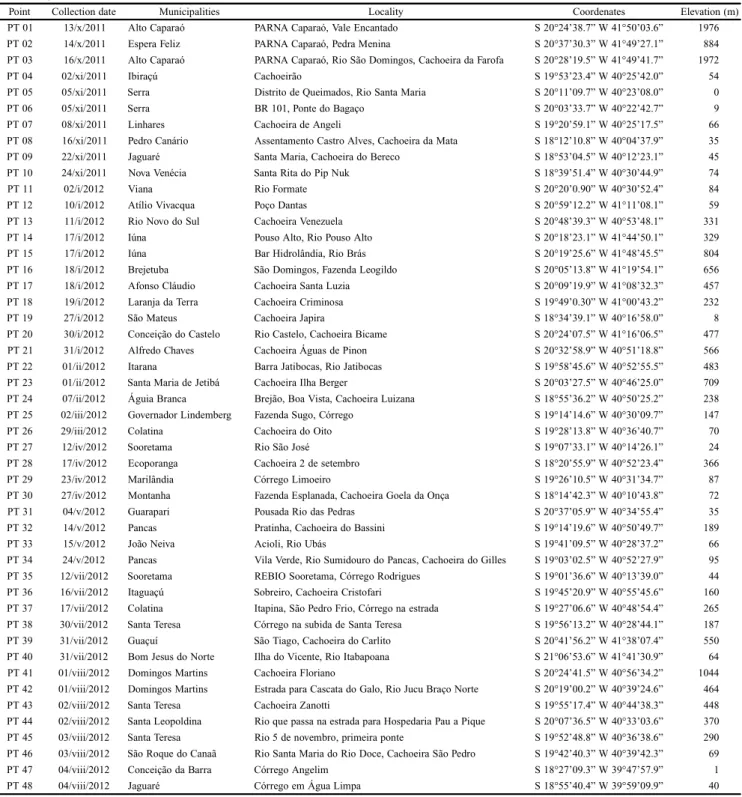
Department of Agrarian and Biological Sciences, Federal University of Espírito Santo, University Center of Northern Espírito Santo, Rodovia BR 101 Norte, Km 60, Bairro
It was observed that the pericycle was responsible for the formation of the vascular system of the rhizome and of the adventitious roots; the endodermis gave rise
