Analysis of Parity Between Protein-based Electrophoretic
Methods for the Characterization of Oral
Candida
Species
EAR Rosa, RT Rosa, CV Pereira, MFG Boriollo, JF Höfling
+Laboratório de Microbiologia e Imunologia, Faculdade de Odontologia de Piracicaba, Unicamp, Av. Limeira 901, 13414-900, Piracicaba, SP, Brasil
Electrophoretic studies of multilocus-enzymes (MLEE) and whole-cell protein (SDS-PAGE) were carried out in order to evaluate the parity between different methods for the characterization of five
Candida species commonly isolated from oral cavity of humans by numerical taxonomy methods. The obtained data revealed that sodium dodecyl sulfate polyacrylamide gel electrophoresis is more efficient in grouping strains in their respective species while MLEE has much limited resolution in organizing all strains in their respective species-specific clusters. MLEE technique must be regarded for surveys in which just one species of Candida is involved.
Key words: polyacrylamide gel electrophoresis - multilocus enzyme electrophoresis - Candida -numerical analysis
The yeasts pertaining to the genus Candida
are found dispersed in different epitelial areas of the body, including oral mucosa. In recent years, they have received more attention due to their in-volvement in a increasing number of cases of op-portunist oral infections in patients with Aids and those having immunosuppresive medication. Of epidemiological interest, characterization proce-dures based on molecular fingerprints have been applied in order to establish possible relationships among Candida isolates involved in oral infections (McCullough et al. 1996).
Different types of electrophoretic techniques have been used for the characterization or typing of Candida including electrophoretic separation of chromosomes (Monod et al. 1990, Asakura et al. 1991), DNA fragments (Scherer & Stevens 1987), multilocus-enzymes (Lehmann et al. 1989a, Pujol et al. 1993, Reynes et al. 1996), and whole-cell proteins (Shen et al. 1988, Vancanneyt et al. 1991, 1992, Höfling et al. 1998). The two latter methods have been used successfully for yeast character-ization. The resulting electrophoretic profiles can be plotted into a binary data matrix that, with
com-puter-assisted support, produces comparative re-sults expressed as similarity or cophenetic corre-lation matrices or dendrograms (Kersters 1985).
In this experiment, we compare multilocus-en-zyme electrophoresis (MLEE) and polyacrylamide gel electrophoresis (SDS-PAGE) for their ability to discriminate five Candida species isolated from saliva of healthy subjects.
MATERIALS AND METHODS
Candidastrains - Representative strains of dif-ferent Candida species isolated from human oral cavity and identified by biochemical and physi-ological tests were obtained from the Microbiol-ogy and ImmunolMicrobiol-ogy Laboratory, Dentistry Col-lege of São José dos Campos: C. albicans (97.a, F.72, E.37, 17.b, CBS.562T), C. guilliermondii
(FCF.405, FCF.152, CBS.566T), C. parapsilosis
(21.c, 7.a,CBS.604T), C. krusei (1M.90, 4.c, CBS.573T), C. tropicalis (1.b, FCF.430, CBS.94T). The superscript T in CBS strains indicates that they are the respective type-strains for each species.
Saccharomyces cerevisiae type-strain (CBS.1171T) was included as an extra-generic organism (Costas et al. 1989).
Cell cultivation and whole-cell protein extrac-tion - All strains were grown in 50 ml of Yeast Peptone Dextrose medium (2% dextrose, 2% pep-tone, 1% yeast extract) in a shaker table under 150 rpm, at 30ºC, overnight. The cells were harvested by centrifugation at 2,000 g for 3 min and the pel-lets were washed four times with cold sterile water in order to remove either culture medium traces or extra-cellular metabolites (Woontner & Jaehning 1990). The last washed pellets were transferred to 2 ml microcentrifuge tubes and acid-washed glass
This work received financial support from “Fundação de Amparo à Pesquisa do Estado de São Paulo” and “Fundo de Apoio ao Ensino e Pesquisa, Unicamp”.
+Corresponding author. Fax: +55-19-430.5218. E-mail:
beads (v/v) plus 200 ml of cold sterile water were added. Cells were lysed using a Mini-Bead Beater cell disrupter (Biospec) at 4600 r.p.m., repeating four times of 30 sec at 5-min intervals, and placed in an ice bath. After cell disruption, the micro-cen-trifuge tubes were cenmicro-cen-trifuged at 10,000 g for 2 min, and the supernatant’s protein concentration were determined according to Bradford (1976) and adjusted to 80 µg/ml (Ames 1974). The MLEE supernatants were applied on Whatman 3 filter paper wicks of 5x12 mm (Selander et al. 1986), and for SDS-PAGE technique equal volumes of supernatant and loading buffer of Bruneau and Guinet (1989) (5mM Tris, 2.5% 2-mercap-toethanol, 1.5% SDS, 0.025% bromophenol blue) were combined and heated in a boiling water bath for 10 min.
MLEE and specific-enzyme staining - The elec-trophoreses were carried out using hydrolyzed corn starch Penetrose 30 (Refinações de Milho Brasil) up to a final concentration of 13% (Val et al. 1981) in 1:30 pH 8.0 Tris-citrate buffer (Selander et al. 1986, Caugant & Sandven 1993). Electrophoreses were carried out at 4ºC and 130 V until the bro-mphenol blue migration markers had run at least 80 mm from application point. At this time, the electrophoresis was interrupted and the gels were sliced with 1.2 mm thickness. The gel slices were revealed for enzyme active band detection, accord-ing to Selander et al. (1986) protocols. Enzymatic systems assayed were: alcohol dehydrogenase (ADH-E.C. 1.1.1.1), lactate dehydrogenase (LDH -E.C. 1.1.1.27), malate dehydrogenase (MDH-E.C. 1.1.1.37), isocitrate dehydrogenase (IDH-E.C. 1.1.1.42), glucose-6-phosphate dehydrogenase (G6PDH-E.C. 1.1.1.49), aspartate dehydrogenase (ASDH-E.C. 1.4.3.x), glucose dehydrogenase (GDH-E.C. 1.1.1.47), mannitol dehydrogenase (MADH-E.C. 1.1.1.67), sorbitol dehydrogenase (SDH-E.C. 1.1.1.14), malic enzyme (ME-E.C. 1.1.1.40), aconitase (ACO-E.C. 4.2.1.3), catalase (CAT-E.C. 1.11.1.6), superoxide dismutase (SOD -E.C. 1.15.1.1), glutamate-oxalacetate transami-nase (GOT-E.C. 2.6.1.1), α-esterase (EST-E.C. 3.1.1.1), β-esterase (EST-E.C. 3.1.1.1), leucine aminopeptidase (LAP-E.C. 3.4.1.1), glucosil trans-ferase (GTF-E.C. 2.4.1.11), peroxidase (PO-E.C. 1.11.1.7) e α-amylase (α-AM-E.C. 3.2.1.1).
SDS-PAGE protein analysis - SDS-PAGE pro-tein profiles were obtained after electrophoresis of 50 µl of protein solution in polyacrylamide slab gel with sodium dodecylsulfate (SDS) in a discon-tinuous buffer system (Laemmli 1970) with 4.5% stacking gel and 12.5% running gel. The electro-phoresis was conduced at 125 volts in a cold cham-ber and the gels were stained with Coomassie blue G-250 0.25%. After destaining, the gels were
scanned and the profiles of each lane transferred to a densitometry interface in the SigmaGel soft-ware (Jandel softsoft-ware) where the exact position of the protein peaks were determined.
Computing numerical data - Dendrograms for the different MLEE systems and SDS-PAGE were generated by using the simple matching (SSM) as-sociation coefficient (Sokal & Michener 1958, Sneath & Sokal 1973, Naumov et al 1997), based on band positions calculated by the NTSYS soft-ware package, version 1.70 (Applied Biostatistics, Inc.). For the present study, a SSM of 1.00 repre-sents identical matches (i.e., all the bands match), a SSM of 0.00 represents no matches, and increas-ing intermediatevalues represent increasing pro-portions of matched bands. Dendrograms, repre-sented by non-rooted trees, based on SSM values were generated by the unweighted pair-group arith-metic average (UPGMA) clustering method (Rohlf 1963, Sneath & Sokal 1973, Naumov et al. 1997).
RESULTS
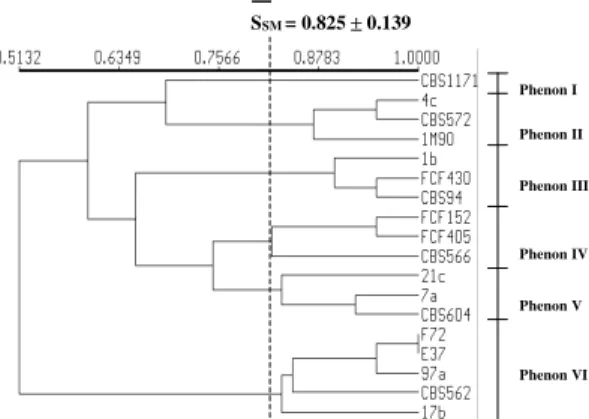
The application of UPGMA clustering pro-duced two similarity dendrograms shown in Figs 1 and 2, in which several clusters (phenons) could be distinguished. These clusters may be defined by their average similarity values (SSM).
Phenons generated by SDS-PAGE
Phenon I: there is the S. cerevisiae type-strain CBS.1171T
Phenon II: there are three strains of C. krusei, with SSM≥ 0.872.
Phenon III: there are three strains of C. tropicalis, with SSM≥ 0.897
Phenon IV: there are three strains of C. guilliermondii, with SSM≥ 0.823
Phenon V: there are three strains of C. parapsilosis, with SSM≥ 0.833
Phenon VI: there are five strains of C. albicans, with SSM≥ 0.833
Interspecific comparison by SDS-PAGE - Among all the species, C. albicans (phenon VI) was the most frequently isolated species and its cluster could be grouped to others with SSM = 0.513.
C. krusei (phenon II) showed some similarity with S. cerevisiae CBS 1171 with SSM = 0.692, and both could be isolated from others with SSM = 0.597.
C. guilliermondii (phenon IV) and C. parapsilosis (cluster V) showed a value of SSM = 0.7749, and these two clusters could be grouped with C. tropicalis (phenon III) with SSM = 0.655.
cerevisiae (CBS 1171) and molecular mass mark-ers were applied in all gels providing mean values SSM = 0.853 and 1.000, respectively.
Enzymatic systems - The one-dimensional elec-trophoreses of protein extracts from 12 Candida
strains, their respective type-strains, and S. cerevisiae type-strain, showed that among twenty assayed enzymes, five did not show any enzymatic activity (ASDH, MADH, SDH, GTF, and α-AM).
Phenon IV: there are three strains of C. parapsilosis (CBS.604T, 21.c, and 7.a) and one
C. albicans strain (17.b), with SSM≥ 0.845 Phenon V: there is a C. krusei strain (4.c) Phenon VI: there is the C. guilliermondii
type-strain CBS.566T
Phenon VII: there are two strains of C. tropicalis
(CBS.94T, and FCF.430), with SSM = 0.917 Phenon VIII: there is the strain 1M.90 of C.
krusei
Phenon IX: there is the C. krusei type-strain (CBS.573T)
Interspecific comparison by MLEE - Exclud-ing phenon I, composed only by C. albicans, and those in which only one strain were detected (phenons III, V, VI, VIII, and IX), all other clus-ters had an impure composition with more than one species component.
DISCUSSION
The analysis of electrophoretic profiles of pro-teins and multilocus-enzymes has allowed the iden-tification, classification of numerous strains, spe-cies and genera of yeasts (Baptist & Kurtzman 1976, Okunishi et al. 1979, Yamazaki & Komagata 1981, Maiden & Tanner 1991, Vancanneyt et al. 1991, 1992).
The reproducibility of electrophoretic profiles on different slab SDS-PAGE gels was evaluated by the inclusion of molecular mass markers, be-sides protein extract of a organism from a non-cor-related genus (Costas et al. 1989, Bruneau & Guinet 1989) and gave similarity correlation values SSM = 0.853 for three repetitions of S. cerevisiae and SSM = 1.000 for three repetitions of molecular mass markers. These values are in agreement with the minimum acceptable proposed by Sneath and Johnson (1972) that was 0.800. The data obtained from grouping of Candida strains based on their electrophoretic profiles showed high level of agree-ment with the inter-specific classification estab-lished by conventional methods. Moreover, the iso-lates of each species showed identical or very simi-lar profiles when compared. This fact suggests that these protein profiles obtained by SDS-PAGE are relatively stable taxonomic characteristics.
As shown in Fig. 1, the use of type-strains al-lowed the identification of clusters at the species level, since the Candida isolates were grouped with their respective type-strains. With regard to clus-ter compositions, the SDS-PAGE technique al-lowed the organization of all isolates in distinct clusters, with similarity coefficients SSM≥ 0.833 for C. albicans, SSM≥ 0.833 for C. parapsilosis, SSM≥ 0.823 for C. guilliermondii, SSM≥ 0.897 for C. tropicalis, and SSM≥ 0.872 for C. krusei. SSM = 0.825 ± 0.139
Phenon I
Phenon II
Phenon III
Phenon IV
Phenon V
Phenon VI
SSM= 0.841±0.091
Phenon I
Phenon II
Phenon III
Phenon IV
Phenon V
Phenon VI
Phenon VII
Phenon VIII
Phenon IX Fig.1: non-rooted dendrogram of similarity among Candida strains grouped by simple matching associative coefficient and UPGMA algorithm from sodium dodecyl sulfate polyacryla-mide gel electrophoresis profiles.
Fig. 2: non-rooted dendrogram of similarity among Candida strains grouped by simple matching associative coefficient and UPGMA algorithm from multilocus enzyme electrophoresis profiles.
Phenons generated by MLEE
Phenon I: there are four strains of C. albicans
(CBS.152T, 97.a, F.72, and E.37) with S SM≥
0.898
Phenon II: there are two strains of C. guilliermondii (FCF.152 and FCF.405) and one C. tropicalis (1.b), with SSM≥ 0.847 Phenon III: there is the S. cerevisiae type-strain
Shechter et al. (1972), using non-denatured acid and basic protein electrophoresis and association coefficient of Jaccard (SJ), that excludes negative matches, obtained a phenogram in which the spe-cies C. albicans, C. krusei and C. parapsilosis com-bined among them with 40% of similarity. The spe-cies C. guilliermondii clustered to this group with 32% and C. parapsilosis was the last one to group, with approximately 25% of similarity. This behav-ior, different from that found in our research, is due to the fact that non-denatured proteins migrate through the gel according to their molecular mass, structural conformation and net charge. In contrast, SDS denatured proteins migrate according to mo-lecular mass only. As momo-lecular mass is more con-served than net charge, electrophoretic profiles based on this criterion should, in theory, detect better taxo-nomic relationships (Kersters 1985).
The systematic proximity between C. krusei and
S. cerevisiae (SSM = 0.692) assessed by SDS-PAGE technique was also observed by Barns et al. (1991) in their analyses based on phylogenetic analysis of 18S ribosomal sub-units RNA genes. Hendricks et al. (1989) support that Candida and
Saccharomyces should have a close phylogenetic relationship, detectable by 18S rRNA sequence analysis.
According to Fig. 2, the MLEE technique grouped most C. albicans strains into a single phenon, except for 17.b strain that was shown to be the less related. These enzymes were able to group all strains of C. parapsilosis with strain 17.b of C. albicans. Such aspect of multispecific cluster gen-erated from MLEE was already observed by Smith et al. (1990), that characterizing different species of
Brettanomyces and Dekkera, obtained a phenogram in which some strains could not be grouped with high similarity values in their respective species-specific clusters and with interference of some strains in other clusters. Jones and Noble (1982) established electrophoretic comparisons among species of der-matophytes based on MLEE technique showing the inclusion of isolates from certain species inner taxa of other species or even of other genera. These au-thors pointed out that this fact may occur when only a few isolates of each species are included in the surveys. Boerlin et al. (1995) used 16 enzymatic systems for characterizing 21 genetically atypical strains of chlamydospore-forming and germ tube-positive C. albicans recovered from human immu-nodeficiency virus-positive drug users, and demon-strated that some of these strains were grouped in different clusters, showing high diversity on allelic composition.
Extensive enzyme heterogeneity among Can-dida or other yeast genera had already been ob-served by other groups of researchers that pointed
out that it may occur increasing the possibility of distributing such specimens in various groups or clusters (Lehmann et al. 1989a, 1989b, Caugant & Sandven 1993, Naumov et al. 1997). Lehmann et al. (1991) related the phenomenon of isoenzymatic patterns changing of C. albicans during its conser-vation in laboratories, what could increase the ap-parent polymorphism. Pujol et al. (1997) found atypical strains of C. albicans in Aids patients, showing diverse allelic polymorphism.
When comparing the results assessed by SDS-PAGE and MLEE, it can easily be seen that the first one is more useful for grouping isolates in their respective species, maybe due to the expression of species-specific bands while the second one per-haps better explores the variability at a sub-spe-cific level, being useful for analyses of genetic polymorphism among strains of a certain Candida
species.
In order to ensure whether or not the UPGMA algorithm assesses resemblance between two OTUs in the dendrogram constructions, a product-mo-ment correlation coefficient was computed between the elements SJK of the original similarity matrix S and cophenetic values CJK of the matrix C de-rived from the dendrogram. The cophenetic corre-lation coefficient is a measure of the agreement between similarity values implied by the dendro-gram and those of the original similarity matrix (Sokal & Rohlf 1962). These coefficient had val-ues rCS = 0.928 for SDS-PAGE and rCS = 0.932 for MLEE, that range between 0.60 and 0.95 (Sneath & Sokal 1973) or higher than 0.90 (Sokal & Rohlf 1970), considered acceptable, corroborat-ing by this way, with the finds of Farris (1969), that pointed out the fact that UPGMA algorithm always maximizes rCS values.
The protein profile analysis by SDS-PAGE improves the knowledge about the taxonomic re-lationships among oral yeasts. This method shows good reproducibility and allows collection of use-ful information for numerical analysis. This meth-odology brings relevant information in systematic evaluation of related species. We propose that the grouping of Candida species by MLEE patterns from the assayed enzymes is not efficient when only based on a few isolates from more than one species, regarding such resource for surveys con-duced with a single species of Candida, for what, the MLEE technique had already proved to be a useful method for systematic or epidemiological purposes.
REFERENCES
Asakura K, Iwaguchi SI, Homma M 1991. Electro-phoretic karyotypes of clinically isolated yeasts
Candida albicans and C. glabrata. J Gen Microbiol 137: 2531-2538.
Baptist JN, Kurtzman CP 1976. Comparative enzyme patterns in Cryptococcuslaurentii and its taxonomic varieties. Mycologia68: 1195-1203.
Barns SM, Lane DJ, Sogin ML, Bibeau C, Weisburg WG 1991. Evolutionary relationships among patho-genic Candida species and relatives. J Bacteriol173: 2250-2255.
Boerlin P, Boerlin-Petzold F, Durussel C 1995. Cluster of oral atypical Candida albicans isolates in a group of human immunodeficiency virus-positive drug users. J Clin Microbiol33: 1129-1135.
Bruneau S, Guinet R 1989. Rapid identification of medi-cally important yeast by electrophoretic protein pat-terns. FEMS Microbiol Letters58: 329-334. Caugant DA, Sandven P 1993. Epidemiological
analy-sis of Candida albicans strains by multilocus en-zyme electrophoresis. J Clin Microbiol31: 215-220. Costas M, Holmes B, Wood AC, On SLW 1989. Nu-merical analysis of electrophoretic patterns of
Providencia rettgeri from human faeces, urine and other specimens. J Appl Bacteriol 67: 441-452. Farris JS 1969. On the cophenetic correlation coefficient.
Syst Zool18: 279-85.
Hendricks L, Goris A, Neefs J 1989. The nucleotide se-quence of the small ribosomal subunit RNA of the yeast Candida albicans and the evolutionary posi-tion of the fungi among the eukaryotes. Syst Appl Microbiol12: 223-229.
Höfling JF, Rosa EAR, Rochelle SLA, Spolidório DMP, Moreira D 1998. Numerical analysis variations of SDS-PAGE protein patterns using different culture media for the cultivation of Candida from the oral cavity. Rev Microbiol28: 79-84.
Jones MG, Noble WC 1982. An electrophoretic study of enzymes as a tool in the taxonomy of the der-matophytes. J Gen Microbiol20: 1101-1107. Kersters K 1985. Numerical methods in the
classifica-tion of bacteria by protein electrophoresis. In M Goodfellow, D Jones, FG Priest (eds), Computer Assisted Bacterial Systematic, Academic Press, En-gland, p. 337.
Laemmli UK 1970. Cleavage of strutural proteins dur-ing the assembly of the head of bacteriophage T4.
Nature227: 680-685.
Lehmann PF, Hsiao CB, Salkin IF 1989a. Proteins and electrophoresis profiles of selected Candida species.
J Clin Microbiol27: 400-404.
Lehmann PF, Kemker BJ, Hsiao CB, Dev S 1989b. Isoenzyme biotypes of Candida species. J Clin Microbiol 27: 2514-2521.
Lehmann PF, Wu LC, Mackenzie DW 1991. Isoenzyme changes in Candidaalbicans during domestication.
J Clin Microbiol29: 2623-2625.
Maiden MFJ, Tanner A 1991. Identification of oral yeasts by polyacrylamide gel electrophoresis. Oral Microbiol Immunol6: 187-190.
McCullough MJ, Ross BC, Reade PC 1996. Candida albicans, a review of its history, taxonomy,
viru-lence attributes, and methods of strain differentia-tion. Int J Oral Maxillofac Surg25: 136-144. Monod M, Porchet F, Baudraz R, Frenk E 1990. The
identification of pathogenic yeast strains by electro-phoretic analysis of their chromosomes. J Med Microbiol32: 123-129.
Naumov GI, Naumova ES, Sniegowiski PD 1997. Dif-ferentiation of European and far east Asian popula-tions of Saccharomyces paradoxus by allozyme analysis. Int J System Bacteriol 47: 341-344. Okunishi M, Yamada K, Komagata K 1979.
Electro-phoretic comparison of enzymes from basidi-omycetes in different stages of development. J Gen Appl Microbiol25: 329-334.
Pujol C, Renaud F, Mallie M, de Meeus T, Bastide JM 1997. Atypical strains of Candida albicans recov-ered from AIDS patients. J Med Vet Mycol35: 115-121.
Pujol C, Reynes J, Renaud F, Raymond M, Tibayrenc M, Ayala FJ, Janbon F, Mallie M, Bastide JM. 1993. The yeast Candida albicans has a clonal mode of reproduction in a population of infected human im-munodeficiency virus-positive patients. Proc Natl Acad Sci USA90: 9456-9459.
Reynes J, Pujol C, Moreau C, Mallie M, Renaud F, Janbon F, Bastide JM. 1996. Simultaneous carriage of Candida albicans strains from HIV-infected pa-tients with oral candidiasis, multilocus enzyme elec-trophoresis analysis. FEMS Microbiol Letters137: 269-273.
Rohlf FJ 1963. Classification of Aedes by numerical taxonomic methods (Diptera, Culicidae). Ann Entomol Soc Amer56: 798-804.
Scherer S, Stevens DA 1987. Application of DNA typ-ing methods to epidemiology and taxonomy of Can-dida species. J Clin Microbiol25: 675-679. Selander RK, Caugant DA, Ochman DA, Musser JM,
Gilmour MN, Whittam TS. 1986 Methods of multilocus enzyme electrophoresis for bacterial population genetics and systematics. Appl Environ Microbiol51: 873-884.
Shechter Y, Landau JW, Dabrowa N 1972. Compara-tive electrophoresis and numeral taxonomy of some
Candida species. Mycologia 64: 841-853. Shen HD, Choo KB, Tsai WC 1988. Differential
identi-fication of Candida species and other yeasts by analysis of [35S] - metionine labeled polypeptide profiles. AnalytBiochem175: 548-555.
Smith MT, Yamazaki M, Poot GA 1990. Dekkera, Brettanomyces and Eeniella, electrophoretic com-parison of enzymes and DNA-DNA homology. Yeast 6: 299-310.
Sneath PHA, Johnson R 1972. The influence on numeri-cal taxonomy similarities of errors in microbial tes-tes. J Gen Microbiol72: 248-255.
Sneath PHA, Sokal RQ 1973. Numerical Taxonomy,
Freeman, San Francisco, 573 pp.
Sokal RR, Michener CD 1958. A statistical method for evaluating systematic relationships. Univ Kansas Sci Bull38: 1409-1438.
Sokal RR, Rohlf FJ 1970. The intelligent ignoramus, an experiment in numerical taxonomy. Taxon19: 305-319.
Val AL, Schwantes AR, Schwantes MLB, de Luca PH 1981. Amido hidrolisado de milho como suporte eletroforético. Ci Cult33: 992-996.
Vancanneyt M, Lerberge EV, Berny JF, Hennebert GL, Kersters K 1992. The application of whole-cell pro-tein electrophoresis for the classification and identi-fication of basidiomycetous yeast species. Antonie van Leeuwenhoek61: 69-78.
Vancanneyt M, Pot B, Hennebert G, Kersters K 1991. Differentiation of yeast species based on electro-phoretic whole-cell protein patterns. Syst Appl Microbiol 14: 23-32.