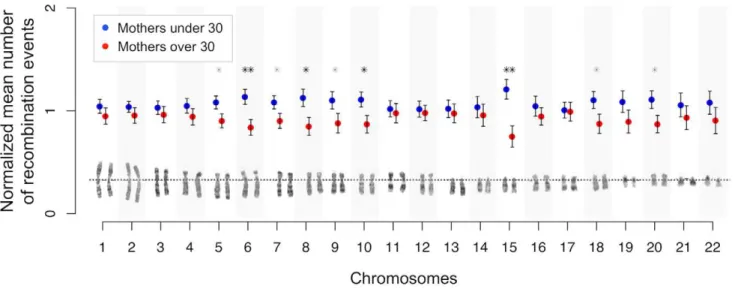
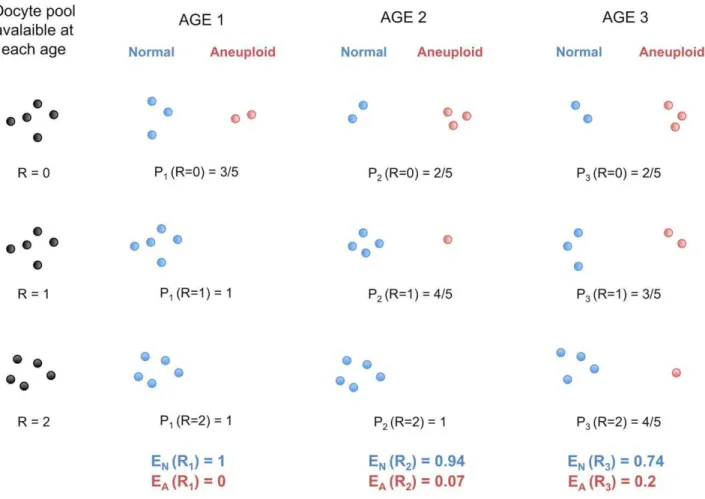
Age-dependent recombination rates in human pedigrees.
Texto
Imagem
Documentos relacionados
Integrase-mediated recombination involving integrons located on multicopy plasmids can generate different recombination products including: (i) free circular DNA molecules
(D) Representation of recombination events along the genome for each branch of the genealogy in (A). True events are shown in blue and events detected by ClonalFrameML are shown in
Assim, verifica-se que foram tomadas medidas importantes ao longo dos últimos anos, tais como, com a entrada dos medicamentos genéricos, o desenvolvimento do sistema de
Mas tem a pretensão de possuir elementos de uma sabedoria que – não sendo conquista sua, mas dom recebido, não sendo desejo de domínio, mas proposta de sentido e serviço – não
A investigação baseia-se no estudo de caso Brisa Inovação que implementou o seu modelo de desenvolvimento de produto assente na sua rede de parceiros: numa primeira fase
Using only the pe_pgrs3 SNPs led to a tree with two large clades (Additional file 6: Figure S5), one containing H37Rv and strains with similar sequence, and the other consistent
Increase in mitotic recombination in diploid cells of Aspergillus nidulans in response to ethidium bromide..
In this work we have demonstrated that yellow fever virus replicon vectors constructed by homologous recombination in yeast can be used for expression of foreign genes in
