Estudo de marcadores genéticos com interesse forense e de
ancestralidade em três populações de imigrantes de PALOP
residentes em Lisboa
Dissertação de Mestrado em Genética Molecular Comparativa e Tecnológica
Ana Rita Dias Inácio
Orientadores:
António Amorim, Especialista Superior de Medicina Legal, Instituto Nacional de Medicina Legal e Ciências Forenses, Delegação do Sul, Serviço de Genética e Biologia Forenses
Gilberto Igrejas, Professor Associado com Agregação, Universidade de Trás-os-Montes e Alto Douro, Departamento de Genética e Biotecnologia
Estudo de marcadores genéticos com interesse forense e de
ancestralidade em três populações de imigrantes de PALOP
residentes em Lisboa
Dissertação de Mestrado em Genética Molecular Comparativa e Tecnológica
Ana Rita Dias Inácio
Orientadores:
António Amorim, Especialista Superior de Medicina Legal, Instituto Nacional de Medicina Legal e Ciências Forenses, Delegação do Sul, Serviço de Genética e Biologia Forenses
Gilberto Igrejas, Professor Associado com Agregação, Universidade de Trás-os-Montes e Alto Douro, Departamento de Genética e Biotecnologia
Composição do Júri: Presidente
Fernanda Maria Leal Madaleno Rei Tomás Santos - Professora Auxiliar da UTAD/ECVA Vogais
Paula Filomena Martins Lopes - Professora Auxiliar da UTAD
Estela Maria Bastos Martins de Almeida - Professora Auxiliar da UTAD
António Amorim - Especialista Sup. de Medicina Legal, Professor Auxiliar Convidado FCUL
Orientadores:
___________________________________________________________________________ (António Amorim, Especialista Superior de Medicina Legal, Instituto Nacional de Medicina Legal e Ciências
Forenses, Delegação do Sul, Serviço de Genética e Biologia Forenses)
___________________________________________________________________________ (Gilberto Igrejas, Professor Associado com Agregação, Universidade de Trás-os-Montes e Alto Douro,
Após o término desta etapa não poderia deixar de agradecer a quem caminhou a meu lado durante a mesma e que me incentivou na concretização da mesma.
À Universidade de Trás-os-Montes e Alto Douro, principalmente à coordenação do Mestrado em Genética Molecular Comparativa e Tecnológica.
Ao meu orientador Professor António Amorim, por ter aceite a orientação da presente Dissertação de Mestrado, pela sua disponibilidade, por todos os conhecimentos transmitidos, por toda a paciência que teve comigo, pela sua amizade e pelos inúmeros incentivos prestados ao longo da realização do presente trabalho.
Ao meu co-orientador Professor Gilberto Igrejas, por ter aceite a co-orientação da presente Dissertação de Mestrado, pela sua prontidão e disponibilidade, especialmente com a sua ajuda nas correções dos trabalhos realizados no decorrer da presente etapa e nos seus incentivos.
Ao Sr. Professor Jorge Costa Santos e ao Dr. Mário Dias, nas condições de antigos diretores da Delegação do Sul do Instituto Nacional de Medicina Legal e Ciências Forenses, por permitiram a realização do presente trabalho nas instalações do Serviço de Genética e Biologia Forenses da Delegação do Sul do Instituto Nacional de Medicina Legal e Ciências Forenses.
À Dra. Maria João Porto por me ter aceite e por ter permitido a realização do projeto no Serviço de Genética e Biologia Forenses e à Dra. Teresa Ribeiro por me ter recebido e integrado no Serviço de Genética e Biologia Forenses da Delegação do Sul do Instituto Nacional de Medicina Legal e Ciências Forenses.
À Dra. Heloísa Afonso Costa por todas as palavras de encorajamento, pelos desabafos nas horas de maior “desespero” e pelos conhecimentos transmitidos ao longo desta etapa.
À Dra. Cláudia Vieira da Silva pela sua prontidão e pelas suas palavras-chave de incentivo quando o trabalho não estava a correr da melhor forma.
À restante equipa do Serviço de Genética e Biologia Forenses da Delegação do Sul, pela ajuda técnica prestada.
À Cathy Paulino pela amizade e pela forma como me encorajou neste processo, fazendo-me acreditar que era capaz e que tudo iria ser possível, colocando sempre um sorriso na minha cara.
conselheiras. Aos meus restantes colegas estagiários que passaram pelo nosso Serviço de Genética e Biologia Forenses ao longo desta fase, que me proporcionaram bons momentos de descontração e que me possibilitaram transmitir-lhes os meus conhecimentos.
Ao meu irmão Henrique Inácio, por todas as palavras de encorajamento e pela sua frase especial “Menina! Não quero essa cabeça para baixo!” que sempre me diz e afirma com a maior convicção.
Aos meus pais, Manuela Inácio e José Inácio, por sempre me incentivarem, por nunca me deixarem desistir, por apoiarem sempre as minhas decisões e escolhas, e por me mostrarem que os objetivos só se atingem com muito trabalho, esforço e dedicação.
O crescente número de imigrantes em Portugal é uma realidade incontornável. De acordo com a Base de Dados Portugal Contemporâneo - PORDATA -, no final de 2014, o número total de imigrantes dos PALOP (Países Africanos de Língua Oficial Portuguesa) em Portugal era cerca de 100.000 e, destes, 75.000 pertencem à população de Lisboa. O fenómeno migratório em Portugal torna-se assim num dos principais fatores para a inserção de variabilidade genética na população portuguesa.
Nos últimos anos, uma nova classe de marcadores autossómicos de inserção/deleção - InDel -, tem vindo a suscitar o interesse da Genética Forense. Estes, são caracterizados pela presença ou ausência de uma sequência específica de nucleótidos. Uma outra classe de marcadores bialélicos - Single Nucleotide Polymorphisms (SNP) - tem, também, nos últimos anos, merecido a atenção da Genética Forense. O estudo de SNP no cromossoma Y; que não sofre recombinação e é transmitido em bloco por via uniparental paterna; pode ter especial utilidade em estudos de ancestralidade e evolução.
Diferenças significativas nas frequências alélicas dos marcadores InDel, tal como noutros marcadores genéticos - Y-SNPs -, entre diferentes grupos ou populações, podem permitir a utilização destes como indicadores de ancestralidade e evolução, ou simplesmente, verificar e confirmar diversidade genética entre grupos e populações.
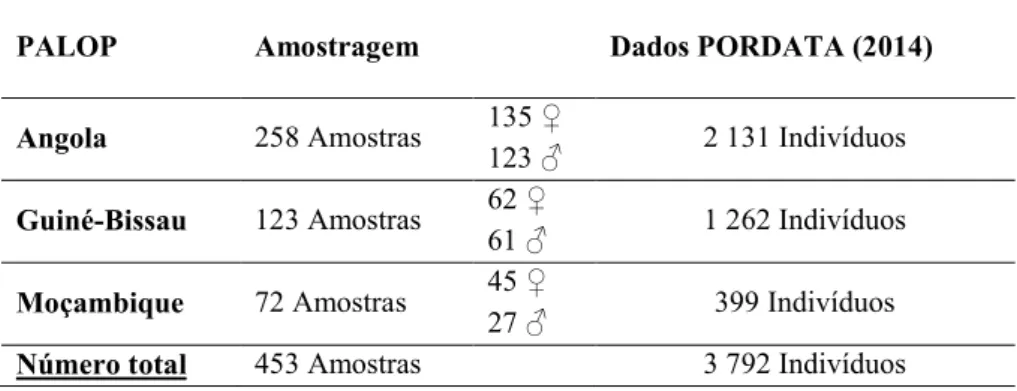
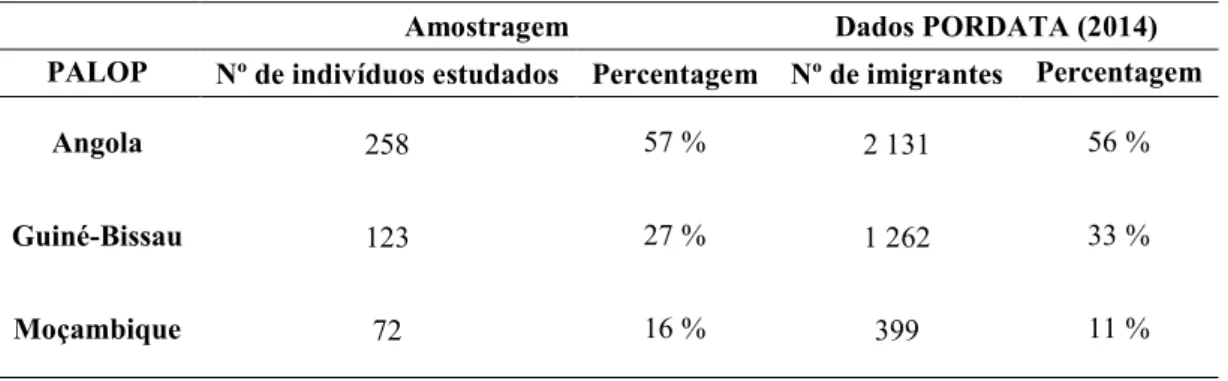
Uma vez que não há dados para os marcadores InDel, bem como para os marcadores do tipo Y-SNPs em imigrantes PALOP a residir na região de Lisboa, o nosso principal objetivo foi a caracterização destes grupos de indivíduos pela tipagem dos mesmos com, pelo menos, 30 InDel e com um painel de 9 Y-SNPs e comparar os diferentes grupos de indivíduos/populações. Assim, para o estudo de marcadores InDel foram estudadas 453 manchas de sangue e para o estudo de marcadores do tipo Y-SNPs foram estudadas 211 manchas de sangue pertencentes a indivíduos do sexo masculino imigrantes de Angola, Moçambique e Guiné-Bissau.
A extração de ADN foi realizada pelo método de Chelex® 100. Após extração as amostras pertencentes ao estudo de marcadores InDel foram tipadas com recurso ao kit Investigator® DIPplex e as amostras destinadas ao estudo de Y-SNPs foram tipadas com recurso ao multiplex 1 descrito por Brion e colaboradores em 2004, constituído por um painel de 9 Y-SNPs.
marcadores InDel estudados se encontravam em equilíbrio de Hardy-Weinberg. Para além disto, também foram calculados parâmetros forenses para todas as populações e foi desenhada uma árvore filogenética representativa das distâncias genéticas entre as populações estudadas. Através dos resultados obtidos para os marcadores do tipo InDel podemos afirmar que estas populações apresentam distâncias genéticas significativas entre elas e entre elas e a população de Lisboa, o que nos permite afirmar que estas populações introduzem variabilidade genética na população de Lisboa. Relativamente, aos marcadores do tipo Y-SNPs é possível verificar algumas diferenças entre as populações africanas, tal como verificado nos marcadores do tipo InDel, mas o painel de Y-SNPs utilizado revelou não ser adequado ao estudo de populações de origem Africana.
Palavras-chave: Populações; Imigrantes; Angola; Moçambique; Guiné-Bissau; InDel; Y-SNPs.
The increasing number of immigrants in Portugal is an unavoidable reality. According to Portugal Contemporary Base - PORDATA - by the end of 2014, the total number of immigrants from PALOP (Portuguese-speaking African countries) in Portugal was about 100.000, and from those, 75.000 are part of Lisboa population. The migratory phenomenon in Portugal can become one of the main factors for the genetic variability.
In the last years, a new class of autosomal insertion/deletion markers - InDel - has gained interest in forensic genetics. InDel are characterized by the presence or absence of a specific sequence of nucleotides.
Significative differences in allele frequencies of InDel and Y-SNPs markers, such as in other genetic markers, between different groups or populations, can be used as ancestry and, eventually, evolutionary indicators or just as indicator of genetic diversity.
Since there is no data for InDel and Y-SNPs markers of PALOP immigrants living in Lisboa, our aim is the characterization of those groups of individuals by typing them with, at least, 30 InDel markers and with a panel of 9 Y-SNPs and compare different groups of individuals/populations. Thus, we studied 453 bloodstain samples for InDel markers and 211 bloodstain samples for Y-SNPs belonging to immigrant individuals from Angola, Guinea-Bissau and Mozambique.
DNA extraction was performed with Chelex® 100 method. After extraction all samples were typed with Investigator® DIPplex method for study of InDel and with multiplex 1 proposed by Brion and collaborators in 2004 for the study of Y-SNPs.
We obtained complete genetic profiles, with the studied genetic markers, for all the 453 studied individuals. Allelic frequencies show that all 30 InDel markers are at Hardy-Weinberg equilibrium. Furthermore, we calculate a panel of forensic parameters for all populations and drawn a phylogenetic tree representative of genetic distances between studied populations.
Through the obtained results with InDel markers we can confirm that those populations show significant genetic distances between them and between them and the host Lisboa population, and so they introduce genetic variability in Lisboa population. In relation to Y-SNPs markers a few differences between the studied African populations are shown, as
Agradecimentos ... i
Resumo ... iii
Abstract ... v
Índice geral ... vii
Índice de tabelas ... xi
Índice de figuras e gráficos ... xiii
Abreviaturas e Siglas ... xvii
Capítulo 1 - Introdução... 1
1.1.Um apontamento sobre o Homem moderno ... 1
1.2.Os fenómenos migratórios a nível mundial ... 3
1.3.A migração em Portugal ... 4
1.4.Populações de imigrantes de Países Africanos de Língua Oficial Portuguesa (PALOP) . 6 1.4.1. República de Angola ... 6
1.4.2. República de Moçambique ... 8
1.4.3. República da Guiné-Bissau ... 9
1.5.Genética Populacional ... 10
1.5.1. Equilíbrio de Hardy-Weinberg - HWE ... 12
1.5.2. Linkage (Dis)equilibrium ... 13
1.5.3. Apontamento sobre métodos estatísticos e estudo de populações ... 14
1.6.Genética Forense ... 15
1.7.Marcadores genéticos em Genética Forense e em Genética Populacional ... 16
1.7.1. Marcadores do tipo STR ... 17
1.7.2. Marcadores do tipo SNP ... 18
1.8.Marcadores do tipo InDel ... 20
1.8.1. Vantagens e desvantagens ... 22
1.8.2. Metodologia de análise: kits comerciais desenvolvidos ... 23
1.9.Marcadores de linhagem ... 26
1.9.1. Marcadores do tipo Y-SNP ... 27
1.9.1.1. Metodologia de análise ...29
1.10. Avaliação de marcadores genéticos... 31
2.2.Amostragem ... 36
2.3.Colheita e armazenamento de amostras ... 37
2.4.Extração de ADN ... 38
2.5.Estudo de marcadores do tipo InDel ... 40
2.5.1. Amplificação por PCR multiplex ... 40
2.5.2. Eletroforese capilar do produto amplificado ... 42
2.5.3. Deteção e análise do produto amplificado ... 43
2.6.Estudo de marcadores Y-SNP ... 43
2.6.1. Amplificação de Y-SNP ... 43
2.6.2. Purificação com Exo-SAP-IT® (USB®) ... 45
2.6.3. Reação de minisequenciação ... 46
2.6.4. Purificação com SAP (USB®) ... 49
2.6.5. Deteção e análise dos produtos SBE ... 50
2.6.5.1. Preparação das amostras para o processo de eletroforese capilar ...50
2.6.5.2. Deteção do produto sequenciado ...51
2.7.Análise estatística e filogenética ... 52
2.7.1. Marcadores do tipo InDel ... 52
2.7.2. Marcadores do tipo Y-SNP ... 52
Capítulo 3 - Resultados e Discussão ... 53
3.1. Estudo de marcadores do tipo InDel ... 53
3.1.1. Microvariantes ... 55
3.1.1.1. Loci HLD92, HLD99 e HLD84 ...55
3.2. Caracterização genética das populações em estudo com marcadores do tipo InDel .... 58
3.2.1. Estudo da população imigrante oriunda de Angola residente na região de Lisboa ... 59
3.2.2. Estudo da população imigrante oriunda de Moçambique residente na região de Lisboa ... 64
3.2.3. Estudo da população imigrante oriunda da Guiné-Bissau residente na região de Lisboa ... 69
3.4. Comparação de populações ... 78
3.5. Análise da variação intrapopulacional... 85
3.6. Análise de distâncias genéticas entre populações ... 87
3.7. Análise de Linkage Disequilibrium ... 89
3.8. Parâmetros forenses ... 90
Capítulo 4 - Conclusão ... 95
Referências Bibliográficas ... 99
Anexos ... 113
Anexo A. Resultados relativos ao estudo dos marcadores Y-SNPs ... 113
Tabela 1: Comparação dos marcadores InDel com os marcadores SNP e STR (adaptado de: Butler, 2012) ... 22 Tabela 2: Painel de 30 marcadores InDel e Amelogenina incluídos no kit Investigator® DIPplex (Qiagen), com respetivo nome dos marcadores, referência na base de dados SNPforID, localização no cromossoma, e sequência inserção/deleção. Adaptado de: Qiagen, 2014 e de http://www.ncbi.nlm.nih.gov/SNP/, consultado a 17.07.2015. ... 25 Tabela 3: Descrição dos multiplex utilizados por Brion et al., em 2004, com respetivo marcador, sequência forward e reverse dos primers, tamanho e concentração. Adaptado de Brion et al., 2004. ... 30 Tabela 4: Descrição das sequências de primers de minisequenciação utilizados na Single
Base Extension (SBE) com respetivo marcador, tamanho e concentração para cada multiplex
utilizado ... 31 Tabela 5: Resumo da amostragem utilizada com respetivos dados PORDATA para cada uma das populações de imigrantes ... 36 Tabela 6: Comparação entre a amostragem utilizada e os dados PORDATA de 2014 e respetivas percentagens das populações de imigrantes em estudo. ... 37 Tabela 7: Componentes e respetivos volumes de preparação da master mix para amplificação com o kit Investigator® DIPplex (Qiagen). ... 40 Tabela 8: Condições do processo de amplificação do kit Investigator® DIPplex (Qiagen) ... 41 Tabela 9: Volumes utilizados por reação para realizar a eletroforese capilar. ... 42 Tabela 10: Componentes e respetivos volumes de preparação da master mix para amplificação com o kit Qiagen® Multiplex PCR. ... 44 Tabela 11: Condições de amplificação dos marcadores do tipo Y-SNP ... 44 Tabela 12: Ciclos realizados no termociclador para purificação com ExoSAP-IT® (USB®) 46 Tabela 13: ddNTP, respetivos fluorocromos e indicação da cor atribuída na sua análise ... 48 Tabela 14: Condições do processo de minisequenciação dos marcadores do tipo Y-SNP com recurso ao SNaPshot® Multiplex kit (Applied Biosystems). ... 48 Tabela 15: Ciclos realizados no termociclador para purificação com SAP (USB®). ... 49 Tabela 16: Volumes utilizados por reação para realizar a eletroforese capilar ... 50 Tabela 17: Dados relativos aos 30 loci estudados para a população imigrante oriunda de Angola residente na região de Lisboa: número de alelos estudados (2N), frequências alélicas,
Tabela 18: Dados relativos aos 30 loci estudados para a população imigrante oriunda de Moçambique residente na região de Lisboa: número de alelos estudados (2N), frequências alélicas, frequências de heterozigotia (H) e homozigotia (h), heterozigotia observada (Ho) e esperada (He) e o valor de p para o HWE. ... 65 Tabela 19: Dados relativos aos 30 loci estudados para a população de imigrantes oriunda da Guiné-Bissau residente na região de Lisboa: número de alelos estudados (2N), frequências alélicas, frequências de heterozigotia (H) e homozigotia (h), heterozigotia observada (Ho) e esperada (He) e o valor de p para o HWE. ... 70 Tabela 20: Caracterização dos Y-SNP que constituem o multiplex utilizado no presente estudo e descrito por Brion e colaboradores, em 2004. ... 77 Tabela 21: Caracterização dos haplogrupos presentes nas três populações de imigrantes estudadas - Angola, Guiné-Bissau e Moçambique -, em que é possível observar o número de indivíduos em que estão presentes (N) e respetiva frequência. ... 77 Tabela 22: Comparação dos resultados obtidos para as frequências alélicas para os 30 loci InDel estudados, obtidos para as populações de imigrantes oriundas de Angola, de Moçambique e da Guiné-Bissau, residentes na região de Lisboa ... 80 Tabela 23: Comparação das heterozigotias observadas para os 30 loci InDel estudados, obtidas para as populações de imigrantes oriundas de Angola, Moçambique e Guiné-Bissau, residentes na região de Lisboa ... 83 Tabela 24: Tabela AMOVA obtida através da análise dos 30 loci InDel para cada uma das populações de imigrantes em estudo: Angola, Moçambique e Guiné-Bissau ... 85 Tabela 25: Valores de diferenciação genética (Fst) obtidos para as populações de imigrantes
de Angola, de imigrantes de Moçambique e de imigrantes da Guiné-Bissau, com respetivos
p-value. ... 86
Tabela 26: Matriz de distâncias genéticas (Fst) entre as três populações em estudo - imigrantes de Angola, Moçambique e Guiné-Bissau - e entre duas populações analisadas com os mesmos marcadores genéticos - Portugal e imigrantes de Cabo Verde -. ... 87 Tabela 27: Parâmetros forenses calculados para os 30 marcadores InDel estudados nas populações de imigrantes de Angola, Moçambique e Guiné-Bissau residentes na região de Lisboa ... 90 Tabela 28: Parâmetros forenses combinados para os 30 marcadores InDel em estudo. ... 92
Figura 1: Representação esquemática da teoria “Out-of-Africa” (Fonte: http://www.progon os.com/furuti/MapProj/Dither/ProjAppl/projAppl.html, consultado a 28.04.2015) ... 2 Figura 2: Representação esquemática da Teoria da Evolução Multiregional. Em que EU, representa a Europa, AF, África e AA, Ásia. Fonte: (Hammer e Zegura, 1996) ... 3 Figura 3: Principais nacionalidades de populações emigrantes residentes em Portugal. Superiormente é possível verificar o número de cidadãos residentes em Portugal bem como a respetiva percentagem e inferiormente está descrito se houve um decréscimo (▼) da população ou acréscimo (▲) no ano de 2014 Fonte: (Serviço de Estrangeiros e Fronteiras - SEF -, 2014) ... 5 Figura 4: Países Africanos de Língua Oficial Portuguesa. (Fonte: http://www.legis-palop.org/bd/Base de dados oficial dos PALOP, consultado a 08.06.2015) ... 6 Figura 5: República de Angola. (Fonte: http://www.africa-turismo.com/mapas/angola.htm consultado a 09.06.2015) ... 7
Figura 6: República de Moçambique. (Fonte: http://www.africa-turismo.
com/mapas/mocambique.htm consultado a 10.06.2015) ... 8 Figura 7: República da Guiné-Bissau. (Fonte: http://www.africa-turismo.com/mapas/guine-bissau.htm consultado a 11.06.2015) ... 9 Figura 8: Representação esquemática da constituição do genoma humano (adaptado de: Alberts, 2008) ... 16 Figura 9: Exemplo de um SNP em que é possível observar que a sequência de ADN 1 é
diferente da sequência de ADN 2, num único par de base, C-G e T-A (Fonte: Pinheiro, 2010) ... 19
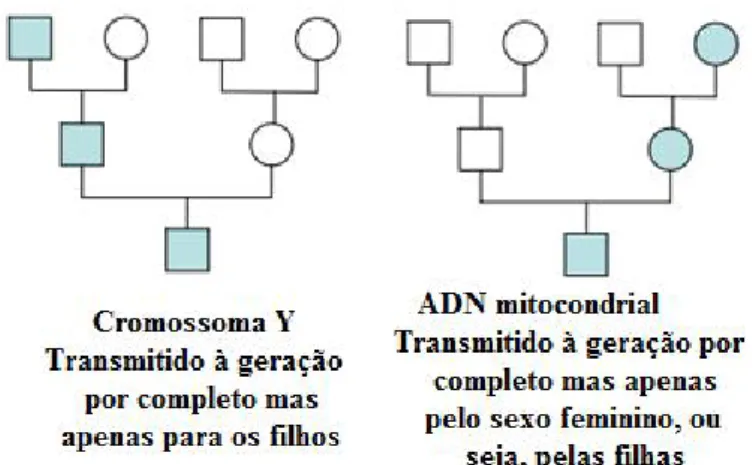
Figura 10: Exemplo de um InDel em que é possível observar a sequência original e as sequências mutadas, deleção de uma Timina e inserção de uma Guanina (Adaptado de: Barış, Karadayı e Yanmış, 2013) ... 21 Figura 11: Exemplos de transmissão uniparental, à esquerda, por via paterna, pelo cromossoma Y e, à direita, por via materna, ADN mitocondrial (□ – indivíduos do sexo masculino; ○ – indivíduos do sexo feminino) (Adaptado de: Butler, 2012) ... 27 Figura 12: Fluxograma representativo das diversas etapas laboratoriais para o estudo de marcadores do tipo InDel e de marcadores do tipo Y-SNP ... 35
Figura 14: Representação esquemática do processo de ação da ExoSAP-IT® (USB®). (adaptado de: http://www.affymetrix.com/catalog/131310/USB/ExoSAPIT+For+PCR+
Product+Cleanup#1_2; consultado a 03.08.2015) ... 45 Figura 15: Esquema geral do processo de minisequenciação por aplicação do SNaPshot® Multiplex (Applied Biosystems) (Adaptado de: Corporation 2012) ... 47 Figura 16: Processo de minisequenciação através do SNaPshot® Multiplex (adaptado de: Applied Biosystems 2005) ... 47 Figura 17: Exemplo de um eletroferograma obtido aquando a análise dos marcadores do tipo InDel. Apresenta-se um genótipo de um indivíduo do sexo masculino como é possível observar pela análise do marcador da amelogenina. Para além disto, é possível observar os 30 marcadores presentes no kit Investigator® DIPplex (Qiagen), em que cada painel de cores corresponde à fluorescência emitida pelos fluorocromos, com exceção do preto que corresponde à fluorescência emitida no comprimento de onda amarelo. Cada painel contém entre 7 a 8 marcadores ... 53 Figura 18: Microvariantes detetadas aquando a análise dos marcadores InDel. Em 1. é possível observar o pico out off ladder D92+ (+1), detetado no painel correspondente à cor verde, com um nucleótido a mais do que aquele que era esperado como se verifica em 2. para o ladder alélico ... 56 Figura 19: Microvariantes detetadas aquando a análise dos marcadores InDel. Em 1. é possível observar o pico out off ladder D99+ (-1) e em 2. o pico out off ladder D99- (+1), ambos detetados no painel correspondente à cor verde, com um nucleótido a menos e a mais, respetivamente, do que aquele que era esperado como se verifica em 3. para o ladder alélico ... 56 Figura 20: Microvariantes detetadas aquando a análise dos marcadores InDel. Em 1. é possível observar o pico out off ladder D84+ (-4), detetado no painel correspondente à cor vermelha, com um nucleótido a mais do que aquele que era esperado como se verifica em 2. para o ladder alélico ... 57
Y-SNP utilizados, em que para cada loci é possível observar a base nucleotídica correspondente. Cada uma das bases possui uma coloração diferente consoante a fluorescência emitida pelos fluorocromos presentes, em que a adenina (A) corresponde ao verde, a citosina (C) ao preto (corresponde à fluorescência emitida no comprimento de onda amarelo), a timina (T) ao vermelho e a guanina (G) ao azul ... 75 Figura 22: Árvore filogenética com os polimorfismos do cromossoma Y bialélicos estudados por Brion e colaboradores, em 2004. A vermelho encontra-se o multiplex utilizado no presente estudo e cada marcador presente no mesmo ... 75 Figura 23: Árvore representativa das distâncias genéticas calculadas pelo índice de fixação apresentada na tabela 24 ... 88
Gráfico 1: Frequências alélicas para os 30 marcadores InDel analisados para a população de imigrantes oriundos de Angola residente na região de Lisboa ... 62 Gráfico 2: Heterozigotias observada e esperada para os 30 marcadores InDel analisados para a população de imigrantes oriundos de Angola residente na região de Lisboa ... 63 Gráfico 3: Frequências alélicas para os 30 marcadores InDel analisados para a população de imigrantes oriunda de Moçambique residente na região de Lisboa ... 67 Gráfico 4: Heterozigotias observada e esperada para os 30 marcadores InDel analisados para a população de imigrantes oriunda de Moçambique residente na região de Lisboa ... 68 Gráfico 5: Frequências alélicas para os 30 marcadores InDel analisados para a população de imigrantes oriunda de Guiné-Bissau residente na região de Lisboa ... 72 Gráfico 6: Heterozigotias observada e esperada para os 30 marcadores InDel analisados para a população de imigrantes oriunda da Guiné-Bissau residente na região de Lisboa ... 73 Gráfico 7: Comparação dos resultados obtidos para as frequências alélicas para os 30 loci InDel estudados, obtidos para as populações de imigrantes oriundos de Angola (ANG), de Moçambique (MOÇ) e da Guiné-Bissau (GB), residentes na região de Lisboa ... 79 Gráfico 8: Comparação das heterozigotias observadas para os 30 loci InDel estudados, obtidas para as populações imigrantes oriundas de Angola, Moçambique e Guiné-Bissau, residentes na região de Lisboa ... 82 Gráfico 9: Distribuição das frequências dos haplogrupos presentes nas três populações imigrantes residentes em Lisboa em estudo, Angola, Guiné-Bissau e Moçambique ... 84
A A: Adenina
ABC: Modelo bayesiano aproximado de computação
ADN: Ácido Desoxirribonucleico ADNmt: Ácido Desoxirribonucleico mitocondrial
AMOVA: Análise de variância molecular ANG: Angola
ARN: Ácido ribonucleico B BSA: Bovine Serum Albumin
C C: Citosina
D
ddNTP: Didesoxinucleótidos trifosfatados
DIP: Deletion-Insertion Polymorphisms dNTP: Desoxinucleótidos trifosfatados F Fst: Índice de Fixação G G: Guanina GB: Guiné-Bissau H H: Taxa de heterozigotia h: Taxa de homozigotia He: Heterozigotia Esperada Ho: Heterozigotia Observada
HWE: Equilíbrio de Hardy-Weinberg
I InDel: Inserção/Deleção
INMLCF: Instituto Nacional de Medicina Legal e Ciências Forenses
L LD: Linkage Disequilibrium LE: Linkage Equilibrium
LINE: Long Interspersed Elements LTR: Long Terminal Repeats
M MOÇ: Moçambique
N NJ: Neighbor Joining
NRY: Porção Não Recombinante do Cromossoma Y
P
PALOP: Países Africanos de Língua Oficial Portuguesa
PCR: Polymerase Chain Reaction PD: Poder de Discriminação
PDC: Poder de Discriminação Combinado PE: Poder de Exclusão
PEC: Poder de Exclusão Combinado PIC: Polymorphism Information Content PM: Probabilidade de Matching
PMA: Países Menos Avançados
PMC: Probabilidade de Matching
Combinada
PORDATA: Base de Dados Portugal Contemporâneo
S
SAP: Shrimp Alkaline Phosphatase SBE: Single Base Extension
SEF: Serviço de Estrangeiros e Fronteiras SGBF: Serviço de Genética e Biologia Forenses
SINE: Short Interspersed Elements SNP: Single Nucleotide Polymorphism SNP: Single Nucleotide Polymorphism STR: Short Tandem Repeats
T: Timina
TPI: Typical Paternity Índex - Índice de Paternidade Típico
TPIC: Índice de paternidade típico combinado
V
VNTR: Variable Number Tandem Repeats Y
1.1. UM APONTAMENTO SOBRE
A origem do Homem moderno
das ciências. Desde a História, à Arqueologia e à entre outras, sempre se procurou
estudo levado a cabo por Klein, o “seres humanos fisicamente modernos quando o único habitante da Eurásia era o H seguida uma expansão sucessiva do H
e Europa Ocidental. O Homem não evoluiu diretamente dos
sapiens sapiens, tendo atravessad
de humanização ao longo de milhões de anos. O que atualmente designamos por
características, tais como, habilidades cognitivas e comunicativas, que se c uma forma muito sucinta,
desenvolvimento do comportamento humano depende de uma mudança a nível neurológico, ou seja, de uma alteração na organização do cérebro, e não
tamanho (Klein, 1995). No entanto, esta teoria comprovar os seus princípios
acerca da estrutura do cérebro
Existem duas grandes teorias que tentam explicar teoria “Out-of-Africa” e a teori
maior acolhimento na comunidade científica.
UM APONTAMENTO SOBRE O HOMEM MODERNO
omem moderno sempre despertou um especial interesse
Desde a História, à Arqueologia e à Antropologia, até à Genética Molecular entre outras, sempre se procurou respostas para a origem do Homem moderno.
estudo levado a cabo por Klein, os registos fósseis existentes demonstram que os primeiros seres humanos fisicamente modernos” surgiram em África há cerca de 50 000 anos atrás,
ico habitante da Eurásia era o Homem de Neandertal (Klein, 1995)
uida uma expansão sucessiva do Homem moderno para a Ásia Ocidental, Europa Oriental omem não evoluiu diretamente dos chimpanzés, mas sim do atravessado os períodos Paleolítico e Neolítico, e um longo processo de humanização ao longo de milhões de anos.
que atualmente designamos por Homem moderno está associado a características, tais como, habilidades cognitivas e comunicativas, que se c uma forma muito sucinta, pela capacidade para a “cultura”. A
desenvolvimento do comportamento humano depende de uma mudança a nível neurológico, ou seja, de uma alteração na organização do cérebro, e não de uma modificação d
No entanto, esta teoria depara-se com um obstáculo ao tentar os seus princípios, uma vez que, os crânios fósseis oferecem pouca informação
(Portin, 2007).
Existem duas grandes teorias que tentam explicar a evolução do H Africa” e a teoria da Evolução Multiregional. No entanto,
na comunidade científica.
Capítulo 1
INTRODUÇÃO
nteresse a várias áreas Genética Molecular, omem moderno. Segundo o entes demonstram que os primeiros surgiram em África há cerca de 50 000 anos atrás, (Klein, 1995), havendo de omem moderno para a Ásia Ocidental, Europa Oriental , mas sim do Homo Neolítico, e um longo processo
está associado a determinadas características, tais como, habilidades cognitivas e comunicativas, que se caracterizam, de Acredita-se que o desenvolvimento do comportamento humano depende de uma mudança a nível neurológico, de uma modificação do seu e com um obstáculo ao tentar cem pouca informação
a evolução do Homem moderno. A a da Evolução Multiregional. No entanto, a primeira merece
Capítulo 1
INTRODUÇÃO
A hipótese “Out-of-Africa” afirma que o Homem moderno tem uma origem relativamente recente em África e que, de seguida, se moveu de África para outras partes do Mundo, principalmente para a Euro-Ásia, substituindo as populações de Homo Erectus existentes (Figura 1). Os defensores desta teoria afirmam que não há nenhuma evidência fóssil relevante que demonstre a continuidade entre o Homem não moderno e o Homem moderno em regiões afastadas do Mundo. Estes, acreditam na existência do fenómeno de deriva genética e de seleção natural que leva as populações arcaicas a percorrerem diferentes trajetórias evolutivas para diferentes continentes (Klein, 1995; Mountain, 1998; Nitecki e Nitecki, 1994; Pinheiro, 2010; Salas et al., 2004).
Figura 1: Representação esquemática da teoria “Out-of-Africa”.
(Fonte: http://www.progonos.com/furuti/MapProj/Dither/ProjAppl/projAppl.html, consultado a 28.04.2015) Quanto à Teoria da Evolução Multiregional (figura 2), esta afirma que o Homem moderno evoluiu diretamente de seres humanos arcaicos e surgiu simultaneamente em todas as regiões do Mundo, não só em África mas também na Europa e na Ásia, onde não existiam homens modernos, negando assim a origem africana, dando realce ao fluxo de genes ao longo do tempo entre as populações combinado com a seleção natural e argumentando uma origem múltipla entre as regiões geográficas (Hammer e Zegura, 1996; Klein, 1995; Portin, 2007; Salas et al., 2007).
Figura 2: Representação esquemática da Teoria da Evolução Multiregional. Em que EU, representa a Europa, AF, África e AA, Ásia.
Fonte: (Hammer e Zegura, 1996)
Os estudos de variação molecular podem permitir reconstruir eventos passados tal como expansão de populações e colonização de regiões, permitindo uma tentativa de correlação entre variação genética com aspetos da história humana, tal como, origem de uma população e movimentos migratórios (Underhill et al., 1996).
1.2.OS FENÓMENOS MIGRATÓRIOS A NÍVEL MUNDIAL
O Homem, desde a sua existência, sempre se deslocou de uma região geográfica para outra em busca, principalmente, de alimentação (Teixeira, Braga e Baeninger, 2012).
O fenómeno migratório pode caracterizar-se pelo deslocamento de indivíduos de determinado espaço geográfico para outro, de forma temporária ou permanente (Martin e Zürcher, 2008). Existem estudos realizados que comprovam que a população jovem tende a emigrar de uma forma mais temporária do que propriamente permanente, pois tem esperança de regressar ao seu país de origem a dada altura (Lopes, 2013).
A partir do início do século 20 houve um aumento exponencial do fenómeno migratório mundial devido às desigualdades a nível demográfico e económico, tal como, desigualdades no mercado de trabalho, mas também devido às oportunidades que surgiram e ao desenvolvimento na mobilização da população de determinada região para outras regiões geográficas mais distantes.
Existem diversos fatores que desencadeiam as migrações, entre eles, económicos, culturais, religiosos, políticos ou mesmo devido a causas naturais, tais como, terramotos, sendo que o fator com maior relevância é o económico (Cardoso, 2007; Castles, 2011). Na
maioria dos casos, os indivíduos deslocam-se não só por uma questão de sobrevivência a nível “materialístico” mas também na esperança e na expetativa de um futuro melhor tanto para si como para os seus descendentes.
O fenómeno migratório mundial tem vindo a aumentar com o decorrer dos anos. Segundo as últimas estimativas das Nações Unidas, cerca de 3,2% da população a nível mundial é população que saiu do seu país de origem e emigrou com objetivo de encontrar uma vida melhor (Martin e Zürcher, 2008). Normalmente, a população desloca-se de países em vias de desenvolvimento para países desenvolvidos mas recentemente até pessoas de países desenvolvidos tendem a deslocar-se para países mais desenvolvidos que os de origem, onde existem maiores oportunidades a nível de empregabilidade.
1.3.A MIGRAÇÃO EM PORTUGAL
Com a queda do Império Colonial Português, Portugal sofreu uma mudança rápida devido à perda das colónias africanas, à recentralização e, mais tarde, à adesão à União Europeia. Assim, Portugal, historicamente, passou de um país emigrante, devido ao grande fenómeno de emigração dos anos 60 e 70, para um país imigrante, muito especialmente resultado de imigrantes provenientes de África, do Brasil e da Europa de Leste. No entanto, contrariamente a outros países da Europa, Portugal continua a experienciar ambos os fenómenos de emigração e imigração.
A imigração em Portugal é um fenómeno social de extrema relevância, uma vez que, o enorme fluxo de emigração da população portuguesa causou escassez a nível de mão-de-obra em alguns dos setores industriais. Para compensar esta escassez surgiu um recrutamento de força de trabalho a partir das colónias africanas, uma vez que nesta época as colónias africanas eram oficialmente território português. Este fenómeno é denominado por migração inter-regional de cidadãos portugueses (Baganha, 1998; Pinheiro, 2008).
Existem indivíduos de diversos países que emigram para Portugal mas os principais países fonte de emigrantes para Portugal em busca de trabalho, para além do Brasil e dos países de Leste, são os Países Africanos de Língua Oficial Portuguesa (PALOP), entre eles, Cabo Verde, Angola, Guiné-Bissau, Moçambique e São Tomé e Príncipe (Rocha-Trindade, 2005). Este fenómeno de imigração deve-se muito à liberalização do mercado de trabalho para “mão-de-obra” não qualificada, à falta de oportunidades de emprego nos seus países de
origem e à instabilidade política existente nos mesmos, o que levou a desencadear guerras civis.
Para fazer uma análise da evolução da população estrangeira em Portugal devem considerar-se diversos fatores, entre eles, a evolução legislativa ocorrida nos últimos anos, o facto de Portugal ser um país de imigrantes e emigrantes, as relações históricas e culturais com os outros países e os contextos económicos e sociais tanto de Portugal como dos países de origem (Serviço de Estrangeiros e Fronteiras - SEF -, 2013).
Segundo o Serviço de Estrangeiros e Fronteiras, no ano de 2014 ocorreu um decréscimo do número de estrangeiros em Portugal, sendo que apenas 395.195 cidadãos possuíam título de residência válido. A explicação para este decréscimo assenta na base legal da aquisição de nacionalidade portuguesa, na alteração de fluxos migratórios e no impacto da crise económica presente no mercado de trabalho.
Através da figura 3 é possível verificar as dez principais nacionalidades de populações emigrantes residentes em Portugal bem como se houve um decréscimo ou acréscimo das mesmas. É de salientar que a nacionalidade brasileira é a principal comunidade estrangeira residente, seguida de Cabo Verde, incluída nos PALOP.
Figura 3: Principais nacionalidades de populações emigrantes residentes em Portugal. Na parte superior é possível verificar o número de cidadãos residentes em Portugal bem como a respetiva percentagem e na parte
inferior está descrito se houve um decréscimo (▼) da população ou acréscimo (▲) no ano de 2014 Fonte: (Serviço de Estrangeiros e Fronteiras - SEF -, 2014).
Seguidamente dedicamos uma secção à caracterização sumária dos principais países de origem dos imigrantes dos PALOP.
1.4.POPULAÇÕES DE IMIGRANTES DE PAÍSES AFRICANOS DE LÍNGUA OFICIAL PORTUGUESA (PALOP)
Os Países Africanos de Língua Oficial Portuguesa, PALOP, dizem respeito aos países africanos que possuem a língua portuguesa como a sua língua oficial, entre eles, Angola, Cabo Verde, Guiné-Bissau, Moçambique e São Tomé e Príncipe (figura 4). São todos ex-colónias de Portugal e entre os anos de 1974 e 1975, tornaram-se independentes devido à descolonização de Portugal. Todos seguiram o seu caminho de independência de acordo com as suas potencialidades económicas e políticas. Contudo, a exclusão social, a pobreza, a fome e a violência ainda são características que demarcam muito os PALOP, apesar destes terem vindo a firmar vários protocolos com outros países e organizações nos mais diversos campos, entre eles, na cultura, educação, economia e diplomacia.
Figura 4: Países Africanos de Língua Oficial Portuguesa. (Fonte: http://www.legis-palop.org/bd/Base de dados oficial dos PALOP, consultado a 08.06.2015)
1.4.1. REPÚBLICA DE ANGOLA
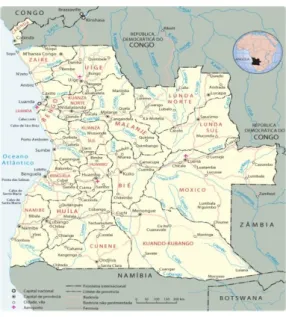
Angola, oficialmente designada por República de Angola, localiza-se na costa
ocidental de África sendo considerada como o maior dos PALOP com 1 246 700 Km2, com
extensão da costa atlântica de cerca de 1 650 Km e delimitada pela República do Congo, pela República Democrática do Congo, pela República da Zâmbia, pela República da Namíbia e pelo Oceano Atlântico como é possível verificar na figura 5. É constituída por cerca de 12 milhões de habitantes, 49,3% do sexo masculino e 50,7% do sexo feminino (Embaixada da
República de Angola em Portugal, 2015, The Embassy of the Republic of Angola and GlobeScope Inc., 2015).
Figura 5: República de Angola.
(Fonte: http://www.africa-turismo.com/mapas/angola.htm consultado a 09.06.2015)
A República de Angola é constituída por 18 províncias, são elas, Bengo, Benguela, Bié, Cabinda, Kuano-Kubango, Kwanza-Norte, Kwanza-Sul, Cunene, Huambo, Huíla, Luanda, Luanda-Norte, Luanda-Sul, Malanje, Moxico, Namibe, Uíge e Zaire, em que Luanda é a capital com cerca de 3 000 000 habitantes (The African Union Commission, 2015).
O país apresenta duas grandes regiões climáticas distintas, a região litoral e a região do interior, devido a possuir uma posição geográfica peculiar, por se situar na zona inter-tropical e sub-tropical do hemisfério Sul, próximo do mar, e pelas especificidades do seu relevo. Assim, o clima é maioritariamente tropical, apresentando duas estações, a estação seca denominada por Cacimbo, de maio a setembro e a estação das Chuvas, período mais quente, de setembro a maio, apresentando temperaturas médias entre os 17 ºC e os 27 ºC. Possui um património natural rico na sua flora e na sua fauna, possuindo florestas húmidas, savanas e zonas desérticas.
1.4.2. REPÚBLICA DE MOÇAMBIQUE
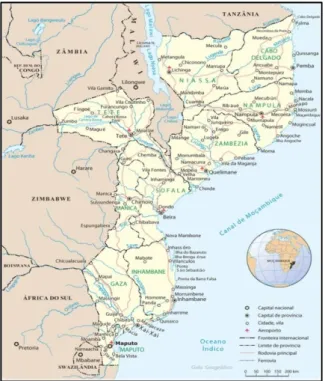
A República de Moçambique localiza-se no sudeste de África, limitado a Norte pela Zâmbia, Malawi e Tanzânia, a Leste pelo Canal de Moçambique e pelo Oceano Índico, a Sul e Oeste pela África do Sul e a Oeste pela Suazilândia e pelo Zimbabwe (figura 6). Este país é
o segundo maior dos PALOP, com cerca de 801 590 Km2 e mais de 20 milhões de habitantes
(Embaixada da República de Moçambique em Portugal, 2015).
Figura 6: República de Moçambique. (Fonte: http://www.africa-turismo.com/mapas/mocambique.htm consultado a 10.06.2015)
A República de Moçambique é constituída por 10 províncias, entre elas, Cabo Delgado, Gaza, Inhambane, Manica, Maputo, Nampula, Niassa, Sofala, Tete e Zambézia, em que Maputo é a maior cidade do país e a sua capital, e durante o período de domínio português denominava-se por Lourenço Marques (Portal do Governo de Moçambique, 2015). Apesar de ser um dos maiores PALOP ainda é um dos mais instáveis a nível político e a nível de segurança, embora tenha um processo de democratização mais avançado que o de Angola (Salgado, 2008).
A República de Moçambique possui uma vasta fauna e flora, uma vez que, o seu terreno varia desde florestas tropicais a montanhas, campos, dunas e praias e compreende várias áreas classificadas como parques nacionais e reservas naturais. Em termos climáticos
apresenta duas estações principais, a estação das chuvas de novembro até março e a estação seca de abril a outubro, sendo um país com um clima geralmente tropical e húmido (Countries
and Their Cultures, 2015; Mcsweeney, New e Lizcano, 2010).
Como país africano que é, a República de Moçambique é rica em recursos naturais, como gás natural, minerais, madeiras, entre outros e as principais exportações dizem respeito, a algodão, açúcar, camarões e chá.
1.4.3. REPÚBLICA DA GUINÉ-BISSAU
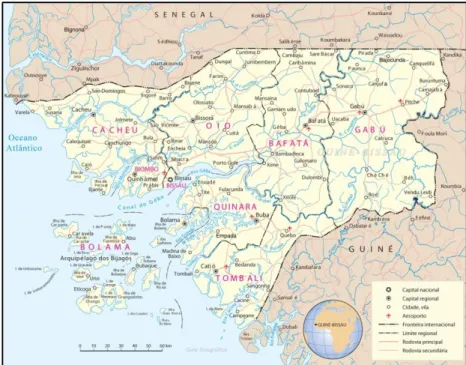
A República da Guiné-Bissau é um dos PALOP, em termos de dimensão, mais
pequeno, com cerca de 36 120 Km2 e com cerca de 1 500 000 habitantes, 52% dos quais do
sexo feminino. A capital do país é Bissau. Esta República situa-se na costa ocidental de África e faz fronteira com o Senegal a Norte, com a Guiné Conacri a Este e Sudoeste e com o Oceano Atlântico a Sul e a Oeste. O seu território pode ser dividido em duas grandes componentes distintas, uma continental, com um cordão de ilhas adjacentes, e uma insular, o arquipélago do Bijagós, constituído por cerca de 40 ilhas, separado do continente pelos Canais de Gêba, Pedro Álvares, Bolama e Canhabaque (figura 7).
Figura 7: República da Guiné-Bissau. (Fonte: http://www.africa-turismo.com/mapas/guine-bissau.htm consultado a 11.06.2015)
A Guiné-Bissau apresenta uma paisagem pouco uniforme ao longo do seu território, sendo que na parte continental o seu relevo se caracteriza pela existência de uma região costeira de planícies baixas, pantanosas e mal drenadas. Quanto às zonas mais altas estas encontram-se na Região do Boé, onde ficam os planaltos Bafatá e Gabu, com altura não superior a 300 metros e o clima é maioritariamente tropical com temperaturas médias de 20ºC.
A República da Guiné-Bissau, a nível económico, é um país pobre, fazendo parte do Grupo de Países Menos Avançados, PMA, em que a maioria da população vive com menos de dois dólares americanos por dia, possuindo uma dependência alimentar enorme e também um atraso a nível tecnológico. Possui alguns recursos minerais, ainda por explorar, como acontece, também, com a pesca, uma das suas riquezas, além das potencialidades turísticas. Devido à escassez a nível monetário, os fluxos migratórios deste país são elevados tanto para fora do país como em termos de êxodo rural dentro do país.
1.5.GENÉTICA POPULACIONAL
Desde a antiguidade que o Homem tenta perceber a transmissão das características de um indivíduo para o outro. As primeiras ideias sobre hereditariedade eram bastante simples e afirmavam que os filhos eram semelhantes aos pais, sem entender ou explicar o mecanismo por detrás desta afirmação.
A Genética é a área da Ciência, mais concretamente da Biologia, que de entre outras matérias, estuda a hereditariedade, ou seja, a forma como as características são transmitidas de geração em geração, sendo que teve os primeiros avanços como Ciência com as leis propostas por Mendel, em 1866.
Uma população é definida como um conjunto de indivíduos da mesma espécie que habitam na mesma área geográfica, que compartilham um ancestral comum, apresentando uma continuidade no tempo e cujos indivíduos possuem a capacidade de se relacionarem ao acaso e, portanto, trocarem características genéticas entre si. Toda a população possui um
pool genético que a caracteriza, e que é transmitido de geração em geração (Hartl e Jones,
1998; Hodge, 2010; Kartavtsev, 2015).
O estudo da evolução assenta em dois princípios básicos, no mecanismo de hereditariedade e no mecanismo de variação, sendo que a evolução está relacionada com as alterações ocorridas em determinada população. A variação genética normalmente presente
nas populações permite que, ao longo das gerações, sejam criadas infinitas combinações genotípicas. Outros fatores como a seleção natural, deriva genética, mutação, fluxo de genes, seleção sexual, entre outros, podem modificar as frequências de genes e de genótipos, designando-se este fenómeno por evolução biológica (Mulligan et al., 2004).
O desenvolvimento da Genética Populacional ocorreu com a definição do princípio do Equilíbrio de Hardy-Weinberg que afirma que numa população infinita, que se reproduz ao acaso e que não é afetada por mutação, migração ou seleção natural, em que todos os seus organismos são férteis e viáveis, tanto as frequências alélicas como as genotípicas se mantêm constantes ao longo das gerações.
A Genética Populacional, como área da biologia evolutiva, estuda o processo evolutivo das populações, como disciplina fundamental para a compreensão do funcionamento da evolução biológica, procurando determinar de que forma os mecanismos evolutivos, tais como, a mutação e recombinação atuam sobre as populações (Madrigal, 2006;
Peng, Kimmel e Amos, 2012). Assim, esta é presentemente um dos campos mais produtivos
da biologia evolutiva, uma vez que estuda as frequências alélicas, genotípicas e fenotípicas de determinada população e, para além disso, estuda a distribuição dos alelos de um determinado gene na população e os fatores que mantêm ou alteram a frequência desses mesmos alelos e genótipos de geração em geração (Hartl e Jones, 1998; Papiha, Deka e Chakraborty, 1999; Templeton, 2006). Entende-se por alelos as versões alternativas de determinado gene ou marcador genético que se encontram no mesmo locus em cromossomas homólogos. Quanto às frequências alélicas, estas dizem respeito às proporções dos diferentes alelos de um determinado locus na população. Já as frequências genotípicas dizem respeito às proporções dos diferentes genótipos para o locus considerado. Entende-se por locus o local físico no cromossoma onde se encontra situado determinado gene, ou sequência de ADN (Kartavtsev, 2015).
A genética populacional faz uma análise das frequências alélicas e genotípicas observadas numa população ou em diferentes grupos da população. O cálculo da frequência de um determinado perfil de ADN numa dada população é possível com recurso a uma base de dados populacional, a qual compreende a frequência dos alelos dos marcadores genéticos nessa população (Gill et al., 2006; Martin, Schmitter e Schneider, 2001; Wallace et al., 2014).
1.5.1. EQUILÍBRIO DE HARDY-WEINBERG - HWE
O conhecimento das frequências alélicas é útil para realizar inferências acerca de determinados fenómenos populacionais, entre eles, fenómenos migratórios, seleção natural, entre outros. As frequências alélicas apresentam uma maior utilidade que as frequências genotípicas devido ao facto dos alelos raramente sofrerem mutação numa determinada geração, por isso consideram-se estáveis na transmissão de geração em geração, contrariamente aos genótipos que podem ir mudando entre gerações. Daí a frequência genotípica não ser tão útil, uma vez que, é alterada pelos processos de segregação e de recombinação que ocorrem durante a meiose.
O equilíbrio de Hardy-Weinberg (HWE, do inglês, Hardy-Weinberg equilibrium) foi descrito no início do século XX e representa um marco importante da genética populacional (Emigh, 2013; Rodriguez, Gaunt e Day, 2009; Wigginton, Cutler e Abecasis, 2005).
Segundo o princípio do equilíbrio de Hardy-Weinberg as frequências alélicas mantêm-se constantes de geração em geração, a menos que alguma força atue de modo a alterá-las. Quando não há forças perturbantes, tal como a seleção, mutação ou migração que afetariam as frequências alélicas ao longo do tempo e tratando-se de uma população em que se dá o acasalamento ao acaso, as frequências genotípicas são dadas pelo produto das frequências alélicas na população e diz-se que a população se encontra em HWE (Bourgain et
al., 2004; Tamarin, 2001; Waples, 2015; Ward e Carroll, 2014).
Considerando as inferências do HWE pode afirmar-se que existem quatro forças de evolução, entre elas, a mutação, a seleção natural, a deriva genética e o fluxo de genes. Relativamente à mutação, esta diz respeito a uma mudança aleatória no genoma, resultando na introdução de novos alelos em determinada população com uma frequência baixa, ou seja, qualquer processo que altere determinado alelo (Nei, 1975; Tamarin, 2001; Waples, 2015). Quanto às outras três forças evolutivas, estas atuam nas mutações e causam o aumento ou o decréscimo da frequência. A seleção natural ocorre quando os indivíduos com diferentes genótipos têm probabilidades diferentes de sobrevivência ou reprodução. É um processo que ocorre nas populações se determinadas condições se verificarem e que possui consequências. Os indivíduos menos susceptíveis poderão transmitir os seus alelos à geração seguinte com maior facilidade, alterando as frequências alélicas com o decorrer do tempo (Slatkin, 1987; Tamarin, 2001; Templeton, 2006). O conceito de deriva genética está intimamente ligado à flutuação aleatória das frequências alélicas de uma população ao longo do tempo, flutuação
esta resultante do tamanho da população, de tal forma que em populações mais pequenas é expetável maior deriva genética que em populações maiores ou em populações teoricamente infinitas (Masel, 2012; Templeton, 2006). Por último, o fluxo de genes é o movimento de alelos de uma população para outra através da migração de indivíduos e, assim, pode introduzir novos alelos numa população, e age para reduzir as diferenças genéticas entre populações (Relethford, 2012; Silva, 2014).
1.5.2. LINKAGE (DIS)EQUILIBRIUM
Os segmentos de ADN presentes em determinado cromossoma, em teoria, encontram-se em associação e são transmitidos à descendência em bloco - linkage -. O
linkage equilibrium (LE) ocorre quando o genótipo presente em determinado locus é
independente do genótipo de um segundo locus.
Sabe-se que o linkage disequilibrium (LD) pode corresponder à associação não aleatória de alelos em dois ou mais loci diferentes, ou seja, ocorre quando dois genótipos em determinado loci não são independentes um do outro, medindo o grau em que os alelos em dois ou mais loci se encontram associados (Ardlie, Kruglyak e Seielstad, 2002; Flint-Garcia, Thornsberry e Buckler, 2003; Pritchard e Przeworski, 2001; Relethford, 2012; Szibor, 2007). Este termo foi usado pela primeira vez em 1960 por Lewontin e Kojima e continua a ser utilizado atualmente.
Inicialmente, o LD era uma preocupação dos geneticistas da área da genética populacional, uma vez que existiam poucos dados para o estudar. Fora da genética populacional a sua importância para a biologia evolutiva não era reconhecida. As duas principais razões para o não reconhecimento eram o facto das associações não-aleatórias dos alelos em dois loci poderem ocorrer mesmo se os dois genes não estivessem associados e também devido ao facto de dois loci estarem ligados não ser sinónimo de desequilíbrio de ligação. No entanto, o interesse pelo LD cresceu rapidamente na década de 1980, uma vez que, a utilidade do LD para o mapeamento genético se tornou evidente e a identificação de
loci intimamente ligados se tornou viável (Goldstein, 2001; Slatkin, 2008).
Existem diversos fatores que levam ao LD e que se podem enumerar: seleção natural, mutação, deriva genética e fluxo de genes, embora a seleção natural seja o mais importante, uma vez que, pode criar níveis consideráveis de desequilíbrio (Weiss e Clark, 2002).
1.5.3. APONTAMENTO SOBRE MÉTODOS ESTATÍSTICOS E ESTUDO DE POPULAÇÕES
A história demográfica do Homem moderno é composta por uma combinação de fenómenos, tais como, eventos de colonização, migrações recorrentes, mudanças ao nível do tamanho da população, entre outros (Henn, Cavalli-Sforza e Feldman, 2012; Palstra, Heyer e Austerlitz, 2015). A combinação entre os dados genéticos obtidos através do estudo de determinados marcadores genéticos e, entre, métodos estatísticos específicos pode permitir-nos a compreensão e inferência da história demográfica de determinada população (Hellenthal
et al., 2014).
A maioria dos métodos estatísticos desenvolvidos são baseados no princípio da probabilidade (Bertorelle, Benazzo e Mona, 2010). Entende-se por probabilidade a hipótese que determinado fenómeno tem de ocorrer em função de dois ou mais fenómenos possíveis nas mesmas circunstâncias. A probabilidade assenta em dois princípios fundamentais, o primeiro afirma que a probabilidade de dois ou mais eventos independentes ocorrerem em conjunto é igual ao produto das probabilidades de ocorrerem separadamente e, o segundo, afirma que a ocorrência de dois eventos que se anulam é igual à soma das probabilidades com que cada evento ocorre.
Assim, o principal objetivo destes métodos estatísticos é encontrar uma resposta para a probabilidade, através da simulação dos dados genéticos, uma vez que continua a ser um tema complexo quando nos deparamos com fenómenos evolutivos (Palstra, Heyer e Austerlitz, 2015).
Um modelo estatístico adequado para o estudo da evolução do Homem moderno não é importante apenas para compreender a sua história mas também para entender os efeitos da demografia e dos factores que influenciam o equilíbrio de determinada população (Fagundes
et al., 2007).
Os métodos estatísticos, por sua vez, desencadearam o desenvolvimento de softwares, que visam esclarecer a estrutura de determinada população, responder a questões como quanto tempo uma população levou até se modificar, a decifrar os diversos fenómenos de evolução que esta sofreu, tais como movimentos migratórios, tráfico de escravos, invasões, entre outros, através dos dados genéticos obtidos.
O software diyABC (Bertorelle, Benazzo e Mona, 2010; Blum e Jakobsson, 2011; Palstra, Heyer e Austerlitz, 2015) e o software DIYABC (Cornuet, Ravigné e Estoup, 2010),
utilizam o modelo bayesiano aproximado de computação (ABC) para decifrar a história demográfica de determinada população. O modelo bayesiano tem como base dois princípios fundamentais, o primeiro afirma que a probabilidade é uma abordagem ao desconhecido, ou seja, a probabilidade é algo incerto que apenas pode ser decifrado através de uma distribuição de probabilidades. Quanto ao segundo princípio este diz respeito aos dados que são conhecidos, que visam aperfeiçoar o conhecimento do desconhecido de modo a se obter uma resposta.
Para além dos softwares mencionados anteriormente, os softwares simcoal2 e fastsimcoal, permitem recombinar sequências de ADN e, por sua vez, a criação de determinados cenários demográficos, calcular distâncias entre populações, determinar as taxas de migração entre populações e eventos históricos sofridos por essas mesmas populações (Excoffier e Foll, 2011). Por último, o software GLOBETROTTER, que utiliza a informação obtida através de SNP para a determinação de ancestrais que sejam partilhados com outras populações (Hellenthal et al., 2014; Leslie et al., 2015).
Concluindo, existem centenas de outros softwares para além dos mencionados, utilizados com dados obtidos a partir de marcadores autossómicos e de linhagem, que podem ser empregues para a obtenção de respostas acerca da evolução do Homem moderno.
1.6.GENÉTICA FORENSE
O desenvolvimento e a aplicação da Genética revolucionaram a área Forense. No campo da Criminalística Biológica diversas são as perícias médico-legais solicitadas e muitas delas são requeridas à Genética Forense, como por exemplo, a simples obtenção de um perfil genético, quer de amostras recolhidas das vítimas, quer de amostras de indivíduos suspeitos como também de amostras encontradas num local de crime de modo a obter uma relação entre vítimas e suspeitos.
Durante os anos 60 e 70, com o desenvolvimento da Biologia Molecular, principalmente com a descoberta das enzimas de restrição, da Sequenciação de Sanger e do
Southern Blotting, passou a ser possível a análise de sequências de ADN. A partir de 1984,
Alec Jeffreys, mostrou o potencial na aplicação forense da Genética através da análise das regiões polimórficas do ADN, DNA Fingerprinting, comummente conhecido como as impressões digitais do ADN. A partir da resolução de diversos casos criminais com a
implementação desta técnica, esta começou a ser aceite para o uso forense e atualmente tem um papel fundamental a nível mundial (Goodwin, Linacre e Hadi, 2011).
Assim, pode-se definir a Genética Forense como a área das Ciências Forenses que utiliza os conhecimentos da Genética e da Biologia Molecular no apoio à administração da Justiça de modo a resolver casos que se encontram sob investigação.
Apesar da Identificação Humana através do ADN ser o ramo mais desenvolvido da Genética Forense e a sua aplicação mais frequente ser a investigação de parentesco, a Genética Forense não se limita apenas a isto, podendo mesmo ser aplicada a vários níveis da criminalística biológica. A análise do ADN é frequentemente utilizada para adquirir informação a partir de material biológico com o objetivo de associar suspeitos e vítimas (Oorschot, Ballantyne e Mitchell, 2010).
Podem considerar três grandes grupos de perícias da Genética Forense, a investigação de parentesco biológico, a identificação de desconhecidos e a criminalística biológica (Baker e Dalebout, 2009; Zhivotovskiĭ, 2006).
1.7.MARCADORES GENÉTICOS EM GENÉTICA FORENSE E EM GENÉTICA POPULACIONAL
O genoma humano é constituído por sequências repetitivas (52%) e por sequências de cópia única (48%) (figura 8).
As sequências repetitivas incluem elementos genéticos móveis (44%), sequências simples repetidas (3%) e duplicações segmentares (5%). Por sua vez, os elementos genéticos móveis incluem Long Interspersed Elements - LINEs - (20%), Short Interspersed Elements - SINEs - (13%), Long Terminal Repeats - LTRs - (8%) e outras sequências de ADN (3%).
Relativamente às sequências de cópia única estas incluem ADN não repetitivo (26,5%) e genes (21,5%) que incluem intrões (20%) e regiões codificantes de proteínas (1,5%).
O genoma humano contém múltiplas sequências repetitivas, que existem em diversos tamanhos e que, habitualmente, são designadas pelo número de repetições da unidade de repetição (Alberts, 2008).
A identificação genética individual é o modo de se estabelecer a identidade de um indivíduo a partir de amostras biológicas (Pinheiro, 2010). A identificação genética individual é realizada, maioritariamente com recurso aos STR, uma vez que estes são marcadores bastante informativos, no entanto, em alguns casos mais complexos, a análise destes marcadores não é suficiente para se alcançar uma identificação individual positiva, sendo necessário o estudo do ADN mitocondrial (ADNmt) ou o estudo de outro tipo de polimorfismo de ADN, como por exemplo, SNP.
Os polimorfismos referidos correspondem a variações ao nível da sequência e ao nível do tamanho da molécula de ADN. Assim basicamente existem duas formas de polimorfismos, os polimorfismos de sequência e os polimorfismos de comprimento. Os polimorfismos de sequência consistem numa troca de uma base por outra e os polimorfismos de comprimento são variações na molécula de ADN como consequência de uma deleção ou inserção de uma ou mais bases, bem como no número de unidades de repetição de um determinado motivo ou sequência.
1.7.1. MARCADORES DO TIPO STR
Os marcadores do tipo short tandem repeat (STR) são os marcadores de eleição na rotina forense, uma vez que preenchem requisitos indispensáveis para serem utilizados como marcadores forenses, designadamente, são extremamente sensíveis, possuem elevado grau de polimorfismo, são muito discriminativos e estão disponíveis em kits que requerem quantidades mínimas de ADN e podem ser facilmente amplificados mediante a reação em cadeia da polimerase, PCR.
Os STR são também conhecidos como microssatélites e consistem em pequenos blocos de nucleótidos que se repetem em tandem, com cerca de 2 a 7 pares de base (pb). O número de repetições varia, geralmente, entre 2 e 40, ou seja, repetem-se em tandem de 2 a 40 vezes. Os STR são designados conforme o tamanho da unidade de repetição e podem ser mono-, di-, tri-, tetra-, penta- ou mesmo hexanucleotídicas, sendo que as repetições de blocos tetranucleotídicos são os mais utilizados na identificação humana (Butler, 2012), pois as suas
15% do alelo real, enquanto nos casos de blocos com unidades di- e tri- nucleotídicas as
stutters apresentam picos com altura que chega a cerca de 30% ou mais do alelo real,
dificultando assim a interpretação nos casos em que ocorram misturas de material biológico (Butler, 2010, 2012).
Existem STR autossómicos e STR nos cromossomas sexuais, STR do cromossoma Y e STR do cromossoma X. Normalmente em estudos forenses são utilizados os marcadores autossómicos, a razão mais óbvia para a sua utilização é devido ao facto de serem utilizados na identificação de indivíduos quer do sexo masculino quer do sexo feminino (Ramachandran
et al., 2004). Quanto aos marcadores dos cromossomas sexuais, são herdados num conjunto
designado por haplótipo, devido ao facto de se localizarem muito próximos uns dos outros, não segregando de uma forma independente (Buckleton, Krawczak e Weir, 2011).
Apesar de possuírem inúmeras vantagens também possuem algumas limitações, mais significativas, por exemplo, aquando da utilização em misturas de amostras, como é o caso de amostras provenientes de crimes sexuais, em que há uma probabilidade elevada de ocorrer uma mistura de perfis. Nestes casos os STR do cromossoma Y, são uma boa alternativa, uma vez que estes marcadores produzem apenas perfil dos indivíduos do sexo masculino (Leat et
al., 2004). Relativamente aos STR do cromossoma X, estes são úteis em casos de
investigação de maternidade e em casos de paternidade complexos, em que o pretenso pai se encontra ausente, sendo importante ter em conta que o poder de discriminação destes varia com o género ao qual pertence a amostra biológica (Pinheiro, 2013).
Em suma, os STR autossómicos são utilizados pela Genética Forense, uma vez que, possuem um poder de discriminação elevado, encontram-se em diferentes cromossomas; e quando se encontrarem no mesmo cromossoma são selecionados os que se localizam distantes uns dos outros (Imad et al., 2014). Para além destas características apresentam elevada robustez e reprodutibilidade dos resultados quando se realiza um ensaio em multiplex e apresentam baixa taxa de mutação.
1.7.2. MARCADORES DO TIPO SNP
Os marcadores do tipo Single Nucleotide Polymorphisms (SNP) envolvem tipicamente substituições de uma base nucleotídica na sequência de ADN, resultante de uma troca, deleção ou inserção de um nucleótido, surgindo de uma mutação pontual e são considerados como polimorfismos de posição (figura 9).
Apesar do número elevado de SNP existentes no genoma humano, 1 SNP por cada 200 a 300 pares de base, a taxa de mutação destes é inferior à dos marcadores do tipo STR, daí que possam ser utilizados para distinguir populações, para definir a história geográfica de determinada população, sendo também utilizados em identificação humana (Costa et al., 2008; Pinheiro, 2010; Vignal et al., 2002).
Figura 9: Exemplo de um SNP em que é possível observar que a sequência de ADN 1 é diferente da sequência de ADN 2, num único par de base, C-G e T-A (Fonte: Pinheiro, 2010).
Os SNP, em termos forenses, são utilizados principalmente para obter informação de amostras em que o ADN se encontra degradado, uma vez que, apenas é necessário uma pequena região alvo do ADN, pois o tamanho dos produtos amplificados tem menos de 100 pb, tratando-se de um estudo mais viável do que o estudo de STR. Apenas é estudado um único nucleótido com este tipo de marcadores, em vez de uma quantidade elevada de nucleótidos como nos STR.
Uma das vantagens que os SNP apresentam é taxa de mutação mais baixa que os STR, cerca de 100 000 vezes mais baixa, sendo assim possível afirmar que os SNP são mais estáveis em termos de transmissão à descendência, o que é útil na Genética Forense. Para além disso, podem ser também amplificados em multiplex e não produzem stutters, ou artefactos de amplificação. Por fim, podem ser utilizados para a determinação da origem étnica do indivíduo a que pertence a amostra (Amorim e Pereira, 2005; Costa et al., 2008; Pinheiro, 2010).
Para além das vantagens da utilização deste tipo marcadores, quando comparados com os STR, possuem diversas desvantagens, tal como o número de loci estudados necessários,