Reversal of corticosterone-induced BDNF alterations by the natural
antioxidant alpha-lipoic acid alone and combined with desvenlafaxine:
Emphasis on the neurotrophic hypothesis of depression
Caren Nádia Soares de Sousa
a, Lucas Nascimento Meneses
a, Germana Silva Vasconcelos
a,
Márcia Calheiros Chaves Silva
a, Jéssica Calheiros da Silva
b, Danielle Macêdo
a,
David Freitas de Lucena
a, Silvânia Maria Mendes Vasconcelos
a,naNeuropsychopharmacology Laboratory, Department of Physiology and Pharmacology, Federal University of Ceará, Fortaleza, Ceará, Brazil bFederal University of Ceará, Fortaleza, Ceará, Brazil
a r t i c l e
i n f o
Article history:
Received 20 January 2015 Received in revised form 11 August 2015 Accepted 31 August 2015 Available online 1 September 2015
Keywords:
Alpha-lipoic acid Desvenlafaxine Major depressive disorder
Treatment resistant depressive disorder Corticosterone model of depression BDNF
a b s t r a c t
Brain derived neurotrophic factor (BDNF) is linked to the pathophysiology of depression. We hypothe-sized that BDNF is one of the neurobiological pathways related to the augmentation effect of alpha-lipoic acid (ALA) when associated with antidepressants. Female mice were administered vehicle or CORT 20 mg/kg during 14 days. From the 15th to 21st days the animals were divided in groups that were further administered: vehicle, desvenlafaxine (DVS) 10 or 20 mg/kg, ALA 100 or 200 mg/kg or the combinations of DVS10þALA100, DVS20þALA100, DVS10þALA200 or DVS20þALA200. ALA or DVS alone or in combination reversed CORT-induced increase in immobility time in the forced swimming test and decrease in sucrose preference, presenting, thus, an antidepressant-like effect. DVS10 alone reversed CORT-induced decrease in BDNF in the prefrontal cortex (PFC), hippocampus (HC) and striatum (ST). The same was observed in the HC and ST of ALA200 treated animals. The combination of DVS and ALA200 reversed CORT-induced alterations in BDNF and even, in some cases, increased the levels of this neu-rotrophin when compared to vehicle-treated animals in HC and ST. Taken together, these results suggest that the combination of the DVSþALA may be valuable for treating conditions in which BDNF levels are
decreased, such as depression.
&2015 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Major depressive disorder (MDD) can become a chronic or lifelong condition, with many patients experiencing recurrent depressive episodes. In this regard, for a significant number of patients, the disorder has an obstinately nature, withfluctuations over time in the severity of depressive symptoms (Kessler, 2003, 2012). This disorder has been considered the leading cause of disease-related disability especially among women. The
prevalence of this mental disorder among women has typically been between one and a half to three times that of men (Kessler, 2003). Depressed mood, diminished interest/pleasure and fatigue or loss of energy are among the core symptoms of MDD (Fried and Nesse, 2015).
The pathophysiological mechanisms related to MDD include alterations in: (i) neurotransmitter metabolism, (ii) neuroendo-crine function, (iii) neurotrophins, (iv) inflammatory pathways; (iv) glutamate release and (v) oxidative imbalance (Miller et al., 2009). The neurotrophic alterations observed in depression are mainly associated with decreased levels of brain-derived neuro-trophic factor (BDNF). Indeed, this neurotrophin is regarded as an important biomarker to various psychiatric disorders such as de-pression (Schmidt et al., 2011; Melo et al., 2013), schizophrenia (Nieto et al., 2013;Vasconcelos et al., 2015;Macêdo et al., 2012a, 2012b), Alzheimer′s disease (Rosa and Fahnestock, 2014) and Parkinson's disease (Howells et al., 2000).
Still at present the treatment of MDD has important limitations. In one of the largest clinical trials on depression, the“Sequenced Treatment Alternatives to Relieve Depression” (STAR*D), only Contents lists available atScienceDirect
journal homepage:www.elsevier.com/locate/psychres
Psychiatry Research
http://dx.doi.org/10.1016/j.psychres.2015.08.042
0165-1781/&2015 Elsevier Ireland Ltd. All rights reserved.
n
Correspondence to: Department of Physiology and Pharmacology, Federal University of Ceará, Cel. Nunes de Melo street, 1127, 60431-270 Fortaleza, CE, Brazil. Fax:þ55 85 3366 8333.
E-mail addresses:[email protected](C.N.S. Sousa),
[email protected](L.N. Meneses),
[email protected](G.S. Vasconcelos),
[email protected](M.C.C. Silva),
[email protected](J.C. Silva),
[email protected](D. Macêdo),
[email protected](D.F. de Lucena),[email protected],
about one fourth of the patients enrolled achieved remission during thefirst treatment stage (Trivedi et al., 2007). In this case, one of the sequenced treatment alternatives for not remitting patients are the serotonin and norepinephrine reuptake inhibitors (SNRIs), such as desvenlafaxine-DVS (Fried and Nesse, 2015). Un-fortunately, the management of treatment-resistant depression (TRD) continues to challenge mental health care professionals and researchers (Kasper, 2014).
Among the available preclinical models of depression, the model based on the repeated administration of corticosterone (CORT) (Zhao et al., 2008) was proposed to model TRD (Ago et al., 2013) and psychotic major depression (Iijima et al., 2010). This evidence was based on preclinical studies in which the selective serotonin reuptake inhibitor (SSRI)fluvoxamine and the tricyclic antidepressant imipramine were absent of effects against the in-creased immobility time induced by chronic CORT administration, while the combination of risperidone and fluvoxamine was ef-fective (Iijima et al., 2010).
Using the CORT-induced model of depression we recently showed that the administration of DVS (10 or 20 mg/kg) and of the antioxidant alpha-lipoic acid (ALA–100 or 200 mg/kg) alone or in combination reversed the increase in the immobility time in the forced swimming and tail suspension tests (Silva et al., 2013). Accordingly, in this previous study we observed in the forced swimming test that the administration of ALA 200 mg/kg and DVS 10 mg/kg or 20 mg/kg to female mice brought the greater anti-depressant-like effects (Silva et al., 2013).
In the last decades, oxidative and nitrosative mechanisms have been associated with MDD (Moylan et al., 2014). In this regard, ALA (1,2-dithiolane-3-pentanoic acid), also known as thioctic acid is a natural antioxidant synthesized in the human body (Ferreira et al., 2009) and its antidepressant effect wasfirst hypothesized in the last decade (Salazar, 2000). Currently, a preclinical study from our research group added further evidences to this hypothesis as aforementioned (Silva et al., 2013). Besides ALA's antioxidant ef-fect, one possible explanation for the augmentation of DVS anti-depressant-like effects by ALA (Silva et al., 2013) seemingly relies on the interference of this antioxidant with dopaminergic neuro-transmission (Deslauriers et al., 2013). This speculation is strengthened by the clinical evidences of greater efficacy for treatment of depression with drugs that interfere with the three monoamines, such as monoamine oxidase inhibitors (Chen and Skolnick, 2007).
Thus, given the current importance of the neurotrophic hy-pothesis of depression and also in the high incidence of MDD in women, we hypothesized that the antidepressant-like effects of ALA alone or combined with DVS, as previously demonstrated by our research group (Silva et al., 2013), could be related to altera-tions in BDNF levels in putative brain areas related to depression in female mice. In order to confirm this hypothesis, herein we aimed to determine the levels of BDNF in the prefrontal cortex (PFC), hippocampus (HC) and striatum (ST) of female mice submitted to the model of depression induced by the chronic administration of CORT and treated with ALA or DVS alone or in combination.
2. Methods
2.1. Animals
Female Swissmice (30–32 g) were obtained from the Animal House of the Federal University of Ceara, Brazil. The animals were kept in a room with a controlled temperature of 2371°C, under a standard light-dark cycle withad libitumaccess to food and water, except during the experiments. Food was removed 4 h prior to the oral gavage procedure and returned 20 min after. The animals
were divided into one of the followingfive experimental groups (n¼7–10 animals/group) according described in each respective.
All experiments were performed according to the National In-stitutes of Health Guide for the Care and Use of Laboratory Ani-mals (NIH, 1996) and all efforts were made to minimize the suf-fering of the animals and to reduce the number of animals used in the experiments. The project was approved by the Animal Ethics Committee of the Faculty of Medicine of the Federal University of Ceará, under protocol number 88/2013.
2.2. Drugs
Corticosterone (CORT, Sigma-Aldrich, St Louis, MO, USA) was dissolved in a saline solution containing 0.1% dimethyl sulfoxide and 0.3% Tween-80. Corticosterone 20 mg/kg was administered as a single daily subcutaneous injection, from 09:00 to 11:30 a.m. for twenty-one consecutive days. The dosage and route of adminis-tration for CORT was selected based previous studies (Zhao et al., 2008;Silva et al., 2013). Alpha-lipoic acid (ALA, Sigma-Aldrich, St. Louis, MO, USA) was dissolved in distilled water and 0.2% car-boxymethyl cellulose and orally administered (p.o.) for 7 con-secutive days at doses of 100 or 200 mg/kg according to previous studies (Silva et al., 2013). Desvenlafaxine succinate monohydrate (DVS, Pristiqs, Wyeth Lab) was dissolved in distilled water and
administered orally (p.o.) for 7 consecutive days at the doses of 10 or 20 mg/kg. DVS was administered alone or 1 h before the ad-ministration of ALA. The dosages for DVS were calculated from human doses on an mg/m2basis (Reagan-Shaw et al., 2008).
2.3. Treatment groups
The animals were randomly divided into fourteen experimental groups as shown inFig. 1. In the present study, different animals were used for BDNF assay and for behavioral determinations.
Control group: Mice in this group received daily injections of saline solution containing 0.1% dimethyl sulfoxide and 0.3% Tween-80 (s.c.).
Groups treated with DVS or ALA alone: Mice in these groups received daily injections of saline solution containing 0.1% di-methyl sulfoxide and 0.3% Tween-80 (s.c.) for 14 consecutive days followed by DVS (10 or 20 mg/kg, p.o.) or ALA (100 or 200 mg/kg, p.o.) for further 7 days.
CORT-induced depression model: Mice in this group received daily subcutaneous injections of CORT (20 mg/kg, s.c.) once a day, between 09:00 and 11:30 a.m., for 21 days (Zhao et al., 2008).
Corticosterone-induced depression model and treatment with DVS or ALA alone: Mice in these groups received repeated injections of CORT during 14 days to induce depressive-like behavior. From the 15th to 21st days of administration, the animals were randomly divided into the four following groups: CORT and DVS (10 or 20 mg/kg) or CORT and ALA (100 or 200 mg/kg).
Corticosterone-induced depression model and treatment with the combination ofDVSþALA: From the 15th to 21st days of CORT
administration, the mice were randomly divided into groups that were further treated with DVS (10 or 20 mg/kg). One hour after DVS administration the animals were given ALA (100 or 200 mg/ kg). The time interval between the administration of DVS and ALA was set at 1 h because a previous study showed that the maximum mean plasma concentration of DVS in mice occurred 1 h after oral administration of the drug (DeMaio et al., 2011). The experimental design is represented inFig. 1.
2.4. Behavioral assessment
2.4.1. Forced swimming test
treatment, 1 h after administration. Mice were individually forced to swim in an open cylindrical container (diameter, 22 cm; height, 40 cm) that contained 20 cm of water held at 2571°C. The total time during which the mouse remained immobile during a 5 min period was recorded. Immobility was defined as the animal
floating in the water without struggling and making only very minimal movements necessary to keep its head above the water. An increase in the duration of immobility is an indicative of de-pressive-like behavior (Porsolt et al., 1977).
2.4.2. Sucrose preference test
Sucrose preference test was carried out on the twentieth day of treatment, 1 h after administration. The test was performed as described previously (Mao et al., 2014). Briefly, 72 h before the test, mice were trained to adapt 1% sucrose solution (w/v): two bottles of 1% sucrose solution were placed in each cage, and 24 h later 1% sucrose in one bottle was replaced with tap water for 24 h. After the adaptation, mice were deprived of water and food for 24 h. Sucrose preference test was conducted at 9:00 a.m. in which mice were housed in individual cages and were free to access two bottles containing 100 ml of sucrose solution (1% w/v) and 100 ml of water. After 1 h, the volumes of consumed sucrose solution and water were recorded and the sucrose preference was calculated by the following formula:
Sucrose preference=water consumptionSucrose consumptionSucrose consumption ×100% +
2.5. Dissection of brain areas
Twenty-four hour after the last drug administration, the ani-mals were sacrificed by rapid decapitation and the brains were quickly removed and placed on aluminum foil in a Petri dish on ice. Prefrontal cortex (PFC), hippocampus (HC) and striatum (ST) were dissected for the measurement of BDNF.
To remove the PFC, anterior portion of the frontal lobes (about 1.5 mm from the olfactory bulb) was removed and a bilateral section taken with the aid of a microdissection scissors (Machado, 2000).
After the withdrawal of PFC, following the median sagittalfi s-sure, cerebral cortical layer of the meninges was removed with the aid of a straight clamp microdissection, which, progressing deli-cate and tangentially to the lateral ventricles, separated the cortex in all its fronto-occipital extension. The cortex separated has been
folded to the side, exposing the hippocampal region (HC) and part of the striatum (ST). The HC and ST were isolated from the sur-rounding structures with a microdissection scissors, oriented with its withdrawal by the diameter of the tuberous visible portion of these nuclei after folding the lateral cortex.
After the dissection, CPF, HC and ST were placed in microtubes properly identified, weighed and stored at 70°C until use for measurement of BDNF.
2.6. Determination of BDNF levels
The brains areas, PFC, HC and ST were homogenized in phos-phate buffered saline (pH 7.4) with a protease inhibitor cocktail (Sigma St. Louis, USA). BDNF levels in each sample were quantified by immunoenzymatic determination according to the manu-facturer′s instructions (EMD Millipore, USA). The results are ex-pressed as BDNF/pg tissue.
2.7. Statistical analysis
Statistical analysis was performed with GraphPad Prism 6.0 for Windows, GraphPad Software (San Diego, CA, USA). The results were evaluated by one-way ANOVA followed by Bonferroni post hoc test. All results are expressed as means7S.E.M (standard er-rors of the mean). Before ANOVA, D′Agostino-Pearson omnibus test was conducted to verify the normal distribution of the data. For all analyses, the significance level was set at
α
¼0.05.3. Results
3.1. Effects of CORT, ALA and DVS on depressive-like behavior of mice in the forced swimming test
The forced swimming test was performed to assess depressive-like behavior in all experimental groups after 21 days of admin-istration of CORT or saline. ANOVA revealed a significant difference between groups [F(13, 88)¼38.94;po0.0001] (Fig. 2). Bonferroni post hoc test showed an increase in the immobility time in ani-mals submitted to the chronic administration of CORT when compared to control animals (po0.0001). Mice treated with DVS (10 or 20) or ALA (100 or 200) and saline presented no significant
alterations when compared to control animals. On the other hand, the administration of DVS (10 or 20) (po0.0001), ALA100 (po0.0001) or ALA200 (po0.01) alone or in combination (po0.0001) after CORT reversed the depressive-like alteration induced by CORT. Remarkably, the mice post-treated with the drugs in combination, i.e. DVS10þALA100, DVS10þALA200 after
CORT showed significant lower immobility time when compared to the respective groups that received DVS10 (po0.0001), ALA100 or ALA200 aloneþCORT. The same potentiation of
antidepressant-like effect was observed in the groups administered DVS20þALA100 and DVS20þALA200 after CORT when compared
to DVS20, ALA100 or ALA200 aloneþCORT (po0.0001) (Fig. 2).
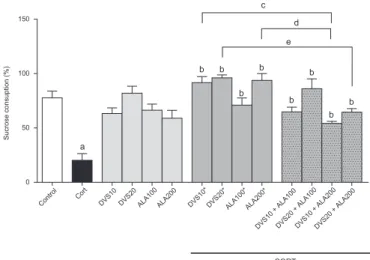
3.2. Effects of CORT, ALA and DVS on anhedonic-like behavior of mice in the sucrose preference test
ANOVA showed a significant difference between groups [F(13, 106)¼13.32;po0.0001] (Fig. 3). Bonferroni post hoc test revealed that CORT-treated animals presented a significant decrease in su-crose preference when compared to control animals (po0.0001), what is related to an anhedonic-like behavior. Desvenlafaxine or ALA when administered with saline caused no significant altera-tion. On the other hand, the post-administration of DVS (10 or 20) or ALA (100 or 200) after CORT reversed the anhedonic-like al-teration induced by CORT (po0.0001). Similarly, the administra-tion of DVS and ALA in combinaadministra-tion reversed the decrease in su-crose consumption observed in mice treated with CORT (DVS10þALA100: po0.001; DVS20þALA100: po0.0001; DVS10þALA200:po0.01; DVS20þALA200:po0.001). Curiously, the groups post-treated with the combinations of DVSþALA200
after CORT presented a decrease in sucrose consumption when compared with the groups post-administered the drugs alone and CORT (DVS10:po0.001; DVS20:po0.05; ALA200:po0.001).
3.3. Effects of CORT, ALA and DVS in brain BDNF levels
In the PFC of mice administered CORT during 21 days (Figure 4A) a stastisticaly significant difference decrease in BDNF levels was observed when compared to control animals (po0.0001) [F(13, 112)¼17.83,po0.0001]). The administration of DVS10 (po0.0001), DVS10þALA200 (po0.0001) or DVS20þALA200 (po0.0001), when compared to CORT-treated
mice, significantly reversed the decrease in BDNF levels induced by CORT. Furthermore, the addition of ALA200 to DVS20 presents a beneficial effect more robust when compared to groups treated with CORT and DVS20 (po0.0001) or ALA200 (po0.01). On the other hand, the administration of DVS20, ALA100, ALA200 or the combinations DVS10þALA100, DVS20þALA100 to CORT-treated
mice did not reverse the decrease in BDNF, showing levels of BDNF similar to CORT-treated. The administration of the drugs alone, i.e. without CORT, caused no changes in BDNF levels when compared with control group.
In the HC (Fig. 4B) the administration of CORT alone during 21 days significantly decreased the levels of BDNF when compared to control (po0.05) [F(13, 116)¼5.720,po0.01]. The administration of ALA200 (po0.05), as well as the combination of DVS10þALA200 (po0.001) or DVS20þALA200 (po0.0001), sig-nificantly reversed the decrease in BDNF induced by CORT. The highest levels of BDNF in the HC were observed in the animals administered DVS20þALA200 since the results obtained in this
group were significant when compared to control (po0.05), CORT (po0.0001) or CORTþDVS20 (po0.0001) groups. The stand-alone treatments caused no changes in HC BDNF levels when compared with control group.
In the ST (Fig. 4C), as observed in the other brain areas used in this study, reduced levels of BDNF were observed after CORT ad-ministration when compared to control animals (po0.0001) [F(13, 112)¼20.46, po0.0001]. The administration of ALA200 (po0.0001) or the combination of DVS10þALA200 (po0.0001) or DVS20þALA200 (po0.0001) reversed the decrease in BDNF levels induced by CORT when compared to CORT-treated mice. On the contrary, the administration of DVS20, ALA100 or the combi-nation DVS20þALA100 did not reverse the decrease in BDNF,
showing levels of this neurotrophin similar to CORT-treated. Higher levels of BDNF were observed in the groups treated with ALA200, DVS10þALA200 or DVS20þALA200. The administration
of the drugs alone, i.e. without CORT, caused no changes in ST BDNF levels when compared with control group.
4. Discussion
The present study is thefirst to show that treatment with ALA, alone or in combination with DVS, is able to reverse the
Fig. 2.Effects of DVS or ALA treatment alone or in combination on the immobility time on the FST in a chronic CORT-induced depression mice model. Each bar re-presents the mean7SEM ofn¼5–8 animals/group.apo0.05 vs. control,bpo0.05
vs. CORT,cpo0.05 vs. CORTþDVS10,dpo0.05 vs. CORTþDVS20,epo0.05 vs.
CORTþALA100,fpo0.05 vs. CORTþALA200, according to one-way ANOVA
fol-lowed by Bonferroni post hoc test. Abbreviations: CORT–corticosterone; DVS–
desvenlafaxine; ALA–alpha-lipoic acid; FST–forced swimming test.
Control Cort
DVS10 DVS20 ALA100 ALA200 DVS10* DVS20* ALA100*ALA200*
DVS10 + ALA100DVS20 + ALA100DVS10 + ALA200DVS20 + ALA200 0
50 100 150
Sucrose consuption (%)
CORT
a
b b
b b
b b
b b c
d
e
Fig. 3.Effects of DVS or ALA treatment alone or in combination on the sucrose preference test in a chronic CORT-induced depression mice model. Each bar re-presents the mean7SEM ofn¼6–10 animals/group.apo0.05 vs. control,bpo0.05
vs. CORT,cpo0.05 vs. CORTþDVS10, dpo0.05 vs. CORTþDVS20,epo0.05 vs.
anhedonic-like behavior and the changes in BDNF concentrations caused by chronic administration of CORT. Besides that, ALA pre-sent antidepressant-like effects demonstrated in FST, mainly when administered in association with DVS, confirming previously published data (Silva et al., 2013).
According to ourfindings, the chronic administration of CORT cause an increase in immobility time in the FST and reduce sucrose consumption in SPT. In fact, the model of depression induced by
chronic administration of CORT induces behavioral alterations that resemble depression (Zhao et al., 2008). Studies show that animals submitted to this model present a depressive-like phenotype as observed by the increases in immobility time in the FST and tail suspension test (TST), anxiety-like phenotype as observed by de-crease in the open arms entries in the plus maze test and anhe-donic-like phenotype represented by reduce in sucrose con-sumption in SPT (Zhao et al., 2008;Huang et al., 2011;Silva et al., 2013), corroborating with data of current study.
Moreover, our results also showed that DVS exhibited anti-depressant activity in the CORT-induced model of depression de-monstrated by FST and SPT. Desvenlafaxine is the major pharma-cologically active metabolite of venlafaxine and exhibit a low risk to pharmacokinetics interactions due display a little interface with the P-450 isoenzyme system, besides not subject to 2D6 influences (i.e., genetic polymorphism) (Sansone and Sansone, 2014).
A recent study showed that DVS prevents the onset of de-pressive-like and anxiety-like phenotypes in animals exposed to unpredictable chronic mild stress (Wang et al., 2014), corroborat-ing with the data observed in our study. Based on the hypothesis that white matter dysfunction is an etiological factor of depres-sion, the same study showed that DVS improved the reduce phosphorylation of AMPK-a and prevented oligodendrocyte im-pairment (Wang et al., 2014). Furthermore, in a previous study of our research group, DVS decreased the immobility time in FST and TST in animal model of depression-induced by CORT, confirming the results of current investigation (Silva et al., 2013).
An investigation about depression post-myocardial infarction also showed that DVS reduces apoptosis in limbic structures such as amygdala, brain area closely related to symptoms as anhedonia and behavioral despair observed in depression (Malick et al., 2014). These data corroborate our findings, which show a sig-nificant improvement in anhedonic-like behavior (SPT) and be-havioral despair (FST) in the groups treated with DVS.
Besides the effects demonstrated by DVS, ALA also showed antidepressant-like effects observed in FST and SPT. Indeed, recent reports suggested a possible antidepressant-like effect from the acute and repeated administration of ALA (Silva et al., 2013,2014). Moreover, another study demonstrated that this antioxidant is able to restore reduced glutathione (GSH) levels, the major brain antioxidant, decrease lipid peroxidation and restore decreased BDNF levels in animal models of mania and schizophrenia (Ma-cêdo et al., 2012a,2012b; Vasconcelos et al., 2015). All of these parameters have been shown to be altered in depressed patients (Chung et al., 2012;Carballedo et al., 2013;Dalby et al., 2013) and in animals in the CORT-induced model of depression (Huang et al., 2011), justifying the antidepressant-like effects observed in FST and SPT.
In our experiments we also observed that the combination of ALA and DVS demonstrated a more robust antidepressant-like effect in FST when compared to the actions of each of the drugs alone in CORT-induced depression model. This effect can be ex-plained by the potentiation of the antioxidant effect of DVS and by association of different mechanisms of action (i.e., reuptake of monoamines and antioxidant). In fact, a recent literature review has asserted that antidepressants also present antioxidant prop-erties in clinical trials on MDD (Behr et al., 2012).
The potentiation effect was not observed in the sucrose pre-ference test and, unlike the expected, the antidepressant-like ef-fect was reduced when associating DVS and ALA compared to the drug alone. Thisfinding may be related to a possible pharmaco-kinetic or pharmacodynamics interaction, causing a reduction in the concentration of drug required for anti-anhedonia effect.
In addition to behavioral changes, the animal model of de-pression induced by corticosterone also was able to cause reduc-tion in levels of BDNF in the prefrontal cortex, hippocampus and
Fig. 4.Alterations in BDNF levels by the administration of ALA or DVS alone or in combination in the prefrontal cortex (A), hippocampus (B) and striatum (C) of mice chronically administered CORT. Each bar represents the mean7SEM ofn¼7–10 animals/group. Infigure A, apo0.05 vs control,bpo0.05 vs CORT,cpo0.05 vs
CORTþDVS10,dpo0.05 vs CORTþDVS20,epo0.05 vs CORTþALA200. Infigure B, apo0.05 vs control,bpo0.05 vs CORT,cpo0.05 vs CORTþDVS20. In figure C, apo0.05 vs control,bpo0.05 vs CORT,cpo0.05 vs CORTþDVS10,dpo0.05 vs
CORTþDVS20, according to one-way ANOVA followed by Bonferroni post hoc test.
striatum of mice, mimicking the BDNF deficit characteristic of the pathophysiology of depression and corroborating data already present in the literature (Yulug et al., 2009;Bumb et al., 2014;O′ Donovan et al., 2014;Guo et al., 2014). Several studies have shown that animal models of depression have, besides the depressive-like behavior, a BDNF deficit in brain areas (Ye et al., 2011;Gibney et al., 2013;Guo et al., 2014;Yi et al., 2014).
The proposed model produces changes in emotional behavior that may correspond to clinical symptoms of depression in hu-mans, as observed in results of current study (Zhao et al., 2008; Silva et al., 2013;O′Donovan et al., 2014). This model relies on the theory that depression is related to a dysregulation of the hy-pothalamic-pituitary-adrenal (HPA) axis and to elevated corticos-terone levels resulting from chronic stress (Ogłodek et al., 2014; Zhu et al., 2014;Jacobson, 2014).
According to the hypothesis of neurotrophins in depression, stress reduces the activity of BDNF, which results in decreased function of the limbic regions of the brain involved in cognition and emotional processing (Duman and Monteggia, 2006;Groves, 2007). There are several pre-clinical and clinical evidence that stress and trauma cause reduction in levels of BDNF and that an-tidepressant treatment reverts this situation (Nibuya et al., 1999; Roceri et al., 2002; Kauer-Sant′Anna et al., 2007;Grassi-Oliveira et al., 2008;Sen et al., 2008;Munno et al., 2013;Molendijk et al., 2014; Martocchia et al., 2014; Mikoteit et al., 2014). However, antidepressant pharmacotherapy is associated with high rates of treatment resistance and this fact is a major challenge in the treatment of depressive disorders. Studies suggest that this re-sistance to therapy is associated with a genetic polymorphism in BDNF gene (Egan et al., 2014;Cattaneo et al., 2013;El-hage et al., 2015) and the stress is one of the factors cited as causes of this polymorphism (Hosang et al., 2014).
Thus, studies have implicated BDNF as a predictive marker re-lated to the efficacy or treatment failure, and can be used as a marker of treatment success (Hu et al., 2010;Tadic et al., 2011; Wolkowitz et al., 2011).
Our data shows that in the CORT groups treated with ALA (100 or 200) or DVS (10 or 20), only DVS at the lowest dose reversed the CORT-induced alterations in BDNF levels in the PFC, seeming to be a dose-dependent effect. In fact, 50 mg/day has been established as the clinically effective dose of this drug for the management of MDD (Liebowitz and Tourian, 2010). When we calculated the translated dose from humans to mice (Reagan-Shaw et al. 2008), the 10 mg/kg dose in mice was the closest to the 50 mg/day dose that is used in the management of MDD in humans, which could explain why we observed the best results with DVS at the 10 mg/ kg dose in our study. Moreover, similar effect was observed in TST of recent study in the same experimental protocol (Silva et al., 2013).
Furthermore, ALA100 did not reverse the effects of CORT on BDNF levels in prefrontal cortex and striatum. Indeed, the action of antioxidants is often variable depending on the redox state of the cellular environment (Flora, 2009), which justifies also the fact that DVS10 is effective alone although it is ineffective in combi-nation with ALA100 in prefrontal cortex. In addition, in behavioral tests of present study, treatment with ALA100 exhibited a lower anti-anhedonia-like effect as compared to DVS or ALA200,
con-firming the effect found on BDNF.
On the other hand, the combination of ALA200 to both doses of DVS was capable to reverse CORT effect on BDNF levels. This data suggests that the combination of ALA200 to DVS, mainly DVS20, is more effective in reversing the low content of BDNF in the three brain areas investigated. Thesefindings corroborate with the be-havioral results presented here and with previous studies from our group that showed the reversal effect of ALA on the depressive-like behavior of animals treated with CORT (Silva et al., 2013,
2014). Furthermore, several studies show that antioxidants are able to reverse depressive-like behavior alone or in association with antidepressants (Lobato et al., 2010; Moretti et al., 2012; Mazloom et al., 2013;Manosso et al., 2013;Amr et al., 2013).
The prefrontal cortex, one of the most studied brain areas in the biology of depression, is associated with important functions such as problem solving, analysis, control of impulses and motivations, selective attention, emotion control and working memory (Garcia et al., 2009; Li et al., 2010; Cai et al., 2013). Studies show that depression evolves with a reduction in the size of the prefrontal cortex and the bloodflow to that area (Porto et al., 2002;Singh and Gotlib, 2014). Possibly, these changes are associated with re-duction of BDNF, since this factor is responsible for neurogenesis and neuroplasticity of the central nervous system (Kimpton, 2012). In addition, playing an important role in the progression of de-pression, BDNF deficiency is implicated as a cause of the dys-function observed in depressive disorders that are associated with the prefrontal cortex, for example, loss of concentration and at-tention, compulsion, cognitive deficits and suicidal ideation (Miller and Cohen, 2001;Yuen et al., 2012;Dombrovski et al., 2013). Thus, as the combination of ALA and DVS shown to be effective in re-versing the low concentrations of BDNF, treatment with both drugs shows promise in reducing symptoms associated with pre-frontal cortex.
Similar to the results obtained in the prefrontal cortex, our study also showed that ALA200, as well as the combination of ALA200 to DVS at both doses, was able to reverse low concentra-tions of BDNF in the hippocampus of mice treated with CORT. In fact, evidence suggests that, besides of antidepressant and anti-oxidant effects when combined with DVS (Silva et al., 2013,2014), ALA is able to elevate the concentrations of BDNF in the hippo-campus of animals (Macêdo et al., 2012a,2012b; Miao et al., 2013), confirming thefindings of the present study.
The hippocampus is a brain area that is closely associated with symptoms such as apathy, loss of interest, sleep disturbances, changes in weight/appetite and memory deficits, seen in many depressed patients (Jiang et al., 2013;Bai et al., 2014;Jayaweera et al., 2015). Expression of BDNF in hippocampus particularly, is related to clinical manifestation of certain psychiatric and cogni-tive disorders such as depression (Yi et al., 2014;Mitchelmore and Gede, 2014). Studies show that observed reductions in the levels of BDNF are associated with decreased hippocampal volume, deficits in episodic memory and dysfunction in activities of daily life (Czéh and Lucassen , 2007;Frodl et al., 2007). Additionally, evidence has shown that the hippocampus is strictly linked to the regulation of the hypothalamus-pituitary-adrenal (HPA) axis by to present dense expression of glucocorticoid receptors responsible for con-trol of HPA negative feedback (Sterner and Kalynchuk, 2010). The importance of this axis to the pathophysiology of depression is based on clinical evidence from patients with Cushing′s syndrome who have high rates of depression, as well as depressed patients present hypercortisolemia and disrupted cortisol rhythmicity (Starkman et al., 2001;Sonino and Fava, 2002).
Given the importance of the integrity of the hippocampus in regulation of the HPA axis and hippocampal function, the asso-ciated treatment with ALA and DVS, mainly ALA200 and DVS20, configures itself as an innovator in reducing symptoms and pro-gression of depressive disorders therapy.
Finally, our study shows that ALA200 alone also was able to reverse the CORT effect on concentrations of BDNF in the striatum of mice. Furthermore, the combination of ALA200 to DVS, at both doses, reversed the low concentrations of BDNF observed in CORT treated group. These data corroborate studies that show the an-tidepressant and antioxidant effects from combination of ALA and DVS (Silva et al., 2013,2014).
striatum in the neurobiology of depression, this area is funda-mentally important for understanding the mechanisms involved in the pathophysiology of this disorder.
This area is related to reward, cognition and movement, being primarily responsible for the symptoms of lack of motivation, fa-tigue and psychomotor alterations observed in depressed patients (Harrington, 2012;Walther et al., 2012). So, it is a region of in-terface between emotion, cognition and motor control. Dysfunc-tion and death of striatal neurons are the major causes of motor disorders and BDNF is among the factors that promote the survival and proper function of this neuronal population (Baydyuk and Xu, 2014). Clinical and preclinical studies show that Parkinson′s Dis-ease, a neurodegenerative disorder characterized by the death of dopaminergic neurons in the striatum, is associated with the de-velopment of depression, indicating the involvement of striatal neurons and the relation of this area in the pathophysiology of depression (Lemke, 2008;Vriend et al., 2014). Furthermore, there is evidence that shows that the rise of BDNF in the striatum is associated with reduction of depressive-like behavior (Marais et al., 2009).
Therefore, the combination of ALA and DVS, by reversing the low concentrations of BDNF caused by treatment with CORT, shows a promising therapy for the treatment of depression and reversal of motor disorders resulting from this disorder.
It is worth mentioning that increase in BDNF levels is not only important for the management of depression. A number of dis-eases are related to alterations in the levels of this neurotrophin. In this regard, patients with neurodegenerative and neuropsychia-trics diseases seems to benefit from strategies that increases BDNF (Zuccato and Cattaneo, 2009; Molendijk et al., 2014; Fernandes et al., 2014).
Moreover, the data obtained in this investigation provide sci-entific subsidy for the antioxidants to be used in treatment of depressive disorders and supports studies that suggest the anti-depressant effect of ALA.
Additionally our results are seemingly promising since the augmentation therapy with the natural antioxidant ALA may possibly reduce DVS dose and consequently the side effects of this therapy.
Conflict of interest
The authors declare no conflict of interest.
Acknowledgments
This study was supported by Grants from the Brazilian Gov-ernment Institutions National Council for Scientific and Techno-logical Development (CNPq), Coordenação de Aperfeiçoamentode Pessoal de Nível Superior (CAPES) and Fundação Cearense de Apoio ao DesenvolvimentoCientífico e Tecnológico, (FUNCAP).
References
Ago, Y., Yano, K., Araki, R., Hiramatsu, N., Kita, Y., Kawasaki, T., Onoe, H., Chaki, S., Nakazato, A., Hashimoto, H., Baba, A., Takuma, K., Matsuda, T., 2013. Metabo-tropic glutamate 2/3 receptor antagonists improve behavioral and prefrontal dopaminergic alterations in the chronic corticosterone-induced depression model in mice. Neuropharmacology 65, 29–38.
Amr, M., El-Mogy, A., Shams, T., Vieira, K., Lakhan, S.E., 2013. Efficacy of vitamin C as an adjunct tofluoxetine therapy in pediatric major depressive disorder: a randomized, double-blind, placebo-controlled pilot study. Nutr. J. 12 (31), 1–8.
Baia, M., Zhua, X., Zhanga, Y., Zhanga, S., Zhanga, L., Xuea, L., Zhonga, M., Zhang, X., 2014. Anhedonia was associated with the dysregulation of hippocampal HTR4 and microRNA Let-7a in rats. Physiol. Behav. 129, 135–141.
Baydyuk, M., Xu, B., 2014. BDNF signaling and survival of striatal neurons. Front Cell Neurosci. 8, 1–10.
Behr, G.A., Moreira, J.C.F., Frey, B.N., 2012. Preclinical and clinical evidence of an-tioxidant effects of antidepressant agents: implications for the pathophysiology of major depressive disorder. Oxid. Med. Cell. Longev. 2012, 13.
Bumb, J.M., Aksay, S.S., Janke, C., Kranaster, L., Geisel, O., Hellweg, R., Sartorius, A., 2015. Focus on ECT seizure quality: serum BDNF as a peripheral biomarker in depressed patients. Eur. Arch. Psychiatry Clin. Neurosci. 265, 227–232.
Cai, X., Kallarackal, A.J., Kvarta, M.D., Goluskin, S., Gaylor, K., Bailey, A.M., Lee, H.K., Huganir, R.L., Thompson, S.M., 2013. Local potentiation of excitatory synapses by serotonin and its alteration in rodent models of depression. Nat. Neurosci. 16, 464–472.
Carballedo, A., Morris, D., Zill, P., Fahey, C., Reinhold, E., Meisenzahl, E., Bondy, B., Gill, M., Moller, H.J., Frodl., T., 2013. Brain-derived neurotrophic factor Val66Met polymorphism and early life adversity affect hippocampal volume. Am. J. Med. Genet. B: Neuropsychiatr. Genet. 22, 32130.
Cattaneo, A., Gennarelli, M., Uher, R., Breen, G., Farmer, A., Aitchison, K.J., Craig, I.W., Anacker, C., Zunsztain, P.A., McGuffin, P., Pariante, C.M., 2013. Candidate genes expression profile associated with antidepressants response in the GENDEP study: differentiating between baseline‘predictors′and longitudinal‘targets’. Neuropsychopharmacology 38, 377–385.
Chen, Z., Skolnick, P., 2007. Triple uptake inhibitors: therapeutic potential in de-pression and beyond. Expert Opin. Invest. Drugs 16, 1365–1377.
Chung, C.P., Schmidt, D., Stein, C.M., Morrow, J.D., Salomon, R.M., 2012. Increased oxidative stress in patients with depression and its relationship to treatment. Psychiatry Res. 11, 665–668.
Czéh, B., Lucassen, P.J., 2007. What causes the hippocampal volume decrease in depression? Are neurogenesis, glial changes and apoptosis implicated?. Eur. Arch. Psy. Clin. Neurosci. 257 (5), 250–260.
Dalby, R.B., Elfving, B., Poulsen, P.H., Foldager, L., Frandsen, J., Videbech, P., Rosen-berg, R., 2013. Plasma brain-derived neurotrophic factor and prefrontal white matter integrity in late-onset depression and normal aging. Acta Psychiatry Scand. 27, 12085.
DeMaio, W., Kane, C.P., Nichols, A.I., Jordan, R., 2011. Metabolism studies of des-venlafaxine. J. Bioequiv. Availab. 03, 151–160.
Deslauriers, J., Desmarais, C., Sarret, P., Grignon, S., 2013.α-Lipoic acid interaction with dopamine D2 receptor-dependent activation of the Akt/GSK-3βsignaling pathway induced by antipsychotics: potential relevance for the treatment of schizophrenia. J. Mol. Neurosci.: MN 50, 134–145.
Dombrovski, A.Y., Szanto, K., Clark, L., Reynolds III, C.F., Siegle, G.J., 2013. Reward signals, attempted suicide, and impulsivity in late-life depression. JAMA Psy-chiatry 70 (10), 1020–1030.
Duman, R.S., Monteggia, L.M., 2006. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 59, 1116–1127.
Egan, M.F., Kojima, M., Callicott, J.H., Goldberg, T.E., Kolachana, B.S., Bertolino, A., Zaitsev, E., Gold, B., Goldman, D., Dean, M., Lu, B., Weinberger, D.R., 2014. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 112, 257–269.
El-hage, W., Vourc’h, P., Gaillard, P., Léger, J., Belzung, C., Ibarguen-Vargas, D., An-dres, C.R., Camus, V., 2015. The BDNF Val66Met polymorphism is associated with escitalopram response in depressed patients. Psychopharmacology 232 (3), 575–581.
Fernandes, B.S., Berk, M., Turck, C.W., Steiner, J., Gonçalves, C.A., 2014. Decreased peripheral brain-derived neurotrophic factor levels are a biomarker of disease activity in major psychiatric disorders: a comparative meta-analysis. Mol. Psychiatry 19 (7), 750–751.
Ferreira, P.M.P., Militão, G.C.G., Freitas, R.M., 2009. Lipoic acid effects on lipid per-oxidation level, superoxide dismutase activity and monoamines concentration in rat hippocampus. Neurosci. Lett. 464, 131–134.
Flora, S.J., 2009. Structural, chemical and biological aspects of antioxidants for strategies against metal and metalloid exposure. Oxid. Med. Cell. Longev. 2 (4), 191–206.
Fried, E.I., Nesse, R.M., 2015. Depression is not a consistent syndrome: an in-vestigation of unique symptom patterns in the STAR*D study. J. Affect. Disord. 172, 96–102.
Frodl, T., Schüle, C., Schmitt, G., Born, C., Baghai, T., Zill, P., Bottlender, R., Rupprecht, R., Bondy, B., Reiser, M., Möller, H., Meisenzahl, E.M., 2007. Association of the brain-derived neurotrophic factor Val66Met polymorphism with reduced hip-pocampal volumes in major depression. Arch. Gen. Psychiatry 64 (4), 410–416.
Garcia, I.S., Comim, C.M., Valvassori, S.S., Reus, G.Z., Stertz, L., Kapczinski, F., Gavioli, E.C., Quevedo, J., 2009. Ketamine treatment reverses behavioral and physiolo-gical alterations induced by chronic mild stress in rats. Prog. Neuropsycho-pharmacol. Biol. Psychiatry 33, 450–455.
Gibney, S.M., Mcguinness, B., Prendergast, C., Harkin, A., Connor, T.J., 2013. Poly I:C-induced activation of the immune response is accompanied by depression and anxiety-like behaviours, kynurenine pathway activation and reduced BDNF expression. Brain Behav. Immun. 28, 170–181.
Grassi-Oliveira, R., Stein, L.M., Lopes, R.P., Teixeira, A.L., Bauer, M.E., 2008. Low plasma brain-derived neurotrophic factor and childhood physical neglect are associated with verbal memory impairment in major depression–a preliminary report. Biol. Psychiatry 64, 281–285.
Groves, J.O., 2007. Is it time to reassess the BDNF hypothesis of depression? Mol. Psychiatry 12, 1079–1088.
behavior in mice. Neuroscience 263, 1–14.
Harrington, M.E., 2012. Neurobiological studies of fatigue. Prog. Neurobiol. 99 (2), 93–105.
Hosang, G.M., Shiles, C., Tansey, K.E., Mcguffin, P., Uher, R., 2014. Interaction be-tween stress and the BDNF Val66Met polymorphism in depression: a sys-tematic review and meta-analysis. BMC Med. 12, 7.
Howells, D.W., Porritt, M.J., Wong, J.Y., Batchelor, P.E., Kalnins, R., et al., 2000. Re-duced BDNF mRNA expression in the Parkinson’s disease substantia nigra. Exp. Neurol. 166, 127–135.
Huang, Z., Zhonga, X., Li, Z., Fenga, C., Pana, A., Mao, Q., 2011. Curcumin reverses corticosterone-induced depressive-like behavior and decrease in brain BDNF levels in rats. Neurosci. Lett. 493, 145–148.
Hu, Y., Yu, X., Yang, F., Si, T., Wang, W., Tan, Y., Zhou, D., Wang, H., Chen, D., 2010. The level of serum brain-derived neurothophic factor is associated with the ther-apeutic efficacy of modified electroconvulsive therapy in Chinese patients with depression. J. ECT 26 (2), 121–125.
Iijima, M., Ito, A., Kurosu, S., Chaki, S., 2010. Pharmacological characterization of repeated corticosterone injection-induced depression model in rats. Brain Res. 1359, 75–80.
Jacobson, L., 2014. Hypothalamic-pituitary-adrenocortical axis: neuropsychiatric aspects. Compr. Physiol. 4 (2), 715–738.
Jayaweera, H.K., Hickie, I.B., Duffy, S.L., Hermens, D.F., Mowszowski, L., Diamond, K., Terpening, Z., Paradise, M., Lewis, S.J., Lagopoulos, J., Naismith, S.L., 2015. Mild cognitive impairment subtypes in older people with depressive symptoms: relationship with clinical variables and hippocampal change. J. Geriatr. Psy-chiatry Neurol. 28, 174–183, pii: 0891988715573535.
Jiang, W., Li, S., Liu, J., Sun, Y., Zhou, S., Zhu, W., Shi, J., Lu, L., 2013. Hippocampal CLOCK protein participates in the persistence of depressive-like behavior in-duced by chronic unpredictable stress. Psychopharmacology 227 (1), 79–92.
Kasper, S., 2014. Treatment-resistant depression: a challenge for future research. Acta Neuropsychiatr. 26, 131–133.
Kauer-Sant’Anna, M., Tramontina, J., Andreazza, A.C., Cereser, K., da Costa, S., Santin, A., Yatham, L.N., Kapczinski, F., 2007. Traumatic life events in bipolar disorder: impact on BDNF levels and psychopathology. Bipolar Disord. 9, 128–135.
Kessler, R.C., 2012. The costs of depression. Psychiatr. Clin. North Am. 35, 1–14.
Kessler, R., 2003. Epidemiology of women and depression. J. Affect. Disord. 74, 5–13.
Kimpton, J., 2012. The brain derived neurotrophic factor and influences of stress in depression. Psychiatr. Danub. 24, 169–171.
Lemke, M.R., 2008. Depressive symptoms in Parkinson's disease. Eur. J. Neurol. 15 (1), 21–25.
Li, N., Lee, B., Liu, R.J., Banasr, M., Dwyer, J.M., Iwata, M., Li, X.Y., Aghajanian, G., Duman, R.S., 2010. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science 329, 959–964.
Liebowitz, M.R., Tourian, K.A., 2010. Efficacy, safety, and tolerability of desvenla-faxine 50 mg/d for the treatment of major depressive disorder: a systematic review of clinical trials. Prim. Care Companion J. Clin. Psychiatry 12 (3), e1–e10.
Lobato, K.R., Cardoso, C.C., Binfaré, R.W., Budnia, J., Wagner, C.L.R., Brocardo, P.S., de Souza, L.F., Brocardo, C., Flesch, S., Freitas, A.E., Dafré, A.L., Rodrigues, A.L.S., 2010.α-Tocopherol administration produces an antidepressant-like effect in predictive animal models of depression. Behav. Br. Res. 209, 249–259.
Macêdo, D.S., Araújo, D.P., Sampaio, L.R.L., Vasconcelos, S.M.M., Sales, P.M.G., Sousa, F.C.F., Hallak, J.E., Crippa, J.A., Carvalho, A.F., 2012a. Animal models of prenatal immune challenge and their contribution to the study of schizophrenia: a systematic review. Braz. J. Med. Biol. Res. 45, 179–186.
Macêdo, D.S., Medeiros, C.D., Cordeiro, R.C., Sousa, F.C., Santos, J.V., Morais, T., Hy-phantis, T.N., McIntyre, R.S., Quevedo, J., Carvalho, A.F., 2012b. Effects of alpha-lipoic acid in an animal model of mania induced byD-amphetamine. Bipolar Disord. 14, 707–718.
Machado, A.B.M., 2000. Functional Neuroanatomy. Atheneu, São Paulo.
Malick, M., Gilberta, K., Barrya, M., Godbouta, R., Rousseaua, G., 2014. Desvenla-faxine reduces apoptosis in amygdala after myocardial infarction. Br. Res. Bull. 109, 158–163.
Manosso, L.M., Neis, V.B., Moretti, M., Daufenbach, J.F., Freitas, A.E., Colla, A.R., Ro-drigues, A.L.S., 2013. Antidepressant-like effect ofα-tocopherol in a mouse model of depressive-like behavior induced by TNF-α. Prog. Neuropsycho-pharmacol. Biol. Psychiatry 46, 48–57.
Mao, Q., Huang, Z., Zhong, X., Xiana, Y., Ipa, S., 2014. Brain-derived neurotrophic factor signalling mediates the antidepressant-like effect of piperine in chroni-cally stressed mice. Behav. Br. Res. 261, 140–145.
Martocchia, A., Curto, M., Scaccianoce, S., Comite, F., Xenos, D., Nasca, C., Falaschi, G. M., Ferracuti, S., Girardi, P., Nicoletti, F., Falaschi, P., 2014. Effects of escitalopram on serum BDNF levels in elderly patients with depression: a preliminary report. Aging Clin. Exp. Res. 26 (4), 461–464.
Marais, L., Stein, D.J., Daniels, W.M.U., 2009. Exercise increases BDNF levels in the striatum and decreases depressive-like behavior in chronically stressed rats. Metabol. Br. Disease 24, 587–597.
Mazloom, Z., Ekramzadeh, M., Hejazi, N., 2013. Efficacy of supplementary vitamins C and E on anxiety, depression and stress in type 2 diabetic patients: a ran-domized, single-blind, placebo-controlled trial. Pak. J. Biol. Sci. 16 (22), 1597–1600.
Mello, B.S., Monte, A.S., McIntyre, R.S., Soczynska, J.K., Custódio, C.S., Cordeiro, R.C., Chaves, J.H., Vasconcelos, S.M., Nobre Jr., H.V., Florenço de Sousa, F.C., Hy-phantis, T.N., Carvalho, A.F., Macêdo, D.S., 2013. Effects of doxycycline on de-pressive-like behavior in mice after lipopolysaccharide (LPS) administration. J. Psychiatr. Res. 47 (10), 1521–1529.
Miao, Y., Ren, J., Jiang, L., Liu, J., Jiang, B., Zhang, X., 2013.α-lipoic acid attenuates obesity-associated hippocampal neuroinflammation and increases the levels of brain-derived neurotrophic factor in ovariectomized rats fed a high-fat diet. Int. J. Mol. Med. 32 (5), 1179–1186.
Mitchelmore, C., Gede, L., 2014. Brain derived neurotrophic factor: Epigenetic reg-ulation in psychiatric disorders. Br. Res. 1586, 162–172.
Mikoteit, T., Beck, J., Eckert, A., Hemmeter, U., Brand, S., Bischof, R., Holsboer-Trachsler, E., Delini-Stula, A., 2014. High baseline BDNF serum levels and early psychopathological improvement are predictive of treatment outcome in major depression. Psychopharmacology 231 (15), 2955–2965.
Miller, E.K., Cohen, J.D., 2001. An integrative theory of prefrontal cortex function. Ann. Rev. Neurosci. 24, 167–202.
Miller, A.H., Maletic, V., Raison, C.L., 2009. Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol. Psychiatry 65, 732–741.
Molendijk, M.L., Spinhoven, P., Polak, M., Bus, B.A.A., Penninx, B.W.J.H., Elzinga, B.M., 2014. Serum BDNF concentrations as peripheral manifestations of depression: evidence from a systematic review and meta-analyses on 179 associations (N¼9484). Mol. Psychiatry 19 (7), 791–800.
Moretti, M., Colla, A., Balen, G.O., Santos, D.B., Budni, J., Freitas, A.,E., Farina, M., Rodrigues, A.L.S., 2012. Ascorbic acid treatment, similarly tofluoxetine, reverses depressive-like behavior and brain oxidative damage induced by chronic un-predictable stress. J. Psychiatry Res. 46, 331–340.
Moylan, S., Berk, M., Dean, O.M., Samuni, Y., Williams, L.J., O’Neil, A., Hayley, A.C., Pasco, J.A., Anderson, G., Jacka, F.N., Maes, M., 2014. Oxidative & nitrosative stress in depression: why so much stress? Neurosci. Biobehav. Rev. 45, 46–62.
Munno, D., Sterpone, S., Fania, S., Cappellin, F., Mengozzi, G., Saroldi, M., Bechon, E., Zullo, G., 2013. Plasma brain derived neurotrophic factor levels and neu-ropsychological aspects of depressed patients treated with paroxetine. Pan-minerva Med. 55 (4), 377–384.
Nibuya, M., Takahashi, M., Russell, D.S., Duman, R.S., 1999. Repeated stress increases catalytic TrkB mRNA in rat hippocampus. Neurosci. Lett. 267, 81–84.
Nieto, R., Kukuljan, M., Silva, H., 2013. BDNF and Schizophrenia: from neurodeve-lopment to neuronal plasticity learning, and memory. Front. Psychiatry 4, 45. NIH, 1996. Guide for the Care and Use of Laboratory Animals, Institute of Laboratory
Animal Research, National Research, National Academies Press Council.
O’Donovan, S., Dalton, V., Harkin, A., McLoughlin, D.M., 2014. Effects of brief pulse and ultrabrief pulse electroconvulsive stimulation on rodent brain and beha-viour in the corticosterone model of depression. Int. J. Neuropsychopharmacol. 17, 1477–1486.
Ogłodek, E., Szota, A., Just, M., Mos, D., Araszkiewicz, A., 2014. The role of the neuroendocrine and immune systems in the pathogenesis of depression. Pharmacol. Rep. 66, 776–781.
Porsolt, R.D., Bertin, A., Jalfre, M., 1977. Behavioral despair in mice: a primary screening test for antidepressants. Arch. Int. Pharmacodyn. Ther. 229, 327–336.
Porto, P., Hermolin, M., Ventura, P., 2002. Neuropsychological impairment asso-ciated with depression. Rev. Bras. Ter. Comport. Cogn. 4 (10), 63–70.
Reagan-Shaw, S., Nihal, M., Ahmad, N., 2008. Dose translation from animal to hu-man studies revisited. FASEB J.: Off. Publ. Fed. Am. Soc. Exp. Biol. 22, 659–661.
Roceri, M., Hendriks, W., Racagni, G., Ellenbroek, B.A., Riva, M.A., 2002. Early ma-ternal deprivation reduces the expression of BDNF and NMDA receptor subunits in rat hippocampus. Mol. Psychiatry 7, 609–616.
Rosa, E., Fahnestock, M., 2014. Amyloid-Beta, BDNF, and the Mechanism of neuro-degeneration in Alzheimer’s disease. In: Handbook of Neurotoxicity, pp. 1597–
1620.
Salazar, M.R., 2000. Alpha lipoic acid: a novel treatment for depression. Med. Hy-potheses 55, 510–512.
Sansone, R.A., Sansone, L.A., 2014. Serotonin norepinephrine reuptake inhibitors: a pharmacological comparison. Innov. Clin. Neurosci. 11 (3–4), 37–42.
Schmidt, H.D., Shelton, R.C., Duman, R.S., 2011. Functional biomarkers of depres-sion: diagnosis, treatment, and pathophysiology. Neuropsychopharmacol.: Off. Publ. Am. College Neuropsychopharmacol. 36, 2375–2394.
Sen, S., Duman, R., Sanacora, G., 2008. Serum brain-derived neurotrophic factor, depression, and antidepressant medications: meta-analyses and implications. Biol. Psychiatry 64, 527–532.
Silva, M.C., Sampaio, L.R., de Araújo, D.P., Araújo, P.V., Monte, A.S., Rodrigues, F.T., Woods, D.J., de Sousa, F.C., Fonteles, M.M., Vasconcelos, S.M., 2014. Central ef-fects of lipoic acid associated with paroxetine in mice. Am. J. Ther. 21 (2), 85–90.
Silva, M.C.C., de Sousa, C.N.S., Sampaio, L.R.L., Ximenes, N.C., Araújo, P.V.P., da Silva, J.C., de Oliveira, S.L., Sousa, F.C.F., Macêdo, D.S., Vasconcelos, S.M.M., 2013. Augmentation therapy with alpha-lipoic acid and desvenlafaxine: a future target for treatment of depression? Naunyn-Schmiedebergs Arch. Pharmacol. 386, 685–695.
Singh, M.K., Gotlib, I.H., 2014. The neuroscience of depression: implications for assessment and intervention. Behav. Res. Ther. 2014, 1–14.
Sonino, N., Fava, G.A., 2002. Residual symptoms in depression an emerging ther-apeutic concept. Prog. Neuropsychopharmacol. Biol. Psychiatry 26, 763–770.
Starkman, M.N., Giordani, B., Berent, S., Schork, M.A., Schteingart, D.E., 2001. Ele-vated cortisol levels in cushings disease are associated with cognitive decre-ments. Psychosom. Med. 63, 985–993.
Sterner, E.Y., Kalynchuk, L.E., 2010. Behavioral and neurobiological consequences of prolonged glucocorticoid exposure in rats: relevance to depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 34, 777–790.
Neuropsychopharmacol. Biol. Psychiatry 35, 415–420.
Trivedi, M.H., Rush, A.J., Gaynes, B.N., Stewart, J.W., Wisniewski, S.R., Warden, D., Ritz, L., Luther, J.F., Stegman, D., Deveaugh-Geiss, J., Howland, R., 2007. Max-imizing the adequacy of medication treatment in controlled trials and clinical practice: STAR(*)D measurement-based care. Neuropsychopharmacol.: Off. Publ. Am. Coll. Neuropsychopharmacol. 32, 2479–2489.
Vasconcelos, G.S., Ximenes, N.C., de Sousa, C.N., Oliveira, T.Q., Lima, L.L., de Lucena, D.F., Gama, C.S., Macêdo, D.S., Vasconcelos, S.M., 2015. Alpha-lipoic acid alone and combined with clozapine reverses schizophrenia-like symptoms induced by ketamine in mice: participation of antioxidant, nitrergic and neurotrophic mechanisms. Schizophr. Res. S0920-9964 (15), 00190–00195.
Vriend, C., Raijmakers, P., Veltman, D.J., van Dijk, K.D., van der Werf, I.D., Foncke, E. M.J., Smit, J.H., Berendse, H.W., van den Heuvel, O.A., 2014. Depressive symp-toms in parkinson's disease are related to reduced [123I]FP-CIT binding in the caudate nucleus. J. Neurol. Neurosurg. Psychiatry 85 (2), 159–164.
Walther, S., Hüglia, S., Höflea, O., Federspielb, A., Horna, H., Brachta, T., Wiestc, R., Strika, W., Müllera, T.J., 2012. Frontal white matter integrity is related to psy-chomotor retardation in major depression. Neurobiol. Disease 47 (1), 13–19.
Wang, J., Qiao, J., Zhang, Y., Wang, H., Zhu, S., Zhang, H., Hartle, K., Guo, H., Guo, W., He, J., Kong, J., Huang, Q., Li, X., 2014. Desvenlafaxine prevents white matter injury and improves the decreased phosphorylation of the rate-limiting en-zyme of cholesterol synthesis in a chronic mouse model of depression. J. Neurochem. 131 (2), 229–238.
Wolkowitz, O.M., Wolf, J., Shelly, W., Rosser, R., Burke, H.M., Lerner, G.K., Reusa, I., Nelsona, J.C., Epela, E.S., Mellonb, S.H., 2011. Serum BDNF levels before treat-ment predict SSRI response in depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 35, 1623–1630.
Ye, Y., Wang, G., Wang, H., Wang, X., 2011. Brain-derived neurotrophic factor (BDNF) infusion restored astrocytic plasticity in the hippocampus of a rat model of depression. Neurosci. Lett. 503, 15–19.
Yi, L., Liu, B., Li, J., Luo, L., Liua, Q., Geng, D., Tang, Y., Xia, Y., Wub, D., 2014. BDNF signaling is necessary for the antidepressant-like effect of naringenin. Prog. Neuropsychopharmacol. Biol. Psychiatry 48, 135–141.
Yuen, E.Y., Wei1, J., Liu, W., Zhong, P., Li1, X., Yan, Z., 2012. Repeated stress causes cognitive impairment by suppressing glutamate receptor expression and function in prefrontal cortex. Neuron 73 (5), 962–977.
Yulug, B., Ozanb, E., Gönülc, A.S., Kilic, E., 2009. Brain-derived neurotrophic factor, stress and depression: a Minireview. Br. Res. Bull. 78, 267–269.
Zhao, Y., Ma, R., Shen, J., Su, H., Xing, D., Du, L., 2008. A mouse model of depression induced by repeated corticosterone injections. Eur. J. Pharmacol. 581, 113–120.
Zhu, L., Liu, M., Li, H., Liu, X., Chen, C., Han, Z., Wu, H., Jing, X., Zhou, H., Suh, H., Zhu, D., Zhou, Q., 2014. The Different roles of glucocorticoids in the hippocampus and hypothalamus in chronic stress-induced HPA axis hyperactivity. PLOS ONE 9 (5), e97689.