Fatores Clínicos, Nutricionais, Comportamentais, Polimorfismo
Pro12Ala do Receptor Ativado por Proliferadores de
Peroxissomos Gama (PPARG) e Risco de Câncer de Mama:
Um estudo caso-controle
GEÓRGIA DAS GRAÇAS PENA
Fatores Clínicos, Nutricionais, Comportamentais, Polimorfismo Pro12Ala do Receptor Ativado por Proliferadores de Peroxissomos Gama (PPARG) e Risco de Câncer de Mama:
Um estudo caso-controle
Dissertação de mestrado apresentada ao Núcleo de Pesquisas em Ciências Biológicas da Universidade Federal de Ouro Preto, como requisito parcial para obtenção do grau de Mestre em Ciências Biológicas, na área de concentração em Biologia Molecular.
Orientadora: Renata Nascimento Freitas Co-orientador: George Luiz Lins Machado Coelho
DEDICATÓRIA
Dedico esse trabalho aos meus pais, Glória Geralda Dias Pena e David Mendes Pena
AGRADECIMENTOS
À Deus, pelo consolo nos momentos difíceis e pela força e alegria da caminhada.
Aos meus pais, Glória e David, pelo exemplo de vida, honestidade, caráter, como também pelo incentivo e apoio à minha formação acadêmica.
À Professora Renata N Freitas pela oportunidade, pelo exemplo de competência e profissional amante da pesquisa.
Ao Professor George Luis L Machado-Coelho pelo auxílio e ensinamento no desenvolvimento da estatística.
Aos meus irmãos Gêner, Dárlen e Breno, por fazerem parte da minha vida, por sermos um laço eterno de amor.
Ao Paulo Renato, pela paciência, compreensão e amor a mim dispensados. Niño, eu achei que tinha tido conquistas importantes... até conhecê-lo!!
Às mulheres voluntárias do presente estudo, por doarem seu tempo e sua paciência na aplicação do questionário e por compartilharem suas mágoas, suas dificuldades e suas estórias conosco.
Aos Mastologistas da MOV pela paciência e atenção durante as cirurgias, discussão e retiradas de dúvidas, além de compartilharem seu espaço de trabalho!!
À Lílian, pela competência e garra que demonstrou durante toda a coleta de dados. Ganhamos muito com sua presença! Obrigada pela amizade e pelo exemplo de ser humano, aprendi muito contigo!!
À Débora e à Rachel pelo auxílio na coleta de dados. Obrigada pela força!
À equipe do Bloco Cirúrgico por terem nos recebido tão abertamente, por toda a dedicação à coleta de material biológico, pela amizade e confiança. Minha eterna gratidão!!
À equipe do Banco de Sangue, por terem compartilhado tão amorosamente seus equipamentos, espaço físico e pessoal para coleta de sangue. Vocês foram maravilhosos!!
À equipe do ambulatório, que compartilhou conosco seus dias cheios de trabalho, e, mesmo assim foram abertos e amorosos.
À equipe do SAME por serem tão competentes na manutenção dos prontuários da MOV e pelo cafezinho delicioso que nos oferecia!!
À equipe da internação por sempre informarem com boa vontade o paradeiro das mulheres que chegavam para a cirurgia.
Aos funcionários do almoxarifado que, na hora do aperto, tanto contribuíram para que continuássemos nosso trabalho!!
Ao Angelo que esteve comigo durante a etapa anterior a esse projeto me ensinando as práticas da biologia molecular.
À Natália e à Denise pela imprescindível ajuda na extração de DNA e na digitação dos dados de consumo alimentar.
À Fabíola e Mirelle que também contribuíram intensamente na árdua tarefa da digitação dos dados de consumo.
Aos amigos do Laboratório de Epidemiologia Molecular: Bruno, Michael, Carlos e Amanda pela companhia e amizade. Em especial à Cristiane e ao Dalton, que, além de contribuir muito para o nosso convívio, são amigos para toda a vida!!
Aos Laboratórios de Nutrição Experimental e Bromatologia da Escola de Nutrição, por cederem sua ajuda, seu tempo e seu espaço para as nossas necessidades quando foram necessárias. Obrigada, Prof. Marcelo, Jair, Emerson, Allan, Lerê!!!
Ao laboratório de Microbiologia dos Alimentos da Escola de Nutrição, em especial ao Prof. Sabione e ao Caetano.
Ao Dr. Carlos Eduardo pela imensa ajuda na classificação dos tumores.
Aos Profs. do Mestrado que muito contribuíram para a minha formação. Serei eternamente grata!!
Aos amigos do mestrado, os quais, nunca esquecerei pelo apoio e amizade!!
Ao Luis, que além de ser o técnico do laboratório, consertava todos os equipamentos que precisávamos e nos salvava nos momentos de maior aperto!!
À D. Erenice e D. Eliete, que com carinho e dedicação nos acompanhou durante essa jornada, dando-nos força e incentivo nos momentos de desânimo!!
Às irmãs Selvagens e à eterna República Bicho do Mato, pela amizade, pelo apoio, pela confiança.... AMO VOCÊS!!
RESUMO
ABSTRACT
Breast Cancer is the second most common cancer in the world and the first among women, with an increasing number of new cases each year, being considered a worldwide public health problem. As a multi-factorial pathology, researches have investigated the role of diet components, environmental, and, genetic factors and their relationship in the modulation of breast cancer. The PPARG is a gene involved in the energetic metabolism, adipose tissue metabolism and in inflammation. It presents a polymorphism, Pro12Ala, which constitutes in the substitution of a proline by an alanine in the exon B, being described in the literature as possibly influencing the risk of cancer. A masked case-control study, hospital based, was conducted to evaluate the associations between clinical and nutritional variables and the Pro12Ala polymorphism with the breast cancer risk. Clinical, obstetrics, dietary and lifestyle data were obtained from the participants, along with the genotyping of the polymorphism. The sample was composed by 654 women, from 12 to 92 years old, most part been not single (75,1%), employed in domestic services (54,5%) and with low education level (64,0%). The distribution of the participants according with the presence of the breast cancer, breast benign disease or controlswas 255 (39,0%), 220 (36,6%) and 179 (27,4%), respectively. The factors that showed to increase the risk in the unvariate analysis were: women aging more than 51 years old, low income, nuliparity, absence of breast-feeding, menopause and sedentary. In the evaluation of the relationship between the polymorphism and anthropometrics factors, it was observed that the CG+GG carriers had an average height higher than the CC carriers, irrespective to diagnoses. The evaluation of the dietary ingestion did not show any relation with the breast cancer risk, independent of the genotype. In the logistical regression analysis, the variables that explain the occurrence of breast cancer in this population are sedentariness, low income, breast-feeding absence and menopause. The stratification according to menopausal status showed that alcoholism is a risk factor in the post menopause and the breast-feeding absence is a risk factor for breast cancer in the pre-menopause. So, it was observed that the risk of breast cancer in this population is related to hormonal factors such as menopause and absence of breastfeeding), and also to environmental factors and lifestyle such as, low income and sedentariness, not being directly affected by the dietary ingestion of calories and lipids, nor related to the PPARG Pro12Ala polymorphism.
LISTA DE FIGURAS
FIGURA 1 Domínios funcionais do PPAR ... ...45 FIGURA 2 Organização do gene PPARG humano...47 FIGURA 3 Posições das mutações/polimorfismos do PPARG...48
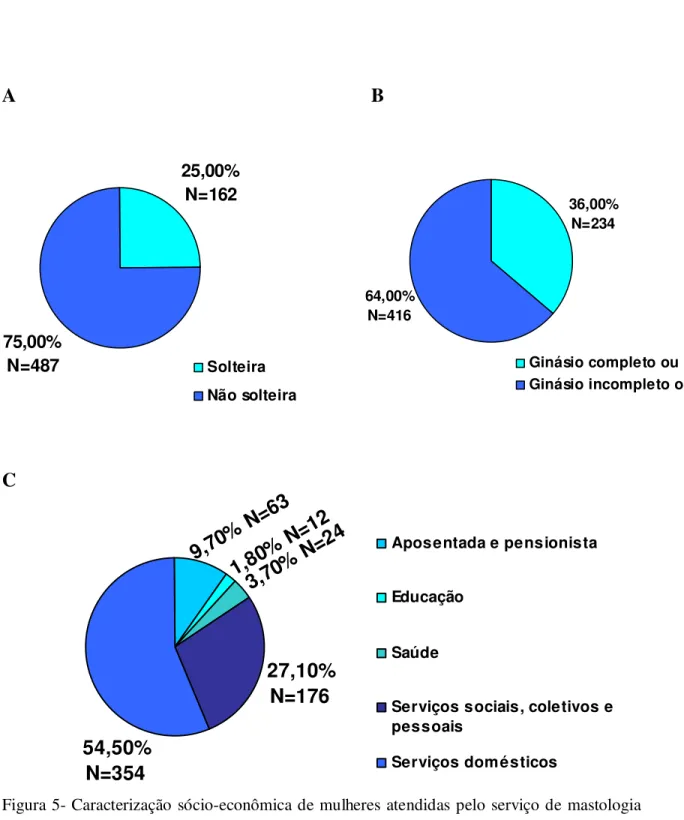
FIGURA 4 Histograma da distribuição da idade em anos de 651 mulheres atendidas pelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte,
2006...80 FIGURA 5 Caracterização sócio-econômica de mulheres atendidas pelo serviço de
LISTA DE QUADROS
QUADRO 1 Classificação do índice de massa corpórea de adultos...66
QUADRO 2 Valores estabelecidos para %GC para sexo feminino. ... 67
QUADRO 3 Estadiamento patológico para câncer de mama (TNM) ... 68
QUADRO 4 Grupamento por estádios ... 69
LISTA DE TABELAS
TABELA 1 Caracterização das variáveis clínicas e gineco-obstétricas de mulheres atendidaspelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte - MG, 2006...83 TABELA 2 Análise univariada da distribuição de acordo com variáveis sócio-demográficas
de 255 mulheres com câncer de mama e 179 controles atendidas pelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte - MG, 2006. ... 88 TABELA 3 Análise univariada da distribuição, de acordo com variáveis clínicas e gineco- obstétricas, de 255 mulheres com câncer de mama e 179 controles atendidas pelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte - MG, 2006...91 TABELA 4 Análise univariada dos valores médios e mediana (Md) das variáveis clínicas e
gineco-obstétricas de 255 mulheres com câncer de mama e 179 controles atendidas pelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte- MG, 2006. ... 93 TABELA 5 Análise univariada dos valores médios e mediana (Md) das variáveis
TABELA 8 Análise univariada da distribuição das freqüências dos alelos e genótipos para polimorfismo Pro12Ala do gene PPARG em 219 mulheres com câncer de mama e 169 controles atendidas no serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte - MG, 2006. ... 100 TABELA 9 Análise multivariada de valores médios ± desvios-padrão das variáveis
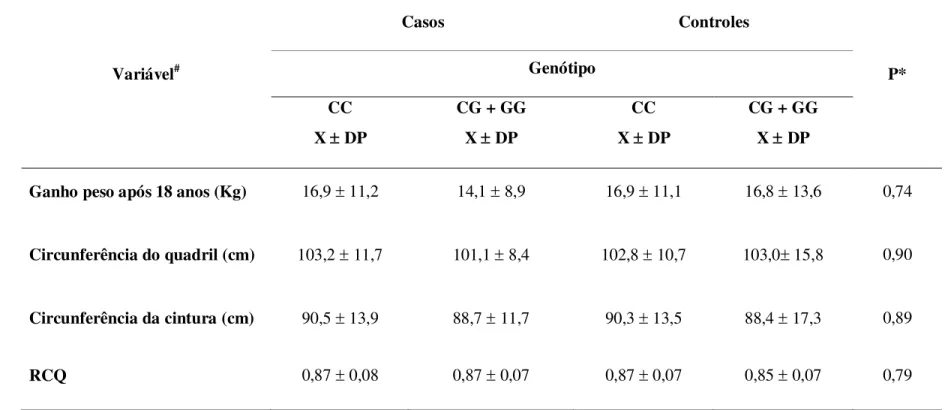
antropométricas de acordo com os genótipos para o polimorfismo Pro12Ala do gene PPARG em 255 mulheres com câncer de mama e 179 controles atendidas no serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte - MG, 2006. ... 104 TABELA 10 Análise descritiva dos valores médios das variáveis de consumo alimentar e
análise univariada das posições médias ou ranqueamento dos genótipos para o polimorfismo Pro12Ala do gene PPARG em 51 mulheres com câncer de mama e 114 sem câncer de mama atendidas no serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte – MG, 2006... 106 TABELA 11 Análise univariada da distribuição, conforme os quartis de consumo de
nutrientes, de 51 mulheres com câncer de mama e 118 mulheres sem câncer de mama, atendidas pelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte – MG, 2006. ... 109 TABELA 12 Análise univariada da distribuição por freqüência dos genótipos para o
polimorfismo Pro12Ala do gene PPARG e consumo alimentar em 51 mulheres com câncer de mama e 114 sem câncer de mama, atendidas pelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte - MG 2006....111 TABELA 13 Modelo de regressão logística múltipla ajustada para os fatores de confusão
TABELA 14 Análise univariada estratificada pelo estado menopausal de variáveis clínicas, gineco-obstétricas e comportamentais entre 255 mulheres com câncer de mama e 179 controles atendidas pelo serviço de mastologia da Maternidade Odete Valadares, Belo Horizonte – MG, 2006. ... 118 TABELA 15 Associação bivariada por gradiente de risco para amamentação e atividade
física para 255 mulheres com câncer de mama e 179 controles atendidas pelo serviço de mastologia da Maternidade Odete Valadares, 2006. ... 122 TABELA 16 Associação bivariada por gradiente de risco para amamentação e porcentagem
LISTA DE ABREVIATURAS E SÍMBOLOS
15PGJ2 - 15-deoxy 12,14-prostaglandina J2 %GC - Porcentagem de gordura corporal AA – Ácido araquidônico
ACS – Enzima acil-Coa sintase ADH – Dehidrogenase alcoólica AF - Atividade física
AF1 – Região de ativação transcricional independente de ligante AINE – Antiinflamatórios não esteroidais
AP-1 – Proteína ativadora 1
aP2 - Proteína ligadora de ácidos graxos BRCA1 – Breast Câncer 1 gene
BRCA2 – Breast Câncer 2 gene
CA - Grupo de mulheres que apresentam câncer de mama (casos) CC - Circunferência da cintura
CG – Indivíduo ou grupo de indivíduos que apresenta uma citosina e uma guanina no locus do PPARG2.
GG – Indivíduo ou grupo de indivíduos que apresenta duas citosinas no locus do PPARG2. CM - Câncer de mama humano
CO – Grupo de mulheres sem câncer de mama (controle) CONEP - Comitê Nacional de Ética em Pesquisa
CQ – Circunferência do quadril COX1 – Cicloxigenase 1 COX2 – Cicloxigenase 2
CDC - Centro para Prevenção e Controle de Doenças dos Estados Unidos da América CDK - Ciclina dependente de quinase
CELAFISCS - Centro de Estudos do Laboratório de Aptidão Física de São Caetano do Sul CTO - Contraceptivos orais
DBD - Domínio de ligação ao DNA DBM – Doença benigna da mama DHA – Ácido Docosahexaenóico
E2F - Fator de transcrição E2F EPA - Ácido Eicosapentaenóico
EUROCARE - European Cancer Registries Study on Cancer FAO – Food and Agriculture Organization
FATP-1 - Proteína transportadora de ácidos graxos FDA – Food and Drug Administration
FHEMIG - Fundação Hospitalar do Estado de Minas Gerais GET - Gasto energético total
GST - Glutationa-S-transferase HDL – Lipoproteína de alta densidade HF – História família de câncer de mama
HMG – Coa - 3-hidroxi-3metilglutaril coenzima-A redutase IARC - International Agency for Research on Cancer IC – Intervalo de confiança
IBGE – Instituto Brasileiro de Geografia e Estatística IBIS-II –Internacional Breast Intervention Study II IL-6 – Interleucina 6
IL-8 – Interleucina 8
IMC - Índice de massa corporal INCA - Instituto Nacional do Câncer
iNOS – Enzima óxido nítrico sintase indutível
IPAQ - The International Physical Activity Questionnaire LBD – Região no DNA de interação com ligantes LC-PUFA – Ácidos graxos poliinsaturados LC-PUFA n-3 – Ácido graxo poliinstaturado n-3 LC-PUFA n-6 – Ácido graxo poliinstaturado n-6 LDL-c – Lipoproteína de baixa densidade LOX – Lipoxigenase
LPL – Lípase lipoproteica M - Macrófagos
Md – Valor da mediana da distribuição MMG - Mamografia
MOV - Maternidade Odete Valadares
N – Número da amostra
NIH - National Institutes of Healthy ONU - Organização das Nações Unidas OR – Odds Ratio
ODC – Ornitina descarboxilase
P – Valor da comparação entre grupos, valor para significância estatística PAGE – Eletroforese em gel de poliacrilamida
PCR - Reação em cadeia da polimerase PKC – Proteína C quinase
PLA2 – Fosfolipases de membrana
PPAR - Receptores ativados por proliferação de peroxissomos PPARA - Receptores ativados por proliferação de peroxissomos alfa PPARB - Receptores ativados por proliferação de peroxissomos beta PPARG - Receptores ativados por proliferação de peroxissomos gama PPARG1 - Receptores ativados por proliferação de peroxissomos gama 1 PPARG2 - Receptores ativados por proliferação de peroxissomos gama 2 PPARG3 - Receptores ativados por proliferação de peroxissomos gama 3 PPARG4 - Receptores ativados por proliferação de peroxissomos gama 4 PPRE- Elementos responsivos do PPAR
QSFA - Questionário Semiquantitativo de Freqüência Alimentar RCQ – Relação cintura - quadril
RDA - Recomended Dietary Allowance RE – Receptor de estrógeno
RFLP - Polimorfismo de tamanho de fragmento de restrição RP – Receptor de progesterona
RXR – Receptor de ácido retinóico
SATR – Study of Tamoxifen and Raloxifene SERM – Modulador do receptor de estrógeno SAH - Adenosilhomocisteína sulfato
SAM - Adenosilmetionina sulfato SMRT – Co-repressores do PPARG
TNF - Fator de Necrose Tumoral alfa
TNM – Tumor, linfonodo e metástase (Classificação de tumores malignos) TRH - Terapia de Reposição Hormonal
TACO – Tabela Brasileira de Composição de Alimentos TCLE – Termo de Consentimento Livre e Esclarecido TZD – Tiazolidinedionas
UICC - União Internacional Contra o Câncer VLDL – Lipoproteína de muito baixa densidade X- Valor médio da distribuição
SUMÁRIO
1 INTRODUÇÃO...23
2 REVISÃO BIBLIOGRÁFICA...25
2.1 ASPECTOS EPIDEMIOLÓGICOS DO CÂNCER...25
2.2 O CÂNCER DE MAMA (CM) ...26
2.2.1 ASPECTOS EPIDEMIOLÓGICOS...26
2.2.2 DETECÇÃO, DIAGNÓSTICO, ESTADIAMENTO...28
2.2.3 FATORES DE ISCO...29
2.2.3.1 FATORES SÓCIO-EMOGRÁFICOS...29
2.2.3.2 FATORES GINECO-OBSTÉTRICOS E HORMÔNIOS EXÓGENOS...29
2.2.3.3 COMPOSIÇÃO CORPORAL E FATORES ANTROPOMÉTRICOS...31
2.2.3.4 FATORES DIETÉTICOS ...33
2.2.3.5 INGESTÃO DE ÁLCOOL...37
2.2.3.6 ATIVIDADE FÍSICA...38
2.2.3.7 HISTÓRIA PESSOAL DE DOENÇA BENIGNA DA MAMA, HISTÓRIA FAMILIAR DE CÂNCER DE MAMA E DENSIDADE MAMOGRÁFICA.38 2.2.3.8 OUTROS FATORES...39
2.2.4 EPIDEMIOLIA MOLECULAR DO CM...41
2.3 RECEPTORES ATIVADOS POR PROLIFERADORES DE PEROXISSOMOS- PPAR...44
2.3.1 LIGANTES DO PPAR E SEU PAPEL BIOLÓGICO...49
2.3.2 PPAR, METABOLISMO DE LIPÍDIOS E INFLAMAÇÃO...50
2.3.5 PPARG E O CÂNCER DE MAMA...55 2.3.6 POLIMORFISMO PRO12ALA DE PPARG 2...56
3 OBJETIVOS...58
3.1 GERAIS...58
3.2 ESPECÍFICOS...58
4 CASUÍTICA E MÉTODOS...58
4.1 DESENHO DO ESTUDO ...59
4.1.1 ÁREA E POPULAÇÃO...59
4.1.2 DELINEAMENTO EPIDEMIOLÓGICO...59
4.1.3 CÁLCULO DO TAMANHO DA AMOSTRA...60
4.1.4 CRITÉRIOS DE EXCLUSÃO DO ESTUDO...60
4.1.4.1 CRITÉRIOS DE EXCLUSÃO PARA AVALIAÇÃO DIETÉTICA....61
4.1.4.2 CRITÉRIOS DE EXCLUSÃO PARA MULHERES CONTROLES...61
4.1.5 COLETA DE DADOS...62
4.1.5.1 VARIÁVEIS SÓCIO-DEMOGRÁFICAS...62
4.1.5.2 VARIÁVEIS CLÍNICAS...64
4.1.5.3 VARIÁVEIS GINECOLÓGICAS E OBSTÉTRICAS...64
4.1.5.4 VARIÁVEIS ANTROPOMÉTRICAS...66
4.1.5.5 VARIÁVEIS DE ESTILO DE VIDA...69
4.1.5.6 OBTENÇÃO DE AMOSTRAS BIOLÓGICAS...72
4.1.5.7 EXTRAÇÃO DE DNA...73
4.1.5.7.1 EXTRAÇÃO DO DNA DE AMOSTRAS PROVENIENTES
DE CÉLULAS DA MUCOSA BUCAL...73
4.1.5.7.2 EXTRAÇÃO DO DNA DAS AMOSTRAS DE SANGUE...73
4.1.6 GENOTIPAGEM DO POLIMORFISMO PRO12ALA DE PPARG POR PCR-RFLP...75
4.1.6.1 REAÇÃO EM CADEIA DA POLIMERASE (PCR)...75
4.1.6.2 DIGESTÃO COM ENZIMA DE RESTRIÇÃO E ELETROFORESE EM GEL
DE POLIACRIAMIDA...76
4.2 PROCESSAMENTO E ANÁLISE DOS DADOS...76
4.2.1 BANCO DE DADOS E ANÁLISE ESTATÍSTICA...76
4.2.2 ANÁLISE DOS DADOS DE CONSUMO ALIMENTAR...77
4.2.3 ANÁLISE DOS DADOS GENÉTICOS...77
4.2.4 REPRODUTIBILIDADE...78
4.3 QUESTÕES ÉTICAS E LEGAIS...78
5 RESULTADOS...79
5.1 FATORES DE RISCO PARA O CÂNCER DE MAMA...87
5.2 MODELO DE REGRESSÃO LOGÍSTICA MÚLTIPLA...115
5.3 ANÁLISE ESTRATIFICADA PELO ESTADO MENOPAUSAL...117
5.4 GRADIENTE DE RISCO...119
6 DISCUSSÃO...121
7 CONCLUSÕES...137
REFERÊNCIAS BIBLIOGRÁFICAS...139
APÊNDICES...175
23 Os SNPs (Single Nucleotide Polymorphisms) ou polimorfismos de nucleotídeo único, são variações de uma única base que ocorrem na seqüência genômica. Os SNPs são responsáveis por 90% das variações no genoma humano (NIH, 2003), e apresentam freqüência maior que 1% na população geral (CARGILL et al., 1999). Na era pós-genômica, os SNPs estão sendo intensivamente estudados para uma melhor compreensão das bases moleculares de doenças complexas (ZHU et al., 2004).
24 As interações gene-nutrientes são recíprocas e numa abordagem de estudos de sistemas, tanto a regulação da maquinaria genômica por nutrientes, constituintes alimentares e/ou seus metabólitos quanto a resposta específica conferida pelo genótipo aos componentes da dieta, são investigados graças aos avanços de diferentes áreas do conhecimento como genômica funcional, genética, bioquímica, fisiologia, nutrição, epidemiologia, bioinformática, etc.
Enfim, os componentes dietéticos têm influência fundamental na saúde através de alterações na expressão gênica por mecanismos genéticos e/ou epigenéticos. Essas modificações, herdadas na transmissão da informação genética podem ser importantes causas da susceptibilidade ao câncer, incluindo genes envolvidos no metabolismo de carcinógenos, na manutenção dos níveis hormonais para os cânceres responsivos a hormônios ou genes envolvidos no metabolismo de nutrientes.
Embora a nível celular o câncer seja reconhecido etiologicamente como patologia de disfunção gênica, existem vários estudos epidemiológicos que demonstram resultados diferenciados por causa da modulação por fatores ambientais (NIH, 2003).
25
2.1 ASPECTOS EPIDEMIOLÓGICOS DO CÂNCER
O câncer tem aumentado de maneira considerável em todo o mundo, principalmente a partir do século passado, configurando-se na atualidade como um dos mais importantes problemas de saúde pública mundial (GUERRA et al., 2005). Os cânceres são responsáveis por mais de seis milhões de óbitos a cada ano, representando cerca de 12% de todas as causas de morte do mundo, tanto em países desenvolvidos quanto em desenvolvimento, sendo que nestes são diagnosticados cinco milhões e meio dos dez milhões de novos casos anuais de câncer (WHO, 2002).
A distribuição epidemiológica do câncer no Brasil sugere uma transição, envolvendo um aumento entre os tipos de câncer normalmente associados a alto status sócio-econômico – câncer de mama, próstata e reto e, simultaneamente, a presença de taxas de incidência persistentemente elevadas de tumores geralmente associados com a pobreza – câncer de colo de útero, pênis, estômago e cavidade oral (KOIFMAN S. e R., 2003).
Segundo estimativas do Instituto Nacional do Câncer, ocorreriam em 2006, mais de 472 mil casos novos de câncer, 234 mil entre os homens e 238 mil entre as mulheres. Os tumores mais incidentes na população brasileira seriam os de pele não melanoma (116 mil), mama feminina (49 mil), próstata (47 mil), pulmão (27 mil) e cólon e reto (25 mil) (INCA, 2006).
26
2.2 O CÂNCER DE MAMA (CM)
2.2.1 ASPECTOS EPIDEMIOLÓGICOS
No ano 2000, houve mais novos casos de câncer de mama (CM) em países desenvolvidos (579.000) que em todos os países em desenvolvimento juntos (471.000) e também mais mortes por esse tipo de câncer (189.000 e 184.000, respectivamente) (IARC, 2003). O CM permanece o segundo tipo de câncer mais freqüente no mundo e o primeiro entre as mulheres (INCA, 2006), o que o confirma como um problema importante de saúde pública (SELVAN et al., 2004).
Para entender as inter-relações entre incidência, mortalidade e sobrevida e suas conseqüências, um estudo utilizando dados de sobrevida da EUROCARE e de mortalidade da WHO, estimou a incidência de CM em 10 países europeus de 1970 a 2005. Neste estudo foi demonstrado um aumento da incidência com aumento da sobrevida e declínio da mortalidade na Suécia e Finlândia; aumento da sobrevida com aumento da incidência e mortalidade estável na Dinamarca, Países Baixos e França; aumento da sobrevida, incidência estável e diminuição da mortalidade na Espanha e Itália; sobrevida estável, aumento da incidência e mortalidade na Estônia (POPAT e SMITH, 2006).
Portanto, na maioria dos países a sobrevida aumentou, indicando um aspecto benéfico quando acompanhada pela diminuição da incidência e mortalidade. Este fato se deve provavelmente pela melhora nos cuidados de prevenção (SANT et al., 2006). Dentro desse contexto, outro ponto positivo para a melhoria dos índices de sobrevida é a triagem precoce pela mamografia que auxilia tanto no aumento da sobrevida quanto na redução da mortalidade (POPAT e SMITH, 2006).
27 De acordo com o Ministério da Saúde, em 2002 registrou-se uma variação percentual relativa de mais de 80% em pouco mais de duas décadas: a taxa de mortalidade padronizada por idade, por 100.000 mulheres, aumentou de 5,77 em 1979, para 9,74 em 2000. Além disso, o número de óbitos em 2005 foi maior para câncer de mama com 9.082, seguido do câncer da próstata com 8.389 e leucemia com 4.816 (MINISTÉRIO DA SAÚDE, 2005).
De acordo com a estimativa do INCA de novos casos para 2006, o CM é o tipo mais incidente entre as mulheres em todas as regiões do país, tendo o Sudeste as mais altas taxas em número de casos (28.640), seguido do Sul (9.540), Nordeste (7.120) e Norte (1.110). Em Belo Horizonte, a estimativa é de 850 novos casos para 2006 (INCA, 2006).
28
2.2.2
DETECÇÃO, DIAGNÓSTICO, ESTADIAMENTO
O Instituto Nacional do Câncer (INCA, 2006) preconiza as ações de detecção e diagnóstico. A detecção precoce é feita pelo exame clínico anual das mamas, rastreamento por mamografia para as mulheres a partir dos 40 anos, e a partir dos 35 para as que possuem um ou mais fatores de risco e, por fim, pelo auto-exame das mamas mensalmente.
São considerados como grupos populacionais com risco elevado para o desenvolvimento do CM:
As mulheres com história familiar de pelo menos um parente de primeiro grau (mãe, irmã ou filha) com diagnóstico de CM, abaixo dos 50 anos de idade;
Com história familiar de pelo menos um parente de primeiro grau (mãe, irmã ou filha) com diagnóstico de CM bilateral ou câncer de ovário, em qualquer faixa etária;
Mulheres com história familiar de CM masculino; ou ainda, mulheres com diagnóstico histopatológico de lesão mamária proliferativa com atipia ou neoplasia lobular in situ. O diagnóstico do CM é feito como parte do exame clínico e ginecológico, e, quando detectada uma lesão palpável, para as mulheres com idade menor que 35 anos é recomendável a avaliação através ultra-sonografia das mamas, e para as mulheres acima de 35 anos é recomendada a mamografia (MMG).
Diante de qualquer tipo de suspeita, incluindo as lesões não palpáveis, a mulher deve ser encaminhada para a punção aspirativa da mama ou punção aspirativa com agulha grossa (core biopsy), permitindo um diagnóstico citológico mais específico. Portanto, ao obter diagnóstico de doença benigna da mama (DBM) ou de CM, a mulher é então encaminhada para a cirurgia conservadora ou não conforme a gravidade, o estadiamento, o tamanho do nódulo/tumor, etc.; seguido da radioterapia e quimioterapia conforme os riscos de recidiva ou outros fatores de risco para metástase.
29 Quanto à localização mais freqüente por distribuição em quadrantes, dados clínicos de registros tumorais revelam que os tumores de mama ocorrem em 66% das pacientes no lado externo, 22% no lado interno e 12% no centro da mama. É instigante o fato de que a maior porcentagem de acometimento de área pelos tumores é no quadrante superior externo (48 a 54%). Um dos fatores que avaliam a razão por esse acontecimento é explicado pelo volume de tecido, fato este debatido entre os diversos estudos (ELLSWORTH et al., 2004).
2.2.3
FATORES DE RISCO
2.2.3.1 FATORES SÓCIO-DEMOGRÁFICOS
A idade é o principal fator de risco para o CM. A incidência do CM é mais baixa (menos que 10 novos casos por 10.000 mulheres) antes dos 25 anos de idade e aumenta cerca de 100 vezes depois dos 45 anos (DUMITRESCU e COTARLA, 2005).
Outros fatores que podem contribuir com o aumento do número de CM, são as condições sócio-econômicas, sendo que o aumento do nível educacional das mulheres e do número de emprego fora de suas casas, as leva a se casarem e constituírem família mais tardiamente, e, a idade mais tardia da primeira gestação, é um fator de risco bem determinado para CM na literatura (GAIL et al., 1989).
2.2.3.2 FATORES GINECO-OBSTÉTRICOS E HORMÔNIOS EXÓGENOS
Dentre os fatores gineco-obstétricos, o risco de desenvolvimento do CM está diretamente relacionado ao tempo de exposição a hormônios sexuais, por seu potencial ativador da mitose, o qual é determinado ao longo da vida da mulher por variáveis como idade da menarca, da primeira gestação completa, número de gestações, idade da menopausa e lactação prolongada.
30 No entanto, fatores que induzem a diferenciação desse tecido, como acontece na gestação e na lactação, oferecem uma proteção (EUROPEAN SOCIETY OF HUMAN REPRODUCTION AND EMBRYOLOGY, 2004)
A idade da menarca menor que 12 anos versus maior que 14 anos têm sido associados com o aumento do risco de CM entre 10 e 20% de magnitude, provavelmente pelo fato da exposição prolongada do epitélio da mama ao estrogênio e progesterona na regulação do ciclo ovulatório (HANKINSON et al., 2004).
Quanto à idade da menopausa, quanto mais tardia, maior é o risco de CM pelo mesmo motivo da idade da menarca e aumenta cerca de 3% a cada ano de não suspensão da menstruação, por outro lado, a retirada dos ovários ou do útero, antes dos 35 anos resultam na diminuição do risco (COLLABORATIVE GROUP ON HORMONAL FACTORS IN BREAST CANCER, 1996; BERNSTEIN, 2002).
A idade da primeira gestação completa antes dos 20 anos versus maior que 30 anos e a alta paridade têm sido associados com menor risco de CM (MCPHERSON K. et al., 2000).
A lactação prolongada tem sido demonstrada como fator protetor. Há uma diminuição de 4,3% no risco relativo de desenvolver CM a cada 12 meses de amamentação, diminuindo 7% a cada gestação. Essa proteção pode ser justificada pela ação da prolactina, hormônio que age diretamente na glândula mamária durante a gestação promovendo o seu crescimento, e durante a lactação, para possibilitar a amamentação (COLLABORATIVE GROUP ON HORMONAL FACTORS IN BREAST CANCER, 1996).
O uso de hormônios sexuais como contraceptivos iniciou em 1960 e, desde então, milhões de mulheres em todo o mundo os utilizam. Os contraceptivos orais (CTO) e a terapia de reposição hormonal (TRH) perfazem a maioria dos hormônios exógenos utilizados por mulheres. Os hormônios exógenos afetam o crescimento da glândula mamária, e, por isso são investigados quanto à sua relação no desenvolvimento no CM (EUROPEAN SOCIETY OF HUMAN REPRODUCTION AND EMBRYOLOGY, 2004).
31 Além disso, o uso de CTO precoce, principalmente antes dos 20 resultou no aumento do risco se comparado com o uso mais tardio (COLLABORATIVE GROUP ON HORMONAL FACTORS IN BREAST CANCER, 1996). Durante a fase reprodutiva, os ovários fabricam mensalmente estrógeno e progesterona, produção que vai diminuindo à medida que a mulher se aproxima da menopausa. A TRH é o tratamento dado às mulheres que se encontram nessa fase e apresentam alguns sintomas físicos como onda de calor, dificuldade para controlar a temperatura do corpo, pele mais flácida, mucosas dos sistemas urinário e reprodutivo mais friáveis e secas, alterações de humor e até quadros depressivos. Entretanto, há alguns anos, grandes estudos têm demonstrado resultados controversos a essa prática. A TRH depois da menopausa aumenta o risco de CM dependendo da duração à exposição desses hormônios e do tipo de combinação com progestinas, no entanto, o uso de anti-estrógenos como o tamoxifeno reduz a incidência do risco (COLLABORATIVE GROUP ON HORMONAL FACTORS IN BREAST CANCER, 1997).
Outros autores investigaram o risco da recidiva de CM avaliado com o uso de TRH, evidenciando um aumento dos casos de recidiva, demonstrando a necessidade de alternativas seguras para tratamento dos sintomas da menopausa em mulheres com CM precoce (CHLEBOWSKI e COL, 2004).
Além disso, aumento do risco de CM com a TRH, é biologicamente plausível e é consistente com dados prévios de grandes estudos como o WISDON “Women´s International Study of Long Durantion Oestrogen after Menopause” (MACLLENNAN e STURDEE, 2002) e o Million Women Study (BERAL, 2003).
2.2.3.3 COMPOSIÇÃO CORPORAL E FATORES ANTROPOMÉTRICOS
32 O ganho de peso nessas condições é associado ao aumento do risco de doenças crônicas não transmissíveis, incluindo vários tipos de câncer (THOMPSON et al., 2004). Existem evidências de associação direta entre índices de massa corporal (IMC) e risco de câncer de mama (ZURLO et al., 1990) tendo sido estimado um risco atribuído de mais de 30% dos cânceres de mama nas sociedades industrializadas por causa da existência de outros fatores de risco juntamente com a provável falha na prevenção do ganho de peso (THOMPSON et al., 2004). Por outro lado, quando a massa livre de gordura foi avaliada, demonstrou-se uma relação inversa quanto ao risco de desenvolvimento de CM (MELLEMKJAER et al., 2006).
Essa relação da composição corporal e a obesidade têm uma complexa relação com o risco do CM e parece ser modulado pela menopausa. Um possível mecanismo para o aumento desse risco é o alto nível de estrógenos em mulheres obesas, sendo o tecido adiposo uma importante fonte de estrógenos (MCTIERNAN et al., 2003). Alguns estudos ainda relatam a diminuição do risco em mulheres obesas na pré-menopausa, mas o mecanismo não está claro (HUANG et al., 1997; M., 2001; LAHMANN et al., 2004).
Outros parâmetros antropométricos como a altura, massa adiposa e circunferência da cintura também são relatados na literatura como fatores de risco para CM (MANJER et al., 2001).
Estudos coortes prospectivos foram analisados em conjunto para avaliar as associações entre os índices antropométricos e o risco de CM. Juntos, foram analisadas 337.819 mulheres sem CM e 4.385 com CM. A análise multivariada ajustada por fatores reprodutivos, dieta, entre outros, demonstrou que a altura é um fator de risco independente para CM na menopausa, que o alto IMC obteve uma relação inversa na pré-menopausa e positiva na menopausa. Portanto, o controle do peso pode reduzir o risco de CM em mulheres na pós-menopausa (VAN DEN BRANDT et al., 2000).
33 2.2.3.4 FATORES DIETÉTICOS
A associação entre consumo alimentar e CM é altamente debatida. A metodologia utilizada nos estudos de consumo é diversificada, e a aproximação qualitativa e quantitativa da ingestão fica cada vez mais dificultada pela falta de padronização e dificuldade de detalhamento inerente do consumo, o que é responsável pela variedade de resultados encontrados. Mecanismos que regulam o balanço energético envolvem interações complexas entre genética, ambiente e comportamento (CECIL et al., 2006).
Modificações na sociedade e avanços tecnológicos têm levado a um aumento da ingestão de gordura na dieta e menos atividade física, contribuindo para a epidemia da obesidade (HILL e PETERS, 1998), que, além de outras conseqüências, aumenta o risco de câncer (ZURLO et al., 1990).
Em uma avaliação da relação de macronutrientes, padrões e grupos alimentares com o risco de CM, não foi observado aumento do risco e ingestão de calorias, macronutrientes ou tipos de lipídios. Interessantemente, houve aumento do risco quanto ao percentual de calorias originadas de carboidratos simples e à freqüência de ingestão dos mesmos. Esse resultado se deveu, provavelmente, segundo os autores, ao aumento da exposição à insulina favorecido por este padrão alimentar (POTISCHMAN et al., 2002). No entanto, esultados de avaliação da freqüência alimentar de 88.678 mulheres do Nurses’Health Study, não demonstraram associação substancial entre quantidade de fibra da dieta, tipos de fibra, carboidratos, índice glicêmico dos alimentos e o risco de CM (HOLMES et al., 2004).
34 Por outro lado, o estudo prospectivo “The Singapure Chinese Health Study”, com 35.298 mulheres de 45 a 74 anos, o consumo de lipídios saturados, monoinsaturados ou LC-PUFA não foi relacionado ao risco de câncer de mama e o The Nurse’s Health Study não encontrou aumento do risco com alta ingestão de gordura na dieta, assim como outros autores (BYRNE et al., 2002).
A importância do consumo de lipídios no risco de câncer parece estar mais relacionada ao perfil de ácidos graxos componentes da dieta e, recentemente os trabalhos têm mostrado a relação entre o consumo de ácidos graxos poliinsaturados de cadeia longa (LC-PUFA) como os ácidos graxos n-3 (LC-PUFA n-3) e ácidos graxos n-6 (LC-PUFA n-6) com o risco de vários tipos de câncer (WAHLE et al., 2003; SAMPATH e NTAMBI, 2005).
O LC-PUFA n-3 e o LC-PUFA n-6 são encontrados em alimentos como óleos de peixe e algumas sementes vegetais oleaginosas como a linhaça, e nos óleos produzidos a partir dessas sementes como soja, girassol, canola, entre outros, respectivamente (MAHAN e ESCOTT-STUMP, 1995). Os derivados dos LC-PUFA da dieta são componentes estruturais vitais de todas as membranas celulares na forma de fosfolipídios, e a sua presença em diferentes proporções altera a fluidez da membrana, uma característica física que pode modular a atividade de uma série de proteínas (enzimas, receptores, canais iônicos) que estão incorporadas na matriz lipídica das membranas celulares. Além disto, os derivados destes ácidos graxos são elementos biologicamente ativos envolvidos em diversas funções celulares como sinalização molecular e metabolismo de modulação refinada, tais como os de esteróides e prostraglandinas (SAMPATH e NTAMBI, 2005).
35 Os mecanismos quimiopreventivos potenciais do LC-PUFA n-3 que podem explicar seus efeitos sobre a carcinogênese foram revisados (LARSSON et al., 2004). Os LC-PUFA n-6 e LC-PUFA n-3 são relacionados à via enzimática que leva a formação de eicosanóides. Há uma gama de eicosanóides presentes nessa via que possuem variadas funções na modulação da resposta inflamatória, agregação plaquetária, crescimento e diferenciação celular.
A interação do metabolismo de LC-PUFA n-6 e LC-PUFA n-3 começa na atuação das enzimas 5-desaturase, seguido pela elongase e posteriormente pela 6-desaturase. Assim, o metabolismo de LC-PUFA n-6 resulta em ácido araquidônico (AA) e de LC-PUFA n-3 resulta no ácido eicosapentaenóico (EPA). Através das fosfolipases de membrana (PLA2), o AA pode se converter em EPA. O EPA, por sua vez, pode se converter no ácido docosahexaenóico (DHA). O AA, pela via das cicloxigenases (COX-1 e 2, enzimas que produzem prostraglandinas e tromboxanos) e lipoxigenases (LOX-5, 12 e 15, enzimas que produzem leucotrienos, hidroxiácidos graxos e lipoxinas), resultam em derivados que ativam o processo de inflamação, angiogênese, inibição da apoptose e adesão celular, levando ao progresso do tumor. No entanto, os produtos gerados pela COX e LOX derivados do EPA, acarretam efeitos antiinflamatórios e pró-apoptóticos, sendo contrários aos efeitos dos derivados do AA (LARSSON et al., 2004).
A proporção dos LC-PUFA nas membranas celulares, bem como o tipo celular, são fatores primários na produção dos variados eicosanóides. Um dos mecanismos de como os LC-PUFA n-3 podem diminuir o risco para o câncer e talvez o mais evidente, é através da supressão da biossíntese de eicosanóides derivados do AA (LARSSON et al., 2004).
Essa supressão se dá de várias formas: primeiro, a alta ingestão de LC-PUFA n-3 acarreta a sua incorporação nas membranas, o que leva a uma maior disponibilidade para as PL2; segundo, por competição pelas enzimas iniciais da via, pelas quais têm uma maior afinidade; terceiro, LC-PUFA n-3 inibe a COX-2 e compete com LC-PUFA n-6, acarretando um aumento na síntese de EPA e seus derivados. Estes derivados são substratos preferenciais das LOX atuantes da via. Portanto, em diferentes níveis, LC-PUFA n-3 reduzem a formação de eicosanóides derivados do AA que estão ligados à modulação da inflamação, proliferação celular, apoptose, metástase e angiogênese (LARSSON et al., 2004).
36 Os LC-PUFA n-3 e seus metabólitos podem exercer alguns efeitos anti-tumorais por afetar a atividade de fatores de transcrição, expressão gênica e transdução de sinais, que geram modificações no metabolismo, crescimento celular e diferenciação (LARSSON et al., 2004).
O primeiro fator de transcrição a ser identificado como regulado por ácidos graxos foi o receptor ativado por proliferação de peroxissomos (PPAR). Os ligantes naturais do PPARG são LC-PUFA n-6 e n-3, AA e EPA, como também outros eicosanóides derivados. O PPARG é expresso em vários tecidos importantes em cânceres humanos (ROSEN e SPIEGELMAN, 2001).
Existem outros genes que têm sua expressão regulada por LC-PUFA n-3 como o fator de transcrição nuclear k (NF-k), Ras e proteína C quinase (PKC), ornitina descarboxilase (ODC), 3-hidroxi-3metilglutaril coenzima-A redutase (HMG-CoA), COX2 e LOXs, que ampliam a atuação dos LC-PUFA n-3 na supressão do câncer (VAMECQ e LATRUFFE, 1999). A dieta com LC-PUFA n-3 no entanto, não mostrou afetar mutações no gene Ha-ras em glândulas mamárias em ratos (RONAI et al., 1991). O potencial protetor da ingestão de vitaminas nos diversos tipos de câncer tem sido bem estabelecido, como o envolvimento da vitamina C com um menor risco de câncer nasofaríngeo (FARROW et al., 1998) e a ingestão de vitamina A com displasia cervical (NAGATA et al., 1999).
Evidências crescentes sugerem que o baixo consumo de folatos pode ser um fator importante na carcinogênese mamária (KIM, 1999). O folato tem sido exaustivamente investigado neste sentido devido a sua função como doador de grupos metila para a síntese de nucleotídeos e metilação biológica. Culturas celulares de animais e estudos humanos mostraram que a deficiência de folato induz a destruição do DNA, assim como alterações no
37 Como relatado, a deficiência de ácido fólico pode levar a danos cromossomais, e, com isso, ao aumento do risco para vários cânceres. Outro ponto é o pontencial de acarretar o acúmulo da homocisteína, o qual tem sido associado a doenças cardiovasculares e defeitos no tubo neural em fetos (AMES, 1999). A vitamina B12 e B6 também agem como o ácido fólico participando da mesma via de metilação do DNA sendo que na deficiência dessas vitaminas ocorrem quebras cromossomais e hipometilação do DNA podendo levar ao aumento do risco de cânceres (AMES, 1999).
2.2.3.5 INGESTÃO DE ÁLCOOL
Estima-se que 2% dos casos de CM nos Estados Unidos (TSENG et al., 1999a) e 15% na Itália (MEZZETTI et al., 1998) são atribuídos ao consumo de álcool. Estudos epidemiológicos têm mostrado consistentemente que o consumo de álcool está relacionado ao aumento do risco de CM (TJØNNELAND et al., 2003; DUMITRESCU e SHIELDS, 2005; BOFFETTA e HASHIBE, 2006). Uma dose de álcool por dia está associada de 1,1 até 4 vezes mais chances de desenvolver o CM (SINGLETARY e GAPSTUR, 2001). Isso pode ser responsável em parte por alterar os níveis de citocinas em vários tecidos como mostrado in vitro (SAEED et al., 2004; CREWS et al., 2006). O risco é dose dependente como demonstrado por um estudo experimental que observou que, para cada 60mg de álcool/dia, existe um aumento de 9% no risco (SMITH-WARNER et al., 1998).
38 2.2.3.6 ATIVIDADE FÍSICA
A Organização Mundial de Saúde, tem estimado que a inatividade física é responsável por 1,9 milhões de mortes por ano (WHO, 2002). Sendo uma das causas da obesidade, a inatividade física contribui fortemente para a morbidade e mortalidade populacional (CAMPBELL, 2003). A associação entre atividade física e o risco de CM é ainda muito controverso. Um grande problema é que não existe um método simples para avaliar a atividade física e é ainda mais difícil quantificar ou predizer em qual época da vida a atividade física é benéfica (BIGLIA et al., 2004). Contudo, muitos estudos encontram uma proteção do exercício físico sobre o risco do CM (DIRX et al., 2001; FRIEDENREICH, 2001; LEE et al., 2001).
Recente meta-análise de 4 estudos coorte e 19 estudos caso - controle investigou a relação do risco de CM com atividade física e demonstrou uma redução de 20% entre adolescentes e adultos jovens de 12 a 24 anos.
Para cada uma hora de atividade física por semana, o risco diminui 3%, (DE JONG et al., 2002). A atividade física pode reduzir o risco por modificar os níveis de hormônios biodisponíveis (HANKINSON et al., 2004). A atividade física também pode estabelecer um efeito protetor no CM pelo aumento da apoptose induzida pelo estresse oxidativo oriundo da peroxidação lipídica que ocorre durante o exercício (GAGO-DOMINGUEZ, 2006).
2.2.3.7 HISTÓRIA PESSOAL DE DOENÇA BENIGNA DA MAMA, HISTÓRIA FAMILIAR DE CÂNCER DE MAMA E DENSIDADE MAMOGRÁFICA
39 Estudo de grande consistência como o realizado pelo Grupo Colaborativo de Fatores Hormonais no CM, demonstrou, através de uma comparação de 53 estudos epidemiológicos, que 12% das mulheres com CM tem um membro na família e 1% tem mais de um membro com CM (COLLABORATIVE GROUP ON HORMONAL FACTORS IN BREAST CANCER, 2001; SINGLETARY e GAPSTUR, 2001; EASTON, 2002).
A densidade mamográfica é outro fator de risco bem estabelecido tanto em mulheres na pré quanto na pós-menopausa. De acordo com dois grandes estudos, o Breast Cancer Detection Demonstration Projetct (BYRNE et al., 1995) e o Canadian National Breast Screning Study (BOYD et al., 1995), mulheres com 75% da mama considerada densa na mamografia (MMG) têm aproximadamente 5 vezes mais chance de risco de desenvolvimento no CM em comparação àquelas que têm menos de 5%. Este fator está relacionado ao número de células na mama, aumentando o risco naquelas com maior densidade (BIGLIA et al., 2004).
Um estudo comparativo entre vários estudos prospectivos relacionando a densidade das mamas com o CM reforça esse fator de risco assim como esclarece que não só a densidade das mamas está relacionada ao risco, mas também a heterogeneidade do padrão tecidual no parênquima mamário (TORRES-MEJIA et al., 2005). A TRH influencia na densidade das mamas, o que pode ser um complicador. O seu uso na menopausa pode dobrar suas chances de ter maior densidade mamográfica quando comparada a mulheres que nunca fizeram TRH (SALA et al., 2000).
2.2.3.8 OUTROS FATORES
A exposição da glândula mamária à radiação ionizante é um fator de risco bem reconhecido para o CM. São importantes preditores dessa avaliação, a idade e o tempo de exposição (PRESTON et al., 2002). Estudos com populações sobreviventes das bombas atômicas de Hiroshima e Nagasaki têm demonstrado uma relação inversa entre o risco e a idade de exposição à radiação. Além disso, o risco é dose-dependente e tende a diminuir progressivamente com o tempo (LAND et al., 2003).
40 Os indivíduos fumantes têm produção de IL-8 em células sanguíneas estimuladas indicando que o hábito de fumar pode ter efeitos sistêmicos na resposta inflamatória (VAN DER VAART et al., 2005). Estudos têm demonstrado que os indivíduos que começam a fumar mais precocemente (antes dos 16 ou 17 anos) aumentam o risco para CM independente do tempo de exposição (BARON et al., 1996; MARCUS et al., 2000; EGAN et al., 2002). Um extenso estudo do tipo caso-controle, com 17.000 casos de CM concluiu que há um aumento pequeno no risco para mulheres na pós-menopausa que começaram a fumar antes dos 16 anos de idade (BARON et al., 1996).
Entre os agentes quimiopreventivos, o tamoxifeno é o único que a Food and Drug Administration (FDA) aprova na terapia de mulheres com alto risco de CM. O raloxifeno, outro modulador do receptor de estrógeno (SERM) está sendo estudado e comparado ao tamoxifeno na clínica pelo Study of Tamoxifen and Raloxifene. O Anastrozole, que é um inibidor de uma enzima chave envolvida na conversão de andrógenos para estrógenos em tecidos periféricos é também avaliado como possível agente quimioprotetor pelo
Internacional Breast Intervention Study II (DUMITRESCU e COTARLA, 2005).
O uso de aspirina ou outras drogas antiinflamatórias não esteroidais (AINE) como os inibidores da enzima COX-2, que são enzimas relacionadas à síntese de prostraglandinas e tromboxanos derivados do ácido araquidônico (AA), têm sido inversamente relacionado a incidência de CM (COTTERCHIO et al., 2001; ARUN e GOSS, 2004),indicando que a alta resposta inflamatória pode elevar esse risco (GONZALEZ-PEREZ et al., 2003; RAHME et al., 2005; HARRIS et al., 2006).
Tanto em modelos animais quanto in vitro, os estudos indicam que os inibidores da COX-2 e outros AINEs têm efeitos quimiopreventivos no CM (MAZHAR et al., 2006), mas outras evidências epidemiológicas são ainda conflitantes (GONZALEZ-PEREZ et al., 2003; MAZHAR et al., 2006).
41
2.2.4
EPIDEMIOLIA MOLECULAR DO CM
O gene P53 foi o primeiro supressor de tumor relacionado ao CM, sendo o gene mais comumente encontrado mutado em todos os cânceres humanos (MALKIN, 1994). Os genes
BRCA1 e BRCA2 (Breast Cancer 1 e 2), quando mutados, são os mais implicados nos casos de CM hereditário e são responsáveis por 80 a 90% de todos os cânceres hereditários na mama, não sendo freqüentemente encontrados mutados nos casos de câncer esporádico (AMENDOLA e VIEIRA, 2005).
O BRCA1 é um gene supressor de tumor cuja função primária é a manutenção da integridade genômica, chamado por essa razão de caretaker gene. Encontra-se mutado em 42% dos cânceres de mama familiar. O BRCA2 é outro caretaker gene envolvido no sistema de reparo celular é encontrado por 76% dos cânceres de mama familiar (AMENDOLA e VIEIRA, 2005).
As prevalências estimadas para portadores de mutações em BRAC1/2 são respectivamente, 0,11% e 0,12% na população geral e entre 12,8% e 16% em família de alto risco com três ou mais casos de CM ou ovário. (AMENDOLA e VIEIRA, 2005). Mutações no BRCA1 e 2 são responsáveis por aproximadamente 20% do total de risco familiar observado (VERONESI, 2002). Os restantes 80% se devem a uma combinação dos efeitos produzidos por mutações em outros genes (cerca de 1%) e alterações em genes ainda não descritos (AMENDOLA e VIEIRA, 2005).
42 O rápido progresso na codificação e anotação do genoma humano tem acelerado as pesquisas na função dos polimorfismos genéticos na patogênese das doenças multifatoriais (LOKTIONOV, 2003). Polimorfismos associados aos cânceres envolvem genes que codificam enzimas do metabolismo de xenobióticos, enzimas de reparo do DNA, fatores envolvidos no controle do ciclo celular e proteínas relacionadas à regulação hormonal. Polimorfismos de baixa penetrância relacionados a susceptibilidade ao CM podem ser sugeridos por mecanismos fisiológicos e bioquímicos envolvidos na carcinogênese da mama. Candidatos a genes polimórficos incluem aqueles que codificam enzimas implicadas no metabolismo de estrogênio ou de vários carcinógenos, de detoxificação ou espécies oxi-reativas que emergem dessas reações, de enzimas relacionadas a metilação do DNA, ao metabolismo do folatos e de fatores relacionados a angiogênese.
O gene CYP1A1, codifica uma hidroxilase, que ativa hidrocarbonos aromáticos constituintes do cigarro, levando a formação de moléculas carcinogênicas (BARTSCH et al., 2000). Em conseqüência, essa enzima também catalisa a hidroxilação do estradiol em vários tecidos extra-hepáticos, incluindo as mamas (HELLMOLD et al., 1998). Entre os polimorfismos identificados nesse gene, o polimorfismo m1 (MspI) é associado com um modesto aumento do risco CM em mulheres brancas e o m2 é associado a um aumento moderado do risco somente em mulheres na pós-menopausa (DE JONG et al., 2002).
Por outro lado, o gene CYP2D6 é membro da família do citocromo P450 que codifica uma hidroxilase que também tem sido associado ao aumento do risco de susceptibilidade ao CM (DE JONG et al., 2002).
Polimorfismos nos genes que estão envolvidos no metabolismo dos hormônios esteróides podem alterar os níveis e ou ações de hormônios endógenos e então influenciar no risco de CM. Outro gene da família do citocromo p450 é o CYP19, implicado na biossíntese de estrogênio, que apresenta um polimorfismo de microssatélite, que em alguns estudos tem apresentado relação com o risco de CM (DE JONG et al., 2002). Por outro lado, alguns estudos demonstraram que homozigotos do PROGINS, uma subfamília de inserção de Alu de 306pb, do gene do receptor de progesterona, diminuiu o risco do CM (DE JONG et al., 2002).
43 O gene GSTP1 é expresso tanto em tecidos normais quanto no CM. Uma meta-análise que estudou o polimorfismo desse gene, que se deve a uma substituição de uma isoleucina por uma valina no códon 105, leva a redução da atividade da enzima e tem sido associado a um aumento do risco de CM em homozigotos (DE JONG et al., 2002).
Como a ingestão de álcool é um fator de risco investigado, a análise de polimorfismos envolvidos na oxidação do álcool tem sido realizada e indicam que a dehidrogenase alcoólica (ADH) pode modular o risco de CM. Mulheres na pré-menopausa com o genótipo ADH1C*1,1 têm 1,8 vez mais risco de CM que mulheres com os outros 2 genótipos (ERGUL et al., 2003).
Outro grupo de genes que estão envolvidos na metilação do DNA ilustra uma importante interação ambiental com os fatores genéticos. Os polimorfismos C677T e A1298C do gene da 5,10 metilenotetrahidrofolato redutase (MTHFR), relacionado à manutenção dos padrões de metilação do DNA, são associadas ao aumento do risco de desenvolvimento do CM de maneira dependente da ingestão de folato (CAMPBELL et al., 2002; ERGUL et al., 2003).
Os genes de reparo constituem outro grupo de genes de susceptibilidade, de baixa penetrância. Polimorfismos nesses genes diminuem a capacidade do sistema de reparo do DNA, especialmente depois de expostos a agentes genotóxicos exógenos e endógenos, contribuindo para o risco de desenvolvimento do CM (CAMPBELL et al., 2002; ERGUL et al., 2003). Um gene envolvido no reparo por excisão de base, o XRCC1399Q parece conferir um aumento no risco de desenvolvimento do CM em afro-americanas (DUELL et al., 2001; CAMPBELL et al., 2002).
Outros polimorfismos em genes como o do receptor de estrógeno (ER), heat shock protein 70 (HSP70) ou no fator de necrose tumoral (TNF) podem também influenciar o risco do desenvolvimento do CM (DE JONG et al., 2002).
44
2.3 RECEPTORES
ATIVADOS
POR
PROLIFERAÇÃO
DE
PEROXISSOMOS – PPAR
O envolvimento dos fatores de transcrição PPAR no desenvolvimento de doenças crônicas multifatoriais como diversos tipos de câncer, diabetes e aterosclerose tem sido recentemente investigado por diversos autores (KERSTEN et al., 2000).
PPARS são fatores de transcrição induzidos por ligantes, que pertencem a uma superfamília de 48 membros de receptores nucleares hormonais (GODOY-MATOS, 2005) junto com os receptores hormonais da tireóide, de hormônios esteróides, retinóides e vitamina D (BOITIER et al., 2003). De acordo com recente nomenclatura proposta para os receptores nucleares, os PPARs formam o Grupo C na subfamília 1 da superfamília de receptores hormonais (BOITIER et al., 2003).
O primeiro PPAR foi descoberto em 1990 e foi fundamental para o entendimento do processo de proliferação do peroxissoma celular (ISSEMANN e GREEN, 1990). Esse receptor funciona como um sensor nuclear de vários processos proliferativos que ocorrem em resposta a estímulos nutricionais e farmacológicos. Inicialmente porém, a relevância médica para o ser humano permaneceu questionável até que estudos mais recentes evidenciaram o papel dessa família de receptores na diferenciação dos adipócitos, no metabolismo glicídico e lipídico, na regulação da função vascular, (ISSEMANN e GREEN, 1990) e, mais recentemente, vários estudos discutem a sua provável influência no diabetes e câncer (GODOY-MATOS, 2005).
Os receptores PPAR são ativados através da heterodimerização com o receptor do ácido retinóico (RXR). Assim, eles funcionam como sensores aos quais se ligam uma série de moléculas relacionadas ao metabolismo como ácidos graxos, eicosanóides e oxiesteróis (GODOY-MATOS, 2005). A ativação dos PPARs induz a transcrição de vários genes, todos virtualmente associados ao metabolismo. No processo de transcrição, os heterodímeros ativados pelos ligantes recrutam co-ativadores.
45 Estruturalmente, os PPARs possuem cinco ou seis regiões estruturais em quatro domínios funcionais (A - F), como esquematizado na Figura 1. A porção amino-terminal A/B possui a função de ativação transcricional independente de ligante (AF-1). A região C composta por duas regiões dedos de zinco, contém o domínio de ligação ao DNA (DBD) no qual o alvo do receptor são as seqüências específicas no DNA. A região D é um importante sítio para ligação de co-fatores. Os domínios E/F servem para a interação com ligantes (LBD), uma interface dimérica que depende da ativação do domínio AF-2. Os ligantes do PPARG modulam a comunicação intramolecular entre a porção amino-terminal A/B e a carboxi-terminal LBD (WANG et al., 2006). A ligação ao DNA requer a formação do heterodímero PPAR e RXR e que interagem com os genes alvo.
46 O heterodímero PPAR-RXR liga-se, na maioria das vezes, aos elementos responsivos do PPAR (PPRE), que nada mais são do que seqüências hexaméricas específicas (TGACCTNTGACCT) que ocorrem sequencialmente espaçadas por um nucleotídeo presentes nos genes alvo (KLIEWER et al., 1992), constituindo um motivo de ligação da proteína com o DNA. Outros autores relatam uma seqüência consenso diferente (AGGTCA), sendo esta, espaçado por um ou dois nucleotídeos (MEIRHAEGHEA e AMOUYEL, 2004). Existem três tipos de PPAR: alfa (PPARA), gama (PPARG) e beta (PPARB) que têm sido identificados por exibirem distribuição distinta e serem associados a ligantes seletivos (WANG et al., 2006).
Os PPARs reprimem a transcrição por transativação de alguns genes que não possuem o PPRE. PPARA pode reprimir a expressão do gene NF-KB por seqüestrar a sua subunidade p65. O PPARB pode reprimir o gene BCL6 por sua própria ligação como repressor, que, quando ativado, libera a proteína, que liga-se a um outro fator de transcrição, inibindo sua transcrição. E, por último, PPARG pode efetivar a transrepressão por interação a co-repressores chamados SMRT inibindo sua ativação como no caso do PPARB, que não consegue se ligar enquanto o co-repressor estiver presente, e, quando é ativado, libera o SMRT, que liga-se a outro repressor, no caso, o STAT3, que por sua vez, inibe a expressão do gene STAT (TAN et al., 2005).
O PPARA é predominantemente expresso em hepatócitos, cariomiócitos, túbulos proximais nos rins e enterócitos enquanto que PPARB está presente em vários tecidos sendo o mais freqüentemente expresso (WANG et al., 2006). PPARB, apresenta maior expressão no tecido adiposo, pele e cérebro, tendo uma importante função no controle da proliferação celular, diferenciação e sobrevivência dos queratinócitos ou inibição da atividade dos outros PPARs em genes alvo (MICHALIK et al., 2004). É o tipo menos estudado e com funções intrincadas. Parece ter uma inter-relação com os outros PPARs e neles exercer um papel modulador.
47 Existem quatro isoformas do PPARG conhecidas: PPARG 1, PPARG 2, PPARG 3 e PPARG 4. As isoformas de PPARG em humanos e camundongos se estendem em mais de 100Kb de DNA genômico, que se diferenciam por suas extremidades 5’ e por diferentes eventos de splicing (Figura 2).
Fonte: WANG et al., Critical Reviews in Oncology / Hematology, v.58, p.1-14. 2006 Figura 2 – Organização do gene PPARG humano
48 As proteínas PPARA e PPARG podem também ser regulados por fosforilação produzindo diferentes perfis de afinidade pelos ligantes (BOITIER et al., 2003), como por exemplo, a fosforilação da serina no resíduo 112 que leva a diminuição da sua atividade (LEHRKE e LAZAR, 2005).
No PPARG, a afinidade de ligação também pode ser influenciada por polimorfismos, como é o caso do Pro12Ala, que tem uma substituição da prolina por uma alanina no resíduo 12. Este polimorfismo, específico da isoforma 2, ocorre no exon B e consiste de uma transição de uma citosina para uma guanina (C G). Outros polimorfismos também são encontrados em diferentes isoformas de PPARG, como Pro467Leu, Val290Met, Arg425Cys, Phe388Leu (GODOY-MATOS, 2005) ou ainda o C161T (JIANG et al., 2005). A figura 3 ilustra a localização dos polimorfismos identificados nas isoformas de PPARG.
Fonte: BOITIER et al., Comparative Hepatology, v.2, n.1, p.3. 2003.
49
2.3.1
LIGANTES DO PPAR E SEU PAPEL BIOLÓGICO
A ativação de PPARs é efetivada por ácidos graxos poliinsaturados de cadeia longa n-6 e n-3 (LC-PUFA), ou seus metabólitos derivados principalmente do ácido araquidônico (AA) e linoléico (AL), resultantes das vias das cicloxigenases e lipoxigenase, como eicosanóides, leucotrienos, prostraglandinas e tromboxanos.
Algumas substâncias geradas em estados patológicos, a partir da oxidação de LDL-colesterol (LDL-c), também podem ativar os PPARs e têm importante papel no acúmulo de macrófagos na placa aterosclerótica. O 15-deoxy 12,14-prostaglandina J2 (15PGJ2), derivado do AA é o ligante endógeno mais potente de PPARG e seu principal agonista natural (GROMMES et al., 2004).
Os ligantes farmacológicos dos PPARA são os fibratos e dos PPARG são as tiazolidinedionas (TZD). Os fibratos atuam como hipolipemiantes via ligação com os PPARA e já demonstraram diminuir significativamente o risco cardiovascular. As TZD são ligantes dos PPARG e acarretam efeitos no metabolismo glicídico, lipídico e sistema vascular. Existem compostos não TZD em estudo que possuem afinidade de 30 a 100 vezes maior com os PPARG (VOGEL et al., 2006).
Algumas drogas antiinflamatórias como indometacina e ibuprofeno, ligam-se aos PPARG e PPARA inibindo a enzima COX-2, contribuindo assim para o efeito antiinflamatório. O uso de antiinflamatório não esteroidais (AINEs) tem sido associado à diminuição do risco de CM em estudos epidemiológicos, justamente pelo fato da resposta inflamatória estar associada ao aumento do risco (VOGEL et al., 2006).
50
2.3.2
PPAR, METABOLISMO DE LIPÍDIOS E INFLAMAÇÃO
A ativação dos PPARG ou PPARA ou de ambos acarreta uma diminuição dos triglicerídeos (TGL) por diferentes mecanismos. A concentração de TGL depende da quantidade de substrato disponível (ácidos graxos) e do ritmo do metabolismo considerando a atividade da lípase lipoprotéica (LPL) (GODOY-MATOS, 2005).
A ativação dos PPARA através da ligação dos fibratos ativa a expressão dos genes das apolipoproteínas AI e AII, que são os maiores componentes das partículas de HDL, aumentando o seu nível sérico, que leva por sua vez, a redução da síntese de VLDL, oxidação e captação mitocondrial de ácidos graxos, diminuindo os níveis séricos de TGL.
Os PPARG são fundamentalmente relacionados a adipogênese e a estocagem de ácidos graxos no adipócito. São também importantes no metabolismo dos lipídios extracelulares. As TZD afetam principalmente o metabolismo de TGL por meio do aumento da expressão da LPL. Com o aumento da lipólise mediada pela LPL, aumenta também a quantidade de LDL-c, porém, não está claro se esse efeito acarreta aumento ou diminuição do risco cardiovascular. Até porque existem respostas divergentes entre as diferentes TZD (GODOY-MATOS, 2005). Os PPARG, participam ainda do transporte reverso de colesterol, que é aquele oriundo das células periféricas para o fígado e, posteriormente para excreção, através do efluxo de colesterol pelos M (KERSTEN et al., 2000).
PPARA e o PPARG apresentam propriedades antiinflamatórias e têm contribuído para a supressão da carcinogênese (VAMECQ e LATRUFFE, 1999).
A ativação dos PPARA e PPARG regula negativamente a atividade da COX-2, sem alterar a atividade da COX-1. Esse é um dos mecanismos pelo qual os fibratos podem diminuir a inflamação nas células endoteliais e modificarem a evolução do processo aterosclerótico (GODOY-MATOS, 2005). É observada uma alta expressão de COX-2 em carcinomas mamários inversamente proporcionais ao grau histológico e nuclear (NAKOPOULOU et al., 2005).
51 Esses efeitos são mediados pela ação dos PPARs sobre o fator de transcrição nuclear kappa-beta (NFk), proteína ativadora 1 (AP-1), sinais de transdução e ativadores da transcrição (STAT), todos responsáveis pela regulação da expressão dos genes das citocinas (DELERIVE et al., 2001).
2.3.3
PPARG, ADIPOGÊNESE, OBESIDADE E RESISTÊNCIA À
INSULINA
A ativação de PPARG é o evento regulador chave da diferenciação dos adipócitos. A ativação dos PPARG aumenta a expressão de genes específicos dos adipócitos, como os da proteína ligadora de ácidos graxos (aP2), acil-Coa sintase (ACS), proteína transportadora de ácidos graxos (FATP-1) e LPL (KERSTEN et al., 2000). Por outro lado, a ativação dos PPARG diminui a expressão do gene da leptina (HOLLENBERG et al., 1997) com conseqüente aumento da ingestão alimentar oferecendo maior substrato para a adipogênese. Diminuem também a secreção de TNF-µ bloqueando seu efeito inibitório na cascata de sinalização da insulina e aumentando a liberação dos ácidos graxos livres (PERALDI et al., 1997). Assim, PPARG é considerados um dos principais genes que mantêm a energia corporal (AUWERX, 1999) juntamente com outros hormônios, fatores de crescimento, insulina e vias neuroendócrinas.
Outro papel importante de PPARG é o de remodelação do tecido adiposo, modulando a apoptose de adipócitos grandes e diferenciados, levando a proliferação de adipócitos pequenos e altamente sensíveis à insulina no lugar dos adipócitos diferenciados e menos sensíveis à mesma (GODOY-MATOS, 2005).