Modelação baseada em agentes para a simulação do uso do
espaço e efeitos causados pela instalação de um parque
eólico sobre Nyctalus leisleri
Dissertação de Mestrado em Engenharia do Ambiente
Daniel Machado Ferreira
Orientador
Professor Doutor Mário Gabriel Santiago dos Santos
Coorientador
Professor Doutor João Alexandre Ferreira Abel dos Santos Cabral
Modelação baseada em agentes para a simulação do uso do
espaço e efeitos causados pela instalação de um parque
eólico sobre Nyctalus leisleri
Dissertação de Mestrado em Engenharia do Ambiente
Daniel Machado Ferreira
Orientador
Professor Doutor Mário Gabriel Santiago dos Santos
Coorientador
Professor Doutor João Alexandre Ferreira Abel dos Santos Cabral
Dissertação apresentada para obtenção do grau de Mestre em Engenharia do Ambiente, pela Universidade de Trás-os-Montes e Alto Douro.
“The essence of the individual-based approach is the derivation of the properties of ecological systems from the properties of the individuals constituting these systems.”
Agradecimentos
Ao concluir a minha dissertação, não posso deixar de agradecer a todos os que, de alguma forma, contribuíram para a sua realização:
Ao professor Mário Santos o meu obrigado por toda a orientação, conhecimento e incentivo, que me transmitiu ao longo de todos estes meses, assim como pela sua permanente disponibilidade e empenho demonstrado nas sugestões que trouxe para a melhoria do desenvolvimento desta dissertação.
Ao professor João Alexandre Cabral por aceitar coorientar este trabalho e por todas as sugestões e conhecimento partilhado.
À Regina Santos pela ajuda preciosa com todo o processo de tratamento de informação geográfica, pela simpatia e enorme disponibilidade para ajudar neste estudo.
Ao Paulo Barros, pela boa vontade na cedência de informação, bibliografia e fotos, compartilhando comigo um pouco dos seus conhecimentos sobre quirópteros.
Ao Paulo Travassos, pela simpatia e disponibilidade na cedência de bibliografia e partilha de conhecimentos.
A toda a equipa do LEA pela simpatia que sempre manifestaram.
Ao Nuno, Diana, Ricardo, Sílvio, Cátia, Fiorenzo e Tânia, pela grande ajuda que deram durante o moroso processo das simulações, e por tornarem esta fase mais divertida com a vossa boa disposição e companheirismo.
A todos os amigos, pelas palavras de apoio e motivação, sem vocês tudo teria sido bem mais difícil.
Aos meus pais e irmão, agradeço especialmente pelo permanente estímulo que me deram ao longo de toda a minha vida e a segurança que me transmitem, acreditando sempre em mim.
Aos meus sobrinhos pela sua alegria natural que me proporcionou momentos valiosos de grande descontração.
À Cláudia por toda a paciência, apoio e companheirismo que me deu durante todo o desenvolvimento deste trabalho, que acabou por coincidir com uma fase marcante para as nossas vidas.
Resumo
Tendo em conta os esforços a nível mundial para o desenvolvimento de métodos sustentáveis de produção de energia, tem-se verificado um crescente desenvolvimento das tecnologias associadas a uma produção através de fontes renováveis, com especial destaque para a energia eólica. Apesar dos claros benefícios ambientais associados a este tipo de tecnologia, diversos estudos têm vindo a confirmar a mortalidade de morcegos em parques eólicos, o que levanta preocupações sobre os efeitos a longo prazo, da presença destas estruturas, sobre as suas populações. Nyctalus leisleri é uma espécie migradora particularmente afetada pela presença de parques eólicos. Neste estudo desenvolveu-se um Modelo Baseado em Agentes, que constitui uma ferramenta preditiva de carácter conservacionista, com enfoque nos padrões de uso do espaço em período de alimentação, bem como no possível impacte de um parque eólico sobre esta espécie. O modelo desenvolvido foi aplicado a uma área centrada no parque eólico de Negrelo e Guilhado, localizado no noroeste de Trás-os-Montes.
Identificou-se uma clara predominância de uso do espaço em zonas identificadas como propícias à alimentação e uma elevada mobilidade evidenciada pelos resultados ao nível da distância máxima percorrida em relação ao abrigo (média = 8,63 km) e das dimensões da área vital (média = 3,59 km2). A área usada de florestas de resinosas, florestas de folhosas e outros usos constituíram os fatores mais relevantes para o incremento das dimensões da área vital. Foi possível constatar que velocidades do vento elevadas (≥ 4 m/s) levam à diminuição do número de fatalidades devido às limitações que estas impõem ao nível da atividade. Observou-se uma clara relação entre o aumento da frequência dos eventos de mortalidade e a proximidade do abrigo ao aerogerador mais próximo. Os aerogeradores instalados em usos do solo identificados como habitats de alimentação apresentam maiores índices de mortalidade, tendo-se constatado uma correlação positiva, entre os índices de atividade próxima dos aerogeradores e a ocorrência de mortalidade.
Palavras-Chave: Nyctalus leisleri, parques eólicos, modelação baseada em agentes, morcegos, mortalidade.
Abstract
Agent-based modeling to simulate the use of space and effects caused by the installation of a wind farm on Nyctalus leisleri
Given the worldwide efforts to develop sustainable methods of energy production, there has been an increasing development of technologies associated with energy production from renewable sources, with special emphasis on wind energy. Despite the clear environmental benefits associated with this type of technology, many studies have confirmed bat fatalities at wind farms, which raise concerns about the long-term effects of these structures on bat populations. Nyctalus leisleri is a migratory species particularly affected by the presence of wind farms. For this study I developed an Agent-Based Model, which constitutes a predictive tool of conservative nature, with focus on space use patterns, as well as the possible impact of a wind farm on this species. The model was applied to an area centered on the Negrelo and Guilhado wind farm located in the northwest of Trás-os-Montes.
A clear predominance in the selection of foraging areas and high mobility was evidenced by the results of maximum distance to roosts (mean = 8.63 km) and home range size (mean = 3.59 km2). The use of coniferous woodland, deciduous woodland and other land cover were the most important factors for increasing the home range size. It was found that high wind speeds (≥ 4 m/s) lead to a decrease in the number of fatalities due to the limitations they impose in bat activity. There was a clear relationship between increased frequency of mortality events and the proximity between roosts and turbines. The wind turbines installed on areas identified as foraging habitats have higher rates of mortality, and there is a positive correlation between the activity levels near the wind turbines and the occurrence of fatalities.
Índice
Agradecimentos ... I Resumo ... II Abstract ... III Índice ... IV Índice de Tabelas ... VI Índice de Figuras ... VIII1. Introdução ... 1
1.1. Morcegos - Ecologia e Conservação ... 1
1.2. Nyctalus leisleri (Kuhl, 1817) ... 4
1.3. Breve Abordagem às Alterações Climáticas ... 9
1.4. Energia Eólica – Situação Atual ... 11
1.5. Vantagens e Desvantagens da Energia Eólica ... 13
1.6. Impacte dos Parques Eólicos nos Morcegos ... 16
1.7. Modelação Baseada em Agentes ... 18
2. Objetivos ... 20 3. Material e Métodos ... 21 3.1. Área de Estudo ... 21 3.2. Procedimento de Modelação ... 23 3.2.1. Ambiente Netlogo ... 23 3.2.2. Protocolo ODD ... 24 3.2.2.1. Vista Geral ... 24 3.2.2.1.1.Objetivo... 24
3.2.2.1.2.Variáveis de Estado e Escalas ... 25
3.2.2.1.3.Visão Geral dos Processos e Sequência ... 27
3.2.2.1.3.1.Seleção dos abrigos ... 27
3.2.2.1.3.2.Estrutura dos abrigos ... 28
3.2.2.1.3.3.Primeiro período de atividade e condicionantes à emergência .... 28
3.2.2.1.3.4.Comportamento em período de alimentação ... 30
3.2.2.1.3.5.Eventos de mortalidade... 31
3.2.2.1.3.6.Regresso aos abrigos ... 32
3.2.2.1.3.7.Segundo período de atividade e condicionantes à emergência ... 32
3.2.2.2. Conceitos de Design ... 34 3.2.2.2.1.Fenómenos Emergentes ... 34 3.2.2.2.2.Adaptação ... 34 3.2.2.2.3.Fitness ... 35 3.2.2.2.4.Deteção ... 35 3.2.2.2.5.Interação ... 36 3.2.2.2.6.Estocasticidade ... 36 3.2.2.2.7.Fenómenos Coletivos ... 38 3.2.2.2.8.Observação ... 38 3.2.2.3. Especificações ... 38 3.2.2.3.1.Inicialização ... 38
3.2.2.3.2.Valores iniciais de entrada ... 42
3.2.2.3.3.Submodelos ... 43
4. Resultados e Discussão ... 46
4.1. Utilização do espaço ... 46
4.2. Mortalidade - Variáveis ambientais e ecologia de N. leisleri ... 56
4.3. Mortalidade - Variáveis ambientais e configuração do parque eólico ... 61
5. Considerações Finais ... 68
5.1. Conclusões ... 68
5.2. Considerações sobre o modelo e recomendações metodológicas ... 70
6. Referências ... 72 6.1. Referências Bibliográficas ... 72 6.2. Sítios consultados... 81 6.3. Documentos legais ... 81 Anexos ... …82 Anexo I………...……….…….82 Anexo II...………...………. 84 Anexo III……….…... 91
Índice de Tabelas
Tabela 1 – Descrição das variáveis de cada entidade presente no modelo. ………….. 26 Tabela 2 – Intervalos que determinam a distribuição dos morcegos pelos locais de abrigo no início de cada simulação. ……… 28 Tabela 3 – Intervalos que determinam o número de emergências para alimentação. .. 29 Tabela 4 – Combinação de variáveis testadas nos cenários de simulação. …………... 42 Tabela 5 – Dados usados para a calibração da variável Seek. ………. 43 Tabela 6 – Disponibilidade de cada tipo de habitat na área de estudo (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos). ……….… 49 Tabela 7 – Valores de q do Teste de Comparações Múltiplas não paramétrico (tipo - Tukey), relativo à relação T/P registada para cada tipo de habitat (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos) (n=100; valor q de referência = 4,387) e respetivo nível de significância (ns não significativo; * < 0,05)………. 51 Tabela 8 – Teste de Spearman entre o número de patches usados e a quantidade de
ticks de permanência em cada tipo de habitat (n=100) (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos) e respetivo nível de significância (*** P <0,001). ……….. 52 Tabela 9 – Valores médios de área vital e distância máxima ao abrigo e respetivos desvios-padrão. ……… 53 Tabela 10 – Equação de regressão para determinação da influência da quantidade de patches usados de cada habitat nas dimensões da área vital (n=100) e respetivos graus de liberdade (g.l.), coeficiente de determinação (R2 ajustado), critério de informação de Akaike (AIC), valor de F e respetivo nível de significância (*** P <0,001). Legenda: AV – área vital, AFR – área usada de floresta de resinosas, AP - área usada de pastagens, ATU - área usada de tecido urbano, AFF - área usada de florestas de folhosas, AOU - área usada de outros usos, ALF - área usada de limites florestais, AEF - área usada de estradas florestais. ……… 55 Tabela 11 – Caracterização dos aerogeradores consoante a sua altimetria e uso de solo da zona de implantação (P- pastagens e OU – outros usos). ……… 61 Tabela 12 – Caracterização dos aerogeradores consoante a sua distância aos diversos tipos de uso do solo presentes na área de estudo (FM – florestas mistas, FR – florestas de resinosas, P – pastagens, TU – tecido urbano, FF – florestas de folhosas, OU – outros usos, LF – limites florestais, CA – cursos de água, EF – estradas florestais). ……… 64 Tabela 13 – Caracterização dos aerogeradores consoante a composição do uso do solo na área envolvente, com raio de 1,5 km (FM – florestas mistas, FR – florestas de resinosas, P – pastagens, TU – tecido urbano, FF – florestas de folhosas, OU – outros usos, LF – limites florestais, CA – cursos de água, EF – estradas florestais). … 64
Tabela 14 – Teste de Kruskal-Wallis para a atividade registada no espaço em torno de cada aerogerador (raio de 100 m), para as duas classes de temperatura aos 1000m (n=1200), e respetivo nível de significância (* p <0,05; *** p < 0,001). ………... 65 Tabela 15 – Valores de q do Teste de Comparações Múltiplas não paramétrico (tipo - Tukey), relativo ao nível de atividade em torno dos aerogeradores (AG), a temperaturas iguais a 10ºC, aos 1000 m (n=120; valor q de referência = 4,474) e respetivo nível de significância (ns não significativo; * < 0,05). ……… 67 Tabela 16 – Valores de q do Teste de Comparações Múltiplas não paramétrico (tipo - Tukey), relativo ao nível de atividade em torno dos aerogeradores (AG), a temperaturas superiores a 10ºC, aos 1000 m (n=120; valor q de referência = 4,474) e respetivo nível de significância (ns não significativo; * < 0,05). ……… 67
Índice de Figuras
Figura 1 – Nyctalus leisleri (foto de Paulo Barros). ………... 4 Figura 2 – Distribuição espacial de N. leisleri (Fonte: IUCN, 2012). ………... 5 Figura 3 – Trajeto típico de voo em período de alimentação de N. leisleri (Fonte: Russ, 1999). ………... 8 Figura 4 – Penetração de energia eólica no consumo total de eletricidade dos países europeus até 2011 (Fonte: EWEA, 2012). ………... 11 Figura 5 – Localização e potência dos parques eólicos do território continental português (Fonte: APREN, 2011). ………. 12 Figura 6 – Evolução do tamanho dos aerogeradores comerciais (Fonte: http://www.dewi.de). ……….... 15 Figura 7 – Localização da área de estudo em Portugal Continental (A). Localização do parque eólico do Negrelo e Guilhado, dentro da área de estudo (B). Pormenor do parque eólico do Negrelo e Guilhado (C) Os círculos indicam a localização dos aerogeradores. …... 21 Figura 8 – Interface principal de um cenário criado no ambiente NetLogo. …………... 23 Figura 9 – Visão geral do processo de simulação. Seleção dos abrigos (A); Estrutura dos abrigos (B); Primeiro período de atividade e condicionantes à emergência (C); Comportamento em período de alimentação (D); Eventos de mortalidade (E); Regresso aos abrigos (F) e Segundo período de alimentação e condicionantes à emergência (G). ……… 33 Figura 10 – Distribuição da variável Soil-Use pelo mundo virtual (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos). Os aerogeradores encontram-se representados pelos ícones presentes no modelo... 39 Figura 11 – Distribuição da variável Altitude pelo mundo virtual. Os aerogeradores encontram-se representados pelos ícones presentes no modelo. ……….. 40 Figura 12 – Posição relativa dos aerogeradores em relação aos patches com risco de mortalidade (A). Distribuição dos valores da variável Risk pelos patches abrangidos pela área de influência do aerogerador (B). ……… 41 Figura 13 – Mapa cumulativo de utilização do espaço. Considerou-se utilização elevada a partir de 50 ticks de permanência. Os aerogeradores encontram-se representados pelos ícones presentes no modelo. ……… 47 Figura 14 – Percentagem de patches com utilização elevada (mais de 50 ticks de utilização), para cada tipo de habitat (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos) ………….. 48
Figura 15 – Relação média de T/P para cada tipo de habitat (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos), com representação do desvio-padrão. ………... 50 Figura 16 – Distribuição da área vital em função da distância máxima ao abrigo. …….. 53 Figura 17 – Representação de três exemplos de áreas vitais calculadas a partir do polígono convexo mínimo (A - área pequena, B - área média e C - área grande), sobre o mapa de usos do solo (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos). ………... 54 Figura 18 - Valor de t e nível de significância (* P<0,05; ** P<0,01; *** P<0,001) das variáveis independentes (AFR – área usada de floresta de resinosas, AP - área usada de pastagens, ATU - Área usada de tecido urbano, AFF - área usada de florestas de folhosas, AOU - área usada de outros usos, ALF - área usada de limites florestais, AEF - área usada de estradas florestais), selecionadas pela regressão múltipla, como fator com influência significativa sobre a área vital (AV). ……….. 56 Figura 19 – Mortalidade média para as duas classes testadas das variáveis Temperatura (A), Velocidade do Vento (B), Harém (C) e Número da Emergência (D), com representação do respetivo erro-padrão. ………. 59 Figura 20 – Localização espacial dos patches de abrigo de indivíduos que sofreram evento de mortalidade, relativamente à linha de distância média entre os abrigos de todos os morcegos e o aerogerador mais próximo. Representação sobre o mapa de usos do solo (FR – florestas de resinosas, FF – florestas de folhosas, FM – florestas mistas, P – pastagens, TU – tecido urbano, LF – limites florestais, CA – cursos de água, EF – estradas florestais, OU – outros usos). ……… 60 Figura 21 – Mortalidade média por aerogerador (AG) para as duas classes testadas de Temperatura (Temp. = 10oC e Temp. > 10oC, aos 1000 m), com representação do respetivo erro-padrão. ………... 63 Figura 22 – Utilização do espaço média por aerogerador (AG) para as duas classes testadas de Temperatura (Temp. = 10oC e Temp. > 10oC, aos 1000 m), com representação do respetivo erro-padrão. ………. 66
1. Introdução
1.1. Morcegos - Ecologia e Conservação
Os morcegos são mamíferos que se incluem na ordem Chiroptera, sendo atualmente conhecidas mais de 1100 espécies (Kunz & Fenton, 2003), o que representa aproximadamente um quarto da riqueza de mamíferos conhecidos (Altringham, 1996). A ordem Chiroptera divide-se tradicionalmente em duas subordens, Megachiroptera e Microchiroptera (Barlow, 1999; Hutson et al., 2001). Os morcegos pertencentes à subordem Megachiroptera, que são também conhecidos por morcegos frugívoros do velho mundo, ou raposas voadoras, encontram-se distribuídos exclusivamente em climas tropicais e subtropicais (Altringham, 1996), enquanto a subordem Microchiroptera, onde se incluem todas as espécies Europeias, apresenta uma distribuição mais ampla, um pouco por todo o mundo, à exceção do Antártico, Ártico e algumas ilhas oceânicas mais isoladas (Hutson, et al., 2001).
Os quirópteros são os únicos mamíferos que possuem a capacidade de voo, para além de um vasto conjunto de características ecológicas que tornam particularmente importante o seu estudo científico (Kunz, 1988). Os microquirópteros desenvolveram ainda um complexo sistema de ecolocalização (Hill & Smith, 1986; Airas, 2003), que permite não só a orientação, mas também a captura de presas, através da emissão de pulsos sonoros, na sua maioria ultrassons, produzidos pela boca e em alguns casos por uma complexa estrutura nasal, que são posteriormente interpretados durante o voo (Hill & Smith, 1986; Moss & Sinha, 2003; Fenton, 2003).
O elevado número de espécies de quirópteros, bem como a sua ampla distribuição, proporciona uma assinalável diversidade ecológica. No que diz respeito ao tipo de abrigo, ocorre uma procura diversificada de espécie para espécie, podendo ainda a mesma espécie utilizar diferentes tipos de abrigos durante o ciclo anual. Os morcegos que habitam zonas tropicais tendem a abrigar-se em zonas mais expostas, tais como ramos de árvores e folhagem. Os morcegos de zonas temperadas usam geralmente abrigos mais resguardados, como é o caso de cavidades subterrâneas, fissuras em rochas, cavidades e orifícios em árvores, podendo haver ainda recurso a estruturas de origem antropogénica, de onde se destacam minas, pontes e edifícios (Altringham, 1996, Hutson et al., 2001).
A alimentação dos quirópteros também é consideravelmente variada, visto que estes podem ser insectívoros, carnívoros, piscívoros, frugívoros, hematófagos, nectarívoros, ou polinívoros (Altringham, 1996). A própria forma como se alimentam diverge de acordo com a espécie, nomeadamente em morcegos insectívoros, como é o caso de todas as espécies que podem ser encontradas no território português, nas quais se pode observar uma ampla
variedade de comportamentos associados à caça (Schober & Grimmberger, 1996). Consoante a espécie, os morcegos podem usar como zonas preferenciais de caça, galerias ripícolas, massas de água, zonas florestais, áreas abertas, zonas agrícolas, áreas de mato, zonas de bordadura e até mesmo áreas urbanas, diversificando-se também as estratégias de captura das presas consoante o tipo de habitat onde caçam (Russ & Montgomery, 2003).
O tipo de dieta alimentar confere aos quirópteros um papel indispensável no funcionamento dos ecossistemas. As espécies insectívoras podem chegar a consumir cerca de metade do seu peso corporal em insetos durante uma noite de caça, o que representa um consumo de dezenas de toneladas de insetos por noite, tornando-os controladores das populações de insetos, incluindo espécies potencialmente prejudiciais para o ser humano, como é o caso das pragas agrícolas e vetores de doenças (Palmeirim & Rodrigues, 1992; Altringham, 1996; Kiser & Kiser, 2002). É ainda assinalável a importância que algumas espécies possuem como dispersores de sementes ou como polinizadores, com um papel preponderante em processos de recuperação e dinâmica de ecossistemas (Altringham, 1996).
A importância ecológica dos morcegos não se resume aos seus hábitos alimentares, sendo que as espécies cavernícolas, dependentes de abrigos subterrâneos em pelo menos uma das fases do seu ciclo de vida, representam uma via de introdução de matéria orgânica em ecossistemas subterrâneos, através do guano (fezes de morcego). Desta forma é garantido o suporte de comunidades de invertebrados que existem exclusivamente nestes meios (Palmeirim & Rodrigues, 1992). Existe ainda a vertente económica ligada à exploração das elevadas quantidades de guano acumuladas sob as colónias de morcegos, como forma de fertilizante agrícola (Altringham, 1996).
Algumas características biológicas únicas apresentadas pelos morcegos têm vindo a ser estudadas no campo da investigação médica, nomeadamente o seu mecanismo de termorregulação, bem como as propriedades anticoagulantes da saliva de alguns morcegos hematófagos, com potencialidades ao nível das doenças coronárias (Hill & Smith, 1986; Altringham, 1996; Liberatore et al., 2003).
As populações de morcegos enfrentam atualmente um declínio generalizado, em diversas partes do mundo (Altringham, 1996), sendo que quase 16% das espécies conhecidas se encontram extintas ou listadas com estatuto de ameaça (criticamente em perigo, em perigo ou vulnerável), no entanto, poderá existir uma subestimação das espécies ameaçadas, devido à preocupante falta de dados relativos a diversas espécies (IUCN, 2012).
Existem fatores intrínsecos que podem agravar as consequências das ameaças externas, tais como a sua maturidade sexual tardia, que para algumas espécies apenas é
atingida ao quarto ano de vida, e a sua baixa taxa de natalidade, visto que as fêmeas da maioria das espécies apenas muito raramente têm mais que uma cria por ano (Palmeirim & Rodrigues, 1992; Racey & Entwistle, 2003). Por outro lado, as principais ameaças para os quirópteros prendem-se com a destruição e alteração global dos ecossistemas, que resultam no decrescimento das suas fontes alimentares. O facto de, por vezes, se aglomerarem em colónias numerosas, como é o caso de colónias de maternidade ou de hibernação, torna-os particularmente vulneráveis à destruição ou perturbação dos locais de abrigo (Palmeirim & Rodrigues, 1992; Altringham, 1996). É ainda de destacar uma série de outras ameaças, tais como o uso excessivo de pesticidas, a perseguição e mais recentemente os registos de mortalidade em parques eólicos ou causada por colisão com veículos (Altringham, 1996; Hutson et al., 2001; Cabral et al., 2006).
Em Portugal estão presentes 27 espécies de morcegos, todas pertencentes à subordem Microchiroptera, das quais 25 ocorrem no território continental, para além das espécies que ocorrem apenas nos arquipélagos dos Açores e Madeira, como é o caso de Nyctalus azoreum, endémica dos Açores e Pipistrellus maderensis, endémica da Macaronésia (ICNB, 2012). Das espécies que ocorrem em Portugal continental, 36% possuem estatuto de ameaça (três criticamente em perigo, uma em perigo e cinco vulneráveis), no entanto existe ainda grande falta de dados relativos a muitas espécies, o que se reflete nos 36% de espécies com informação insuficiente (Cabral et al., 2006).
Devido às suas caraterísticas biológicas únicas e importância ecológica, a conservação das populações de morcegos tornou-se uma questão extremamente relevante e neste sentido todas as espécies que ocorrem em Portugal estão protegidas por legislação nacional e internacional, nomeadamente a Convenção de Berna, de 1982 (Decreto-Lei nº 316/89, de 22 de Setembro); a Convenção de Bona, de 1983 (Decreto nº103/80, de 11 de Outubro); a Directiva Habitats, de 1992 (Decreto-Lei nº140/99, de 24 de Abril) e o Acordo para a Conservação das Populações Europeias de Morcegos - EUROBATS, de 1994 (Decreto-Lei nº 31/95, de 18 de Agosto).
1.2. Nyctalus leisleri (Kuhl, 1817)
O morcego-arborícola-pequeno, Nyctalus leisleri (Kuhl, 1817), pertencente à família Vespertilionidae, é um morcego migratório de tamanho médio, apesar de ser a espécie de menores dimensões do género Nyctalus presente na Europa continental (Palmeirim, 1990). Apresenta uma envergadura de asas de 260 a 320 mm, com um comprimento total que pode ir dos 83 aos 113 mm e um peso que pode variar entre os 11 e os 20 g. O seu dorso exibe uma tonalidade castanho-escura enquanto a tonalidade do ventre é mais clara. A sua pelagem é claramente bicolor, com a parte basal dos pelos a apresentarem coloração mais escura que as extremidades. Por sua vez, as suas orelhas, face e membranas alares exibem tons de castanho-escuro ou preto (Figura 1). As suas orelhas são arredondadas e possuem tragus em forma de cogumelo, característico do género Nyctalus (Palmeirim, 1990; Schober, 1997; Stebbings, 1986; Stebbings, 1988).
Figura 1 – Nyctalus leisleri (foto de Paulo Barros).
Poucos dados existem acerca da longevidade de N. leisleri, havendo registo de indivíduos que atingiram os 12 anos (Schorcht, 2009). No entanto, outros morcegos da família Vespertilionidae podem viver dezenas de anos, por isso esta estimativa de longevidade poderá constituir uma subestimação dos valores reais (Schober, 1997).
Esta espécie distribui-se predominantemente pelo Paleártico ocidental (Figura 2), ocorrendo em grande parte da Europa. A sua distribuição vai desde o Mar Mediterrâneo até ao Norte da Irlanda, Inglaterra e Centro da Escandinávia, e desde Portugal até ao mar Cáspio, Rússia e Irão (Schober, 1997; Stebbings, 1986; Stebbings, 1988), ocorrendo ainda
nas Ilhas Canárias (Trujillo, 1991). Relativamente a Portugal, parece ser progressivamente mais rara de Norte para Sul (Palmeirim et al., 1999), encontrando-se também presente na Ilha da Madeira, onde é representada pela subespécie endémica N. leisleri verrucosus (Palmeirim, 1991).
Figura 2 – Distribuição espacial de N. leisleri (Fonte: IUCN, 2012).
As migrações representam uma parte essencial nos ciclos de vida de uma fração considerável da fauna animal. Nos vertebrados, as migrações constituem um movimento sazonal de um habitat para outro que ofereça condições energéticas mais vantajosas, ou de forma a evitar condições climatéricas desfavoráveis. Neste prisma, os morcegos das zonas temperadas exibem três padrões gerais de comportamento espacial: comportamento sedentário, no qual procriam e hibernam num raio igual ou inferior a 50 km; migrações regionais, nas quais podem percorrem distâncias de 100 a 500 km entre os abrigos de verão e inverno; e migrações de larga escala, nas quais os morcegos podem perfazer distâncias superiores a 1000 km entre os abrigos sazonais (Fleming, 2010).
No género Nyctalus, constituído por espécies de morcegos arborícolas, nos quais parece ser comum a segregação sexual durante a época de procriação (período em que as fêmeas dão à luz e cuidam das crias), existem registos de comportamentos migratórios que podem ir desde o sedentarismo parcial a migrações com diferentes padrões entre machos e fêmeas (Fleming & Eby, 2003). Nestas espécies é comum as fêmeas deslocarem-se 1000 km na primavera em direção às áreas de procriação localizadas em latitudes mais elevadas, enquanto os machos tendem a realizar viagens mais curtas, ou a manter-se nas mesmas áreas durante todo o ano (Cryan 2003; Fleming & Eby, 2003; Strelkov, 1997). Um estudo
sobre a migração de morcegos na Europa incluiu inequivocamente N. leisleri na categoria das espécies migradoras de larga escala (Hutterer et al., 2005), existindo registos de deslocações de 810 km durante períodos migratórios (Nowak, 1994; Schober, 1997).
Esta espécie acasala no final de Agosto e novamente na Primavera. Durante este período os machos defendem os seus abrigos de outros machos, e tentam atrair potenciais parceiras através de vocalizações, constituindo haréns de até nove fêmeas (Schober, 1997). Na época de procriação as fêmeas agregam-se em colónias de maternidade onde dão à luz 1 ou 2 crias uma vez por ano, no final da primavera. Apesar dos escassos dados existentes acerca do desenvolvimento das crias de N. leisleri, sabe-se que estas começam a voar a partir de 30 dias depois do seu nascimento (Schober, 1997; Shiel & Fairley, 1999). As colónias de maternidade tendem a concentrar-se no Norte e Centro da Europa, enquanto os abrigos de Inverno se localizam nas áreas do Sul do continente (Hutterer et al., 2005). A migração por parte das fêmeas desta espécie é aceite como a causa da sua ausência durante a época de procriação nas áreas mais a Sul da Europa e a sua posterior chegada a estas regiões no outono (Roer, 1995; Strelkov, 1969). O estudo de Ibañez et al. (2009), que compila dados de captura de três espécies do género Nyctalus, durante um período superior a 20 anos, indica que no Norte da Península Ibérica os machos de N. leisleri mantêm a sua presença durante todo o ano. Salienta ainda, que as fêmeas ocorrem exclusivamente entre o final de agosto e outubro, verificando-se um aumento na proporção de fêmeas no final do verão - atingindo os 50% no mês de setembro (Ibañez et al., 2009). Este estudo destaca ainda que nenhum juvenil foi capturado durante o período de procriação, o que poderá apontar para a ausência de colónias de maternidade nesta região.
Quase dois terços das espécies Europeias de morcegos usam cavidades em árvores como forma de abrigo durante parte do seu ciclo de vida, sendo que algumas destas espécies dependem deste tipo de abrigo para a sua reprodução e hibernação (Stebbings, 1988; Agnelli et al., 2004; Russo et al., 2004).
N. leisleri é uma espécie florestal associada aparentemente a florestas de folhosas bem desenvolvidas, onde se abriga essencialmente em cavidades de árvores, manifestando uma clara preferência por árvores localizadas em áreas florestais mais antigas, com idades superiores a 100 anos (Ruczyński et al., 2010). Podem no entanto ser encontrados tanto em edifícios antigos como em novos, em sótãos, sob telhas ou em chaminés desativadas (Corbert & Harris 1991; Mitchell-Jones et al., 1999; Shiel et al., 1999; Waters et al., 1999). Utilizam ainda caixas-abrigo para morcegos, quer em florestas de folhosas, quer em florestas de resinosas (Corbet & Harris, 1991).
A variação sazonal na escolha de abrigos, pode refletir as diferenças térmicas do próprio abrigo ao longo do ano. Um abrigo mais quente é importante durante o período
reprodutivo, enquanto as fêmeas necessitam de manter a homeotermia durante a gestação e período de lactação (Racey, 1973; Racey & Swift, 1981; Grinevitch et al., 1995; Altringham, 1996; Russo et al., 2004), optando tipicamente por abrigar-se em cavidades de árvores (Gebhard & Bogdanowicz, 2004; Strelkov, 2000). Por outro lado, na primavera e outono os morcegos podem economizar energia durante o dia entrando num estado de torpor, optando por abrigos que proporcionem temperaturas mais reduzidas (Grinevitch et al., 1995; Altringham, 1996).
Uma estrutura de fissão-fusão tem sido observada em colonias reprodutivas de N. leisleri (Kerth & König, 1999). Neste tipo de estrutura ocorrem variações diárias na composição estrutural do grupo presente em cada abrigo, sendo a coesão social de um grupo mais abrangente mantida. Alguns estudos sugerem que este tipo de comportamento tem como objetivo reforçar os laços sociais entre morcegos pertencentes à mesma colónia distribuída por uma vasta área florestal (O’Donnell, 2000; Willis & Brigham, 2004; O’Donnell & Sedgeley, 2006), podendo ainda aumentar a troca de informação relativamente à localização de abrigos (Kerth & Reckardt, 2003; Russo et al., 2005; O’Donnell & Sedgeley, 2006).
Esta espécie pode apresentar dois períodos de atividade durante uma noite, à semelhança de outras espécies do género Nyctalus (Russ, et al., 2003; Shiel, et al., 1998). Segundo Waters et al. (1999) o padrão exibido durante uma noite de caça é similar ao descrito por Catto et al. (1995) para Eptesticus serotinus, com um período inicial de emergência bem definido, logo após o pôr-do-sol, seguido de um segundo período de emergência menos bem definido, já no final da noite após os indivíduos terem regressado aos abrigos.
Devido à sua fisionomia, com asas relativamente longas e estreitas, N. leisleri consegue atingir velocidades elevadas, no entanto a sua capacidade de manobra em voo é mais reduzida, quando comparada com a de outras espécies de voo mais lento (Norberg & Rayner, 1987; Fleming, 2010). Por este motivo, caça geralmente em áreas abertas ou sobre a copa das árvores, em espaços livres de obstáculos (Figura 3) (Schober, 1997; Russ, 1999). Devido às características de voo, principalmente no que diz respeito à sua velocidade, é capaz de evitar a predação com maior facilidade que outras espécies, o que faz com que o seu primeiro período de emergência ocorra bastante cedo (Russ, 1999).
Figura 3 – Trajeto típico de voo em período de alimentação de N. leisleri (Fonte: Russ, 1999).
Diversos estudos revelaram que esta espécie apresenta uma grande diversidade de habitats de alimentação (Walsh & Harris, 1996; Vaughan et al., 1997, Waters et al., 1999; Shiel et al., 1999; Russ & Montgomery, 2003). Um estudo levado a cabo na Inglaterra por Waters et al. (1999) determinou que N. leisleri pode percorrer distâncias relativamente grandes entre os abrigos e as áreas de alimentação, utilizando como áreas preferenciais de caça zonas florestais, particularmente áreas entre limites florestais e espaços abertos, para além de pastagens e zonas ao longo de estruturas lineares, como é o caso de galerias ripícolas e estradas limitadas por vegetação arbustiva ou arbórea. O mesmo estudo indica que esta espécie evita áreas urbanas e zonas agrícolas enquanto se alimenta. Outro estudo, realizado na Irlanda por Shiel et al. (1999), indica que existe uma preferência por zonas de pastagem e canais de drenagem, que representam dois terços da atividade registada, mas também foi registada atividade sobre lagos e áreas florestais. Por sua vez, Vaughan et al. (1997), sugerem que esta espécie apresenta maiores indícios de atividade em áreas de pastagem e nas zonas envolventes de lagos. O estudo mais recente realizado por Szentkuti (2006), na Suíça, veio corroborar os registos de Walsh & Harris (1996), quanto à existência de uma preferência pelo uso de florestas folhosas relativamente a outro tipo de áreas florestais.
A sua dieta específica varia dependendo do habitat, ocorrendo no entanto uma predominância de insetos voadores de tamanho pequeno a médio. Tendo em conta a sua adaptação a voos rápidos e menor manobrabilidade, a sua estratégia de caça não passa por recolher insetos a partir de objetos imóveis, mas antes pela captura de presas em pleno voo (Russ, et al., 2003; Shiel, et al., 1998). Segundo um estudo de Shiel et al. (1998), a dieta de N. leisleri é principalmente composta por dípteros de várias famílias (Scathophagidae, Tipulidae, Anisopodidae e Chironomidae) de onde se destaca Scathophaga stercoraria, que perfaz 20 % da sua dieta. Segundo o mesmo estudo, outras possíveis fontes alimentares
são, coleópteros (Scarabaeidae), lepidópteros (Noctuidae), himenópteros (Ichneumonidae) e neurópteros (Chrysopidae e Hemerobiidae).
Relativamente à conservação, esta espécie apresenta atualmente uma ampla distribuição e abundancia, não existindo indícios de declínio significativo da sua população mundial. Por conseguinte, encontra-se classificada com estatuto de pouco preocupante (Hutson et al., 2008). No entanto no território continental Português apresenta um estatuto de informação insuficiente, visto não existirem informações adequadas para proceder à avaliação do seu risco de extinção, nomeadamente quanto ao tamanho da sua população e tendência de declínio (Cabral et al., 2006).
Esta espécie é particularmente suscetível à diminuição das florestas folhosas bem desenvolvidas, com a consequente redução das áreas de alimentação e disponibilidade de árvores com as características propícias para a constituição de abrigo. Outro fator que pode ameaçar esta espécie é o uso generalizado de pesticidas, devido à diminuição das presas disponíveis, bem como pela contaminação dos morcegos por ingestão de insetos expostos a este tipo de produto (Cabral et al., 2006). Por outro lado, tendo em conta a natureza migratória desta espécie, existe o risco de impacte provocado pela presença de parques eólicos nas suas rotas migratórias (Amorim, 2009; Rydell et al., 2010). Neste contexto, devido ao elevado número de registos de fatalidade em parques eólicos, torna-se premente a realização de estudos que permitam contrariar a escassez de dados relativos à sua real distribuição e tendência populacional em Portugal, de forma a objetivamente estabelecer medidas para a sua conservação (Cabral et al., 2006).
1.3. Breve Abordagem às Alterações Climáticas
A constituição da atmosfera terrestre tem sofrido transformações desde a sua formação, provocando uma dinâmica permanente no clima do planeta (IPCC, 2007), que está diretamente relacionada com a ocorrência de diversos episódios climáticos de arrefecimento e aquecimento, durante os cerca de 4600 milhões de anos de existência da Terra (Dodson et al., 2007). No entanto, desde o final do século XIX tem-se verificado um acentuar do aumento da temperatura atmosférica à superfície, com diferentes níveis de afetação a nível global (IPCC, 2007). As alterações climáticas provocadas por gases com efeito de estufa (GEE) de origem antropogénica, onde se inclui o dióxido de carbono, são aceites de uma forma geral, como a maior ameaça ambiental da atualidade e consequentemente, os esforços para manter a temperatura global em níveis sustentáveis tornaram-se numa das preocupações centrais dos decisores políticos (GWEC, 2008; Rodrigues et al., 2008).
Um dos maiores consumidores de energia é o sector das indústrias energéticas, principalmente a produção de eletricidade a partir da queima de combustíveis fósseis, tais como o petróleo, o carvão e o gás natural (IA, 2005), acabando por representar a maior fonte emissora, num total de cerca de 40 % das emissões de CO2 e aproximadamente 25 %
das emissões globais de GEE (EWEA, 2008). Por outro lado a depleção generalizada de combustíveis fósseis tem contribuído para a crescente preocupação relativamente à problemática das alterações climáticas (Hoel & Kverndokk, 1996). Perante este cenário, diversos países assinaram e ratificaram a Convenção Quadro das Nações Unidas sobre as Alterações Climáticas, em 1992, na Cimeira da Terra que teve lugar no Rio de Janeiro e mais recentemente o Protocolo de Quioto, em 1997 (Jones et al., 2009). Estes documentos constituem o enquadramento internacional para o combate às alterações climáticas, sendo que o Protocolo de Quioto determina o compromisso de redução das emissões de GEE por parte dos países desenvolvidos, estabelecendo limites juridicamente vinculativos (Jones et al., 2009).
No que diz respeito ao contexto Europeu, foi assumido um compromisso de desenvolvimento de métodos sustentáveis de produção de energia, de forma a combater as alterações climáticas e ao mesmo tempo dar resposta a um incremento das necessidades energéticas. Este compromisso foi declarado na Directiva 2001/77/EC do Parlamento Europeu e no Conselho de 27 de Setembro de 2001 sobre a promoção da produção de eletricidade utilizando fontes renováveis (Rodrigues et al., 2008). Desta forma, e atendendo às metas estabelecidas pelo Protocolo de Quioto, constatou-se um crescente desenvolvimento das tecnologias associadas à produção de energia através de fontes energéticas renováveis, com especial destaque para a energia eólica, que sofreu um aumento da potência instalada na União Europeia superior a qualquer outra tecnologia de produção elétrica nos últimos 10 anos, à exceção do gás natural (EWEA, 2011).
1.4. Energia Eólica – Situação Atual
A instalação anual de energia eólica tem vindo a aumentar de uma forma sustentada ao longo dos últimos 17 anos, de 814 MW em 1995 até 9 616 MW em 2011, resultando numa média anual de crescimento de 15,6%. Em 2011, encontravam-se instalados na União Europeia 93 954 MW, o que representa um incremento na capacidade cumulativa instalada de 11% relativamente ao valor registado no ano anterior (EWEA, 2012).
A Alemanha apresenta-se como o país da União Europeia com maior capacidade instalada, seguida pela Espanha, Itália, França e Reino Unido. Portugal encontra-se num grupo de 9 países, para além dos já referidos, com mais de 1 GW de capacidade instalada, juntamente com a Dinamarca, Holanda, Suécia, Irlanda, Grécia, Áustria, Polónia e Bélgica (EWEA, 2012).
Estima-se que com a capacidade instalada até ao final de 2011 seja possível a produção de 204 TWh, num ano de vento normal, o que representa 6,3% do consumo bruto anual da União Europeia. Neste contexto, a Dinamarca apresenta-se como o país de maior penetração de energia eólica no consumo de eletricidade, aproximadamente 26%, seguida da Espanha com 15,9% e de Portugal com 15,6%, enquanto Irlanda, Alemanha, Chipre e Grécia completam o grupo de países que superam a marca dos 5% (Figura 4) (EWEA, 2012).
Figura 4 – Penetração de energia eólica no consumo total de eletricidade dos países europeus até
Em Portugal, que se assume como um dos líderes Europeus em termos de penetração de energia eólica, durante o ano de 2011, a potência eólica ligada à rede pública aumentou 375 MW, totalizando no final do ano 4 081 MW que correspondem a uma potência instalada de 4 603 MVA (REN, 2011).
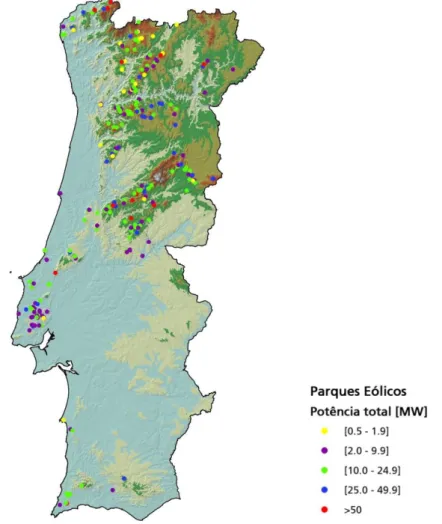
De acordo com a Directiva 2001/77/EC do Parlamento Europeu, Portugal deve aumentar a sua quota de energias renováveis no consumo final de energia, dos 20,5% registados em 2005, para 31% até 2020. De forma a cumprir os objetivos estipulados o governo português pretende aumentar a capacidade eólica para 6 875 MW até 2020 (GWEC, 2010). Neste sentido, em 2011 concluíram-se 13 novos parques eólicos, contabilizando-se no final do ano, 240 parques em funcionamento (Figura 5), com um total de 2349 aerogeradores (APREN, 2011).
Figura 5 – Localização e potência dos parques eólicos do território continental português (Fonte:
Apesar do forte investimento, nos últimos anos a Europa tem-se debatido com uma profunda crise económica. Confrontados com significativos défices orçamentais, vários países europeus visam conter o investimento nas energias renováveis a curto prazo. A troika composta pelo Fundo Monetário Internacional, o Banco Central Europeu e a Comissão Europeia têm supervisionado os orçamentos de Portugal, Grécia, Irlanda e Espanha, tendo mesmo aconselhado, por escrito, o governo Português a interromper o financiamento das energias renováveis através do seu orçamento. A dificuldade em obter financiamento tem provocado um significativo impacto no desenvolvimento de novos projetos de implementação de parques eólicos, sendo previsível um agravamento deste efeito e um abrandamento do desenvolvimento deste mercado (EWEA, 2012).
1.5. Vantagens e Desvantagens da Energia Eólica
Os benefícios ambientais do recurso a energias renováveis são claros, e neste prisma a energia eólica afigura-se como uma das fontes energéticas mais limpas, visto que não implica a libertação de GEE para atmosfera, para além de não produzir, nem utilizar materiais radioativos. De facto, calcula-se que com o recurso à energia eólica, em oposição ao uso de combustíveis fósseis, seja possível evitar a emissão para a atmosfera de 600 ton de CO2, por cada GWh produzido (GWEC, 2006).
Quando comparada com as fontes energéticas tradicionais, para além das vantagens ambientais, este tipo de energia não acarreta riscos geopolíticos ao reduzir a dependência energética externa, para além de não apresentar flutuações de custo, visto ser independente do preço dos combustíveis (TPWind Advisory Council, 2006). Tendo em conta que um parque eólico apresenta um período de construção menos demorado que outros projetos de energias renováveis, e que representa uma reduzida ocupação superficial de solo, este tipo de energia torna-se muito competitiva, relativamente às outras fontes energéticas renováveis. Existem ainda benefícios de cariz social devido aos postos de trabalho criados direta ou indiretamente para dar resposta ao desenvolvimento neste sector (EWEA, 2009).
Apesar das evidentes vantagens da energia eólica, algumas das limitações, bem como os impactes negativos da sua utilização devem ser considerados, sendo que a sua relevância depende em muitos casos da dimensão das turbinas e principalmente da localização da área em que é realizada a instalação do parque eólico (Barclay, 2007; Berkhuizen & Postma, 1991).
Uma das desvantagens no que diz respeito à localização dos parques deve-se ao facto de muitas vezes apenas se conseguirem reunir as condições necessárias para a sua implementação em locais remotos, onde a ligação à rede nem sempre é fácil, diminuindo
assim a competitividade relativamente com outras fontes energéticas. A disponibilidade de vento nem sempre coincide com o período de necessidade de abastecimento elétrico, o que aliado ao facto de existirem ainda diversas restrições ao nível do armazenamento acaba por comprometer a total satisfação das necessidades energéticas das populações (Denholm, 2006). Devido à intermitência deste recurso energético, verifica-se um fraco aproveitamento em regiões onde o vento é pouco constante, ou apresenta pouca intensidade (Regueiro, 2006).
Com o objetivo de rentabilizar a produção elétrica, a instalação dos parques eólicos deve ocorrer em áreas abertas com elevada velocidade de vento, requisitos que são cumpridos em zonas costeiras, zonas de alta montanha ou offshore (Drewitt & Langston, 2006). No caso particular de Portugal, verifica-se que os parques eólicos se distribuem por áreas de elevado interesse conservacionista, alguns dos quais de elevada sensibilidade ambiental (Travassos et al., 2005), o que levanta questões acerca do impacte visual, ruído e efeito sobre a fauna local (GWEC, 2006).
Alguma controvérsia tem surgido relativamente ao impacto visual dos aerogeradores nas paisagens, devido às suas consideráveis dimensões (GWEC, 2006). Em Portugal este aspeto é agravado pelo facto de os aerogeradores serem maioritariamente instalados em zonas de alta montanha (Travassos et al., 2005). O ruído provocado pelo movimento das hélices, embora seja relativamente reduzido, particularmente quando comparado com outras fontes de ruído industrial, pode também representar uma causa de impacte ambiental. No entanto, o desenvolvimento tecnológico e as melhorias ao nível do design e isolamento levaram à produção de turbinas eólicas modernas e com desempenhos muito superiores neste capítulo. Por outro lado, a abordagem das autoridades reguladoras tem sido garantir que as turbinas estão posicionadas suficientemente longe de zonas habitacionais, de modo a evitar perturbações relevantes (GWEC, 2006).
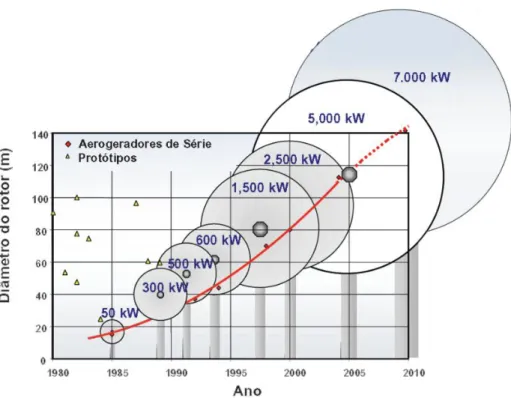
O incremento no número de parques eólicos e o progressivo aumento do tamanho dos aerogeradores (Figura 6), poderá representar um agravamento dos impactes causados sobre a fauna, para além de outros impactes gerais resultantes da instalação deste tipo de projeto (Barclay et al., 2007). Neste prisma, os efeitos negativos diretos sobre a avifauna e os quirópteros, nomeadamente a mortalidade provocada por colisão com aerogeradores, tem despertado o interesse da comunidade científica e do público em geral (Travassos et al., 2005).
Figura 6 – Evolução do tamanho dos aerogeradores comerciais (Fonte: http://www.dewi.de).
Para além dos efeitos diretos, existem impactes indiretos que devem ser tidos em conta, relativamente à avifauna, tais como a alteração do sucesso reprodutor; as alterações nos padrões migratórios; a perda de habitat de reprodução e alimentação, para além da alteração na disponibilidade de presas, por impactes diretos sobre a sua ecologia e comportamento (Orloff & Flannery, 1992). No caso dos quirópteros é de destacar, a perturbação ou destruição dos habitats de alimentação e dos corredores de comutação; a possível desorientação durante o voo devido à emissão de ultrassons e a perturbação ou destruição de áreas com potencial de abrigo (Rodrigues et al., 2008).
A generalidade dos estudos que aferem os impactes ecológicos na fauna, inerentes aos parques eólicos e estruturas associadas têm-se focado apenas na mortalidade de aves e morcegos, tendo-se vindo a prestar escassa atenção aos efeitos na biodiversidade em geral, particularmente no que diz respeito a outros grupos de animais (Hoover & Morrison, 2005; Rabin et al., 2006). No entanto, estudos mais recentes apontam para impactes no comportamento e riqueza específica de outros grupos de vertebrados terrestres (Santos et al., 2009).
Devido ao previsível incremento do número de parques eólicos instalados e considerando os possíveis impactes referidos, desde o ano 2000 os projetos de energia eólica encontram-se juridicamente sujeitos a um processo de Avaliação de Impacte Ambiental (AIA). O Decreto-Lei nº69/2000, de 3 de Maio, posteriormente alterado pelo Decreto-Lei nº197/2005, de 8 de Novembro, define que qualquer projeto com mais de 20
torres, ou localizado a uma distância inferior a 2 km de outro projeto semelhante, está sujeito a um processo de AIA, passando o número de torres a 10 nos casos em que os parques se localizem em áreas sensíveis. Neste contexto, são consideradas áreas sensíveis todas as áreas protegidas classificadas ao abrigo da legislação atual, os Sítios de Rede Natura 2000, Zonas Especiais de Conservação, Zonas de Proteção Especial e Áreas de proteção de monumentos nacionais e dos imóveis de interesse público.
1.6. Impacte dos Parques Eólicos nos Morcegos
A preocupação acerca do possível impacte dos parques eólicos, sobre a fauna prontamente surgiu associada às aves, devido à perturbação de espécies migradoras, que denotaram elevadas taxas de mortalidade por colisão com turbinas eólicas (Erickson et al, 2001). No entanto, foi apenas a partir do final dos anos 90 que começaram a surgir resultados de estudos que apontavam para os efeitos negativos destas estruturas em quirópteros, bem como para as dimensões deste impacte sobre as suas populações (e.g. Bach et al., 1999; Rhamel et al., 1999; Johnson et al., 2001; Kerns & Kerlinger, 2004). Desde então, diversos estudos têm vindo a confirmar a mortalidade de morcegos em parques eólicos (e.g. Erickson et al., 2002; Johnson, 2005; Arnett et al., 2008; Arnett et al., 2009). Tendo em conta o crescente número de projetos eólicos, bem como os indícios de declínio de muitas espécies, têm-se levantado preocupações sobre os efeitos a longo prazo, que podem ser induzidos pelos impactes cumulativos sobre as populações (Kunz et al., 2007b).
Anteriormente ao registo de mortalidade de morcegos em parques eólicos existiam escassos relatos de fatalidades por colisão com outras estruturas de origem antropogénica de grandes dimensões (Cryan & Barclay, 2009). Nas espécies de morcegos afetadas não existem evidências de outra forma de mortalidade induzida pelo Homem com magnitude similar à provocada pelas turbinas eólicas, ao contrário da situação verificada com as aves, relativamente ao seu fenómeno de colisão com estruturas artificiais de elevadas dimensões (Avery et al., 1980; Kuvlesky et al., 2007).
Nos últimos anos têm-se realizado progressos consideráveis ao nível: da estimação de taxas de mortalidade; do estudo comportamental de morcegos nas imediações de turbinas eólicas; na descrição das condições climatéricas de noites em que ocorreu mortalidade e na discussão dos potenciais impactes dos parques eólicos nas populações de morcegos em declínio. No entanto, apesar de numerosas hipóteses terem sido formuladas, não são ainda reconhecidos, de uma forma inequívoca, os fatores que levam a que os morcegos sejam atraídos para estas estruturas, aumentando assim a probabilidade de
mortalidade (Cryan & Barclay, 2009). Por outro lado, subsistem dúvidas relativamente às causas de morte que, para além da colisão direta, se podem dever a um fenómeno designado por barotrauma. Este fenómeno ocorre devido a uma rápida redução na pressão do ar perto das turbinas em movimento, provocando danos nos tecidos e consequentes lesões internas, principalmente em órgãos como os pulmões, devido à expansão do ar no seu interior em níveis incomportáveis para a expiração (Kunz et al., 2007a; Dürr & Bach, 2004).
Diversas espécies de morcegos utilizam estruturas lineares durante os períodos de alimentação e nas migrações. A instalação de parques eólicos em áreas florestais pode levar à formação deste tipo de estruturas (lineares), aumentando consequentemente a utilização destes espaços pelos morcegos (Kunz et al., 2007a). Alguns estudos (Ahlén, 2003; Horn et al., 2008) indicam que os parques eólicos podem também representar pontos de referência na paisagem, funcionando assim como focos de atração. Algumas teorias têm sido desenvolvidas neste sentido, explicando este fenómeno de atração devido ao som produzido pelas pás das turbinas em funcionamento ou devido ao movimento e consequente turbulência do ar por elas criada (Ahlén, 2003; Kunz et al., 2007a; Horn et al., 2008). Outros estudos (Rydell et al., 2010) indicam que a mortalidade em parques eólicos se pode dever à elevada velocidade na extremidade das pás, que dificulta ou impossibilita a sua deteção por parte dos morcegos. Apesar de não ser totalmente conhecida a forma como os morcegos se orientam durante as migrações, julga-se que a emissão de sons é menos frequente de forma a evitar gastos energéticos durante os voos em maior altitude, aumentando assim a probabilidade de colisão com obstáculos inesperados (Ahlén, 2003; Kunz et al., 2007a). Os morcegos podem ainda ser atraídos pelas turbinas, na tentativa de encontrarem zonas de descanso ou abrigo, ao confundirem as torres dos aerogeradores com árvores altas, principalmente durante as migrações (Ahlén, 2003; Kunz et al., 2007a; Horn et al., 2008). Outro fator prende-se com a concentração de insetos junto aos aerogeradores que pode ser induzida pela radiação térmica emitida pela turbina, ou ainda por fenómenos conhecidos de aglomeração em zonas mais altas da paisagem por parte de diversos insetos voadores (Ahlén, 2003). Os morcegos respondem a este tipo de aglomeração aumentando assim o risco de mortalidade durante o período de alimentação (Ahlén, 2003; Kunz et al., 2007a).
Dados relativos a 2011 apontavam para a ocorrência de mortalidade em mais de 13 países Europeus, num total de 3545 fatalidades registadas até abril desse ano, 466 das quais ocorridas em Portugal (EUROBATS, 2011). Existe registo de mortalidade para 11 das 25 espécies de morcegos que ocorrem no território continental português, destacando-se devido à elevada mortalidade as espécies Pipistrellus pipistrellus, Nyctalus leisleri, Hypsugo savii, Pipistrellus pygmaeus e Pipistrellus kuhli (ICNB, 2010).
Neste contexto, torna-se fundamental obter dados que ajudem a compreender as verdadeiras razões que levam à mortalidade, bem como desenvolver ferramentas de carácter preditivo, que permitam proceder à determinação dos possíveis riscos associados à instalação de novos projetos eólicos, de forma a delinear de uma forma mais objetiva, estratégias para a minimização destes impactes sobre as espécies afetadas (Barnard, 2000).
1.7. Modelação Baseada em Agentes
Os Modelos Baseados em Agentes (Agent-based models - ABMs), também denominados Modelos Baseados em Indivíduos, são ferramentas de simulação computacional capazes de incorporar inteligência, combinando elementos de aprendizagem, adaptação, evolução e lógica difusa. Este tipo de modelos depende de uma lógica de abordagem invertida, que começa por considerar os componentes de um sistema (i.e. os agentes individuais), tentando posteriormente compreender a forma como as propriedades do sistema emergem a partir das interações entre os seus diversos componentes (Grimm, 1999; Grimm & Railsback, 2005). Desta forma, a característica primordial de um ABM é a utilização do indivíduo como unidade básica, deixando de se representar a população por uma variável contínua, mas antes pelo conjunto das entidades discretas pelas quais é composta, os indivíduos (Jorgensen, 1994).
Os agentes contemplados neste tipo de modelação agem de forma autónoma, sem qualquer tipo de controlo externo; apresentam-se direcionados para o cumprimento de objetivos específicos; podem mover-se pelo ambiente revelando consciência e reatividade relativamente às mudanças ambientais; podem ainda ser concebidos para adaptar o seu estado e comportamento em resposta a estímulos por parte de outros agentes ou do ambiente. Esta ênfase na interação entre os agentes e o ambiente é o que distingue os ABMs de outras abordagens de modelação (Marceau, 2008).
A modelação baseada em agentes permite a compreensão efetiva de um conjunto complexo de comportamentos e interações, sendo por isso altamente adequado para simular fenómenos complexos (Grimm et al., 2005). Ao longo dos últimos 15 anos, os ABMs têm sido aplicados a uma vasta gama de questões relacionadas com a gestão de recursos ambientais (Bousquet & Le Page, 2004), embora a sua utilização ocasional se tenha iniciado na década de setenta (Kaiser, 1979). Este tipo de modelo tem também sido amplamente utilizado em ecologia, no estudo de dinâmicas populacionais, devido à sua flexibilidade que permite o uso detalhado de parâmetros com maior significado biológico, sendo portanto
mais realistas do que modelos populacionais clássicos, mais presos dentro de um rígido formalismo matemático (DeAngelis & Mooij, 2005).
Mais recentemente tem-se recorrido aos ABMs de uma forma interdisciplinar para abordar questões relativas às interações entre o Homem e a vida selvagem (An et al., 2005; Anwar et al., 2007). A recente proliferação dos ABMs em estudos de ecologia, nomeadamente no campo da mobilidade e comportamento animal (Wang & Grimm, 2007; Stillman, 2008), sugere que a sua aplicação poderá ter um papel fulcral na compreensão do processo de seleção de habitats e avaliação da necessidade de implementação de planos conservacionistas.
Tendo em conta, a singularidade das espécies selvagens, que constituem entidades adaptativas e sensíveis ao ambiente, o recurso aos ABMs, como ferramenta de gestão ambiental, poderá ser vantajoso visto que acomoda facilmente uma interação dinâmica entre agentes, para além de potenciar uma aproximação realista às condições ambientais e possibilitar a simulação de um conjunto de cenários hipotéticos (McLane et al., 2011).
2. Objetivos
Tendo em conta o elevado número de ameaças a que os morcegos se encontram sujeitos, para além da enorme falta de conhecimento relativo a diversas espécies, é expectável que as populações de morcegos possam estar em risco. Este estudo teve como principal objetivo o desenvolvimento de uma ferramenta preditiva de carácter conservacionista, com enfoque no possível impacte de parques eólicos sobre o morcego arborícola Nyctalus leisleri. Em concreto simulou-se a atividade de N. leisleri, numa “paisagem real” de forma a criar mapas que evidenciem as áreas de maior potencial de uso do espaço, por parte desta espécie. Criaram-se ainda cenários com o intuito de compreender a forma como algumas variáveis influenciam a mortalidade em parques eólicos, e como esta poderá depender de fatores relacionados com a ecologia da própria espécie.
3. Material e Métodos
3.1. Área de Estudo
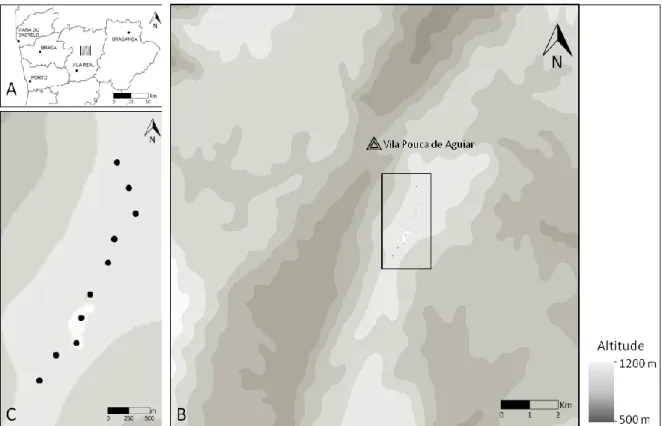
O presente estudo incidiu numa área de 22 500 ha, centrada no parque eólico do Negrelo e Guilhado, situado nas freguesias de Soutelo de Aguiar e Vila Pouca de Aguiar, do Concelho de Vila Pouca de Aguiar, distrito de Vila Real (Figura 7) (EDP, 2006). A área abrangida pelo parque eólico localiza-se na serra da Padrela, uma região montanhosa, com predominância granítica, a nascente da serra do Alvão, sendo que o projeto se encontra integrado, quase na sua totalidade, no Sítio de Importância Comunitária (SIC) PTCO0003 - Alvão-Marão. A maior parte da área abrangida pelo parque está sujeita a Regime Florestal, encontrando-se a restante classificada como Espaços Agrícolas (LEA, 2011).
Figura 7 – Localização da área de estudo em Portugal Continental (A). Localização do parque eólico do Negrelo e Guilhado, dentro da área de estudo (B). Pormenor do parque eólico do Negrelo e Guilhado (C) Os círculos indicam a localização dos aerogeradores.
O parque eólico é constituído por 10 aerogeradores modelo E-82, da fabricante de turbinas eólicas ENERCON. Cada aerogerador, com potência unitária de 2 000 KW, apresenta uma altura de torre de 80m e um diâmetro de rotação das pás de 82m. A área de implantação do parque atinge altitudes superiores a 1000 m com cota máxima no marco geodésico de Negrelo (1112m), embora os aerogeradores se encontrem
instalados, mais precisamente, ao longo da linha de cumeada da serra da Padrela, entre os 1000 m e os 1100 m (EDP, 2006).
A nível edáfico, verificam-se limitações significativas à exploração agrícola ou pastagem melhorada. Estas limitações devem-se essencialmente à presença abundante de afloramentos rochosos, aos riscos acentuados de erosão e a fatores climáticos, visto que o regime de temperaturas origina a formação de geadas durante quase todo o ano (EDP, 2006).
A rede hidrográfica é formada por diversas linhas de água, num raio inferior ou igual a 10 km do parque eólico, das quais se destaca a ribeira da Peliteira, que se encontra a cerca de 2,4 km de distância da área abrangida pelo parque (Moreira, 2006).
Na área de implantação do parque eólico verifica-se uma modificação considerável da vegetação, de origem antropogénica, sobretudo pelo uso do fogo que propicia uma paisagem dominada por formações arbustivas e herbáceas de baixa complexidade e reduzida biomassa, onde dominam diversas espécies de leguminosas, nomeadamente tojos, carquejas e giestas. As zonas mais húmidas encontram-se ocupadas predominantemente por vidoal (LEA, 2011).
Relativamente à fauna, é de destacar o registo de presença de Tartaranhão-caçador (Circus pygargus) e de Lobo (Canis lupus signatus) (EDP, 2006; Ecosfera, 2009), ambos com estatuto de conservação de “Em Perigo” (Cabral et. al., 2006). O grupo dos Quirópteros encontra-se representado por 8 espécies (Nyctalus leisleri, Rhinolophus ferrumequinum, Pipistrellus kuhli, Pipistrellus pipistrellus, Hypsugo savii, Eptesicus serotinus, Barbastella barbastellus, Tadarida teniotis) e ainda a possibilidade de utilização da área do parque por indivíduos de 3 grupos de espécies (Myotis myotis / Myotis blythii, Plecotus auritus / Plecotus austriacus e Pipistrellus pipistrellus / Miniopterus schreibersii / Pipistrellus pygmaeus) (Moreira, 2006; Ecosfera 2007; Ecosfera 2008; LEA, 2009).